



Introduction
Toyotamaphimeia is an extinct crocodylian lineage, the name of which derives from a Japanese myth, depicting the transformation of a princess into a crocodile. After the original discovery and subsequent taxonomic revision of the type species, Toyotamaphimeia machikanensis (Kamei and Matsumoto in Kobatake et al., Reference Kobatake, Chiji, Ikebe, Ishida, Kamei, Nakaseko and Matsumoto1965), and its holotype (Kobatake et al., Reference Kobatake, Chiji, Ikebe, Ishida, Kamei, Nakaseko and Matsumoto1965; Aoki, Reference Aoki1983), Toyotamaphimeia remained only known to Japan. Interestingly, a recent paper reported the first possible occurrence of Toyotamaphimeia outside of Japan (Ito et al., Reference Ito, Aoki, Hirayama, Yoshida, Kon and Endo2018) based on a historic specimen (WU-HPAC-D 007) originally collected in Tainan, Taiwan (Tokunaga, Reference Tokunaga1936). That specimen (a partial rostrum) was long considered to have been lost during World War II, and its rediscovery in the Waseda University collection (Tokyo, Japan) offers the first evidence of Toyotamaphimeia in Taiwan. As in the Japanese material, the largest maxillary tooth of the Waseda specimen likely occupies the 7th alveolus, but given the limited material, the presence and evolutionary implications of the Toyotamaphimeia lineage in Taiwan remain elusive.
In addition to the Waseda specimen (also from the Pleistocene of Tainan), the second formally published report of crocodylian fossils from Taiwan, corresponding to Tomistoma taiwanicus, which was described as a new species by Shikama (Reference Shikama1972). WU-HPAC-D 007 and the NSMT materials of Tomistoma taiwanicus likely come from the Chiting Formation in Tainan, broadly dated from 0.8–0.4 Ma (Middle Pleistocene). The syntype of Tomistoma taiwanicus includes six specimens: an isolated tooth and five skull/mandible fragments. Owing to the incomplete nature of the type specimens, Shikama (Reference Shikama1972, p. 126) used a question mark in Tomistoma (?) taiwanicus, indicating the uncertain generic attribution of the species. Shikama (Reference Shikama1972) also noticed the morphological similarity between Tomistoma taiwanica and Tomistoma machikanense, later assigned to Toyotamaphimeia by Aoki (Reference Aoki1983).
An obvious pending question then arises: does Tomistoma taiwanicus also belong to Toyotamaphimeia? To clarify this taxonomic issue, we re-examined the type specimens of Tomistoma taiwanicus and conducted phylogenetic analyses to test this hypothesis. Further relevant to this taxonomic question are the underlying paleogeographic interpretations. Taiwan is geographically between the Pleistocene occurrence of Toyotamaphimeia (Japan) and the extant distribution of Tomistoma (Southeast Asia); thus, whether those crocodylian fossils belong to either Tomistoma or Toyotamaphimeia leads to entirely disparate evolutionary scenarios. In addition, if Tomistoma taiwanicus indeed pieces together with the Toyotamaphimeia lineage, this resolves the paleogeographic puzzle of a crocodylian lineage, Toyotamaphimeia, solely inhabiting relatively high latitudes (Japan).This is because Tomistoma taiwanicus, from the Pleistocene of Tainan, is geographically farther south and geologically older, which offers a novel perspective to understand the origin and evolutionary history of Toyotamaphimeia.
Geological setting
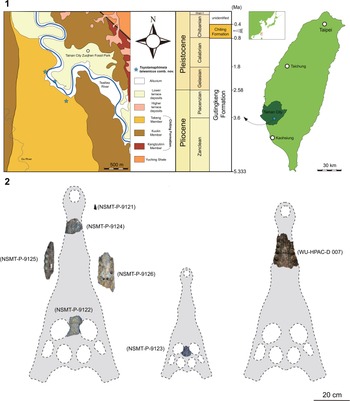
The exact locality of Tomistoma taiwanicus remains uncertain because the late and famous local collector, C. M. Chen, collected the specimen in Tainan without properly documenting specific information and sent it to Japan for study. After the description of Tomistoma taiwanicus, principal paleontologists in Taiwan at that time (e.g., T. Shikama or H. Otsuka) likely acquired more collecting information from C. M. Chen, pinpointing three localities along the Tsailiao River (Fig. 1; Otsuka and Shikama, Reference Otsuka and Shikama1978; Otsuka, Reference Otsuka1984). Based on our recent field experience around the Tainan area, those three identified localities likely do not represent the actual fossil-bearing sites, but pick-up spots that accumulated fossils washed away along the river. A field-digging program teamed up with local government agencies and led by CHT aims to collect more complete materials and reveal the fossil-bearing horizon to further discuss the paleoecological settings. Given the discovery of abundant vertebrate fossils from the Chiting Formation in Tainan since the early twentieth century (Hayasaka, Reference Hayasaka1932; Shikama, Reference Shikama1937; Otsuka and Lin, Reference Otsuka and Lin1984; Chang et al., Reference Chang, Takai and Ogino2012; Tsai et al., Reference Tsai, Fordyce, Chang and Lin2013; Lin et al., Reference Lin, Chien, Lee and Chang2019; Tsai and Mayr, Reference Tsai and Mayr2021; Tsai and Tseng, Reference Tsai and Tseng2022), the age of Tomistoma taiwanicus can be broadly established as 0.8–0.4 Ma (late Early to early Middle Pleistocene; Fig. 1). Of note, Shan et al. (Reference Shan, Wu, Cheng and Sato2009) suggested a Late Pliocene/Early Pleistocene age, but this should be corrected to Middle Pleistocene, as we discussed here.

