No CrossRef data available.
Published online by Cambridge University Press: 02 February 2024
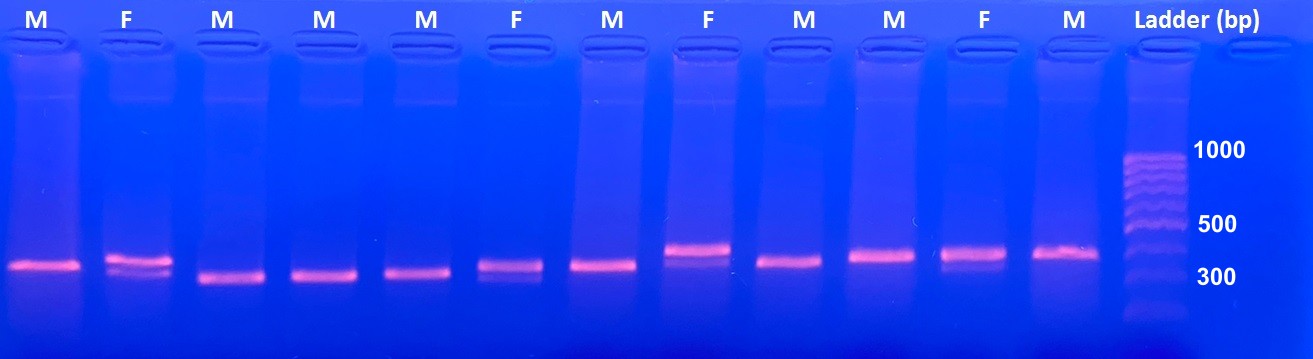
Antioquia Brushfinch Atlapetes blancae is a recently rediscovered Colombian microendemic species with a restricted distribution. Although the species might use a variety of habitats, current information suggests that it is a shrubland specialist. It is categorised as “Critically Endangered” because due to habitat loss its population has been estimated to be smaller than 50 individuals. Such estimates are based on indirect measurements of its population and assumptions regarding its restricted range/habitat. We provide the first direct population estimates and adult sex ratio for seven localities, using mark-resighting techniques. Individuals were captured using mist-nets, bled, and colour banded. Subsequently, we performed censuses along transects to record banded and non-banded individuals. Population estimates were obtained using the classic closed population mark-resighting model. In addition, we used the abundance–area relationship to estimate population size at other sites where the species has been previously recorded. Finally, individuals were sexed by amplifying the Z and W-linked Chromo-Helicase DNA-binding gene (CHD) genes. In total, we banded 55 individuals at four locations. Summarising across the seven sampled localities, we found that the population is most likely composed of 109 individuals in the 122 ha sampled. Based on the abundance–area relationship, we estimated that the population of Antioquia Brushfinch could be as large as 405 individuals in 322 ha of suitable habitat. We found that male/female sex ratio was 0.45 (95% confidence interval [CI] 0.32–0.6) and that it was not significantly biased. Our results suggest that the population of Antioquia Brushfinch is at least eight times larger than suggested by secondary information. We found that populations are properly structured at least regarding sex ratio, although we caution for further research on sex biases of immature and juvenile individuals. We recommend that future studies should focus on population trends and the impact of habitat transformation on the demography of this iconic species.
El Gorrión-Montés Paisa Atlapetes blancae es una especie endémica de Colombia recientemente redescubierta con una distribución restringida. A pesar de que la especie puede utilizar una gran variedad de hábitats, la información disponible sobre la historia natural sugiere que es un especialista de matorrales. Ha sido categorizado como En Peligro Crítico principalmente porque se ha estimado que su población consta de menos de 50 individuos y por la pérdida de hábitat. Tales estimaciones, sin embargo, se basan en mediciones indirectas de su población y suposiciones dado su rango restringido. Aquí proporcionamos las primeras estimaciones utilizando técnicas de marcaje-recaptura. Los individuos fueron capturados usando redes de niebla. Cada individuo fue anillado con anillos de colores utilizando una combinación única por sitio y se tomó una muestra de sangre antes de su liberación. Después de anillar a los individuos, realizamos censos a través de transectos estandarizados que registraron individuos anillados y no anillados para ajustar una distribución hipergeométrica para estimar el tamaño total de la población por sitio. Luego realizamos una estimación bruta de la población presente en sitios con presencia confirmada de esta especie. En total, anillamos a 55 individuos en cuatro localidades. Sumando las siete localidades muestreadas, encontramos que la población probablemente esté compuesta por 109 individuos en 122 ha muestreadas. Con base en una relación individuos-área, estimamos que la población del Gorrión-Montés Paisa podría ser tan grande como 405 individuos en 322 hectáreas de hábitat adecuado. Al agrupar a los individuos en todos los sitios, encontramos que la proporción de sexos macho/hembra era de 0.45 (95% CI 0.32 – 0.6) y no estaba significativamente sesgada. Nuestros resultados sugieren que la población del Gorrión-Montés Paisa es al menos ocho veces mayor que lo sugerido por información secundaria. Además, mostramos que las poblaciones están adecuadamente estructuradas, al menos con base en la proporción de machos y hembras, pero sugerimos que estudios futuros investiguen los sesgos en los sexos en individuos inmaduros y juveniles. Recomendamos que los estudios futuros deben enfocarse en las tendencias poblacionales y el impacto de la degradación del hábitat sobre la demografía de esta especie icónica.