Figure 1. The occurrence of Toyotamaphimeia taiwanicus n. comb. (1) Location and geological map and generalized stratigraphic column with collecting information (modified from Otsuka, Reference Otsuka1984). (2) Fossil materials of Toyotamaphimeia taiwanicus n. comb. and the possible positions.
Materials and methods
We follow Kobayashi et al. (Reference Kobayashi, Tomida, Kamei and Eguchi2006), Shan et al. (Reference Shan, Wu, Cheng and Sato2009), and Iijima et al. (Reference Iijima, Momohara, Kobayashi, Hayashi, Ikeda, Taruno, Watanabe, Tanimoto and Furui2018) for the anatomical terms. To further explore the phylogenetic position of the Tainan tomistomine, we combined Tomistoma taiwanicus and WU-HPAC-D 007 as a single operational taxonomic unit (OTU) in order to achieve a higher percentage of coding in the cladistic analysis (see Systematic Paleontology section). We adopted the matrix of Iijima and Kobayashi (Reference Iijima and Kobayashi2019), which is mainly based on Brochu (Reference Brochu1999) and Jouve et al. (Reference Jouve, Bouya, Amaghzaz and Meslouh2015). In addition, we expanded the dataset by adding newly discovered fossils, including Hanyusuchus sinensis Iijima et al., Reference Iijima, Qiao, Lin, Peng, Yoneda and Liu2022, Gunggamarandu maunala Ristevski et al., Reference Ristevski, Price, Weisbecker and Salisbury2021, Toyotamaphimeia cf. T. machikanensis (reported by Iijima et al., Reference Iijima, Momohara, Kobayashi, Hayashi, Ikeda, Taruno, Watanabe, Tanimoto and Furui2018), Melitosaurus champsoides Owen, Reference Owen1849, Tomistoma calaritanum Capellini, Reference Capellini1890, Tomistoma gaudense (Hulke, Reference Hulke1871) (reported by Nicholl et al., Reference Nicholl, Rio, Mannion and Delfino2020), and NTUM-VP 200702 (a partial skull from another locality in Taiwan). We also modified Character 145 to a multistate character with (three states) based on Nicholl et al. (Reference Nicholl, Rio, Mannion and Delfino2020) and updated the scorings of Toyotamaphimeia machikanensis and Penghusuchus pani Shan et al., Reference Shan, Wu, Cheng and Sato2009. Our dataset then includes 254 characters and 77 OTUs (see Supplementary material for the updated dataset). We performed the maximum parsimony analysis in TNT v 1.5 (Goloboff et al., Reference Goloboff, Farris and Nixon2008; Goloboff and Catalano, Reference Goloboff and Catalano2016) under Traditional Search. Detailed settings include 1000 Wagner tree replicates; saving ten trees per replicate under tree bisection reconnection branch swapping; equal-weighted and unordered for all characters. Additionally, we performed a second round of skull-only cladistic analysis (193 characters) to test the phylogenetic position of Tomistoma taiwanicus due to the preservation of the type specimens.
Repositories and institutional abbreviations
KSNHM, Natural History Museum, Kishiwada City, Osaka, Japan; MOU, Museum of Osaka University, Osaka, Japan; NMNS, National Museum of Natural Science, Taichung, Taiwan; NSMT, National Museum of Nature and Science (previously known as National Science Museum, Tokyo), Tsukuba, Japan; NTUM-VP, Vertebrate Paleontology (Laboratory of Evolution and Diversity of Fossil Vertebrates), Museum of Zoology, National Taiwan University, Taipei, Taiwan; WU-HPAC, Waseda University-Honjo Paleontological Archaeological Collection, Tokyo, Japan.
Systematic paleontology
Crocodylia Gmelin, Reference Gmelin1789 (sensu Benton and Clark, Reference Benton, Clark and Benton1988)
Tomistominae Kälin, Reference Kälin1955 (sensu Brochu, Reference Brochu2003)
Toyotamaphimeia Aoki, Reference Aoki1983
Type species
Toyotamaphimeia machikanensis (Kamei and Matsumoto in Kobatake et al., Reference Kobatake, Chiji, Ikebe, Ishida, Kamei, Nakaseko and Matsumoto1965) from the Kasuri Tuff of the Osaka Group, Osaka, Japan. The holotype is MOUF00001.
Toyotamaphimeia taiwanicus (Shikama, Reference Shikama1972) new combination
Figures 2–4, 5.1–5.5
- Reference Shikama1972
Tomistoma (?) taiwanicus Shikama, p. 126, pl. 2, figs. 1–14, pl. 3, figs. 1–7, text-fig. 1.

Figure 2. Toyotamaphimeia taiwanicus n. comb. (1) NSMT-P-9121, an isolated tooth; (2–4) NSMT-P-9126, a middle section of the lower jaw: (2) posterior view; (3) dorsal view; (4) ventral view. White lines indicate the suture; dashed lines represent the proposed border. Left scale (1); right scale (2–4).

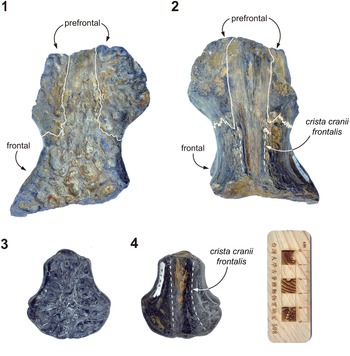
Figure 3. Toyotamaphimeia taiwanicus n. comb. (1, 2) NSMT-P-9122, a partial frontal/prefrontal complex: (1) dorsal view, (2) ventral view; (3, 4) NSMT-P-9123, a partial frontal (misidentified by Shikama, Reference Shikama1972, as the parietal): (3) dorsal view, (4) ventral view. White lines indicate sutures; dashed lines represent proposed borders.

Figure 4. Toyotamaphimeia taiwanicus n. comb. (1–4) NSMT-P-9124, a partial rostrum including the anterior portion of the maxilla and the dorsal posterior process of the premaxilla: (1) dorsal view, (2) ventral view, (3) left lateral view, and (4) right lateral view. White lines indicate sutures; dashed lines represent proposed borders.

Figure 5. Toyotamaphimeia taiwanicus n. comb. (1–5) NSMT-P-9125, a middle section of the lower jaw: (1, 2) dorsal view, (3) ventral view, (4, 5) medial view. (6) NSMT-H-5870, a left lower jaw of Tomistoma schlegelii in medial view. The red areas show the primordial canal in the lower jaw. Dotted areas indicate eroded regions. Black lines indicate sutures; dashed lines represent proposed borders.
Type materials
The original type specimens include six specimens: NSMT-P-9121, an isolated tooth (Fig. 2.4); NSMT-P-9122, a partial frontal/prefrontal complex (Fig. 3); NSMT-P-9123, originally identified as a partial parietal by Shikama (Reference Shikama1972), but here revised to be a partial frontal (see our discussion; Fig. 3); NSMT-P-9124, a partial rostrum, including the anterior portion of the maxilla and the dorsal posterior process of the premaxilla (Fig. 4); NSMT-P-9125, a middle section of the left lower jaw (Fig. 5.1–5.5) identified as the anterior segment of the lower jaw by Shikama (Reference Shikama1972); and NSMT-P-9126, a middle section of the lower jaw (Fig. 2.1–2.3). Our detailed re-examination confirms the existence of at least two individuals in the type specimens—two frontals, likely indicating the size disparity between a large adult (NSMT-P-9122) and a young juvenile (NSMT-P-9123). All six specimens are syntypes, and we refrain from designating a lectotype, for now, owing to the incomplete nature of the type specimens. The high-resolution 3D data of the NSMT materials are freely available at: https://doi.org/10.5281/zenodo.7340097 or https://scholars.lib.ntu.edu.tw/handle/123456789/625770.
Diagnosis
Our examination shows that the morphological combination seen in NSMT-P-9122 (frontal/prefrontal complex) corresponds to that of all known Toyotamaphimeia specimens (Figs. 6, 7), which differ from Tomistoma schlegelii (Muller, Reference Müller1838) (NSMT-H-5870) by having medially curving posterolateral margin of the prefrontal (Fig. 6) and a slightly, but not prominent, upturned orbital margin (Fig. 7). More importantly, there is a difference between the Tainan material and Toyotamaphimeia machikanensis in the position of the medial maxillary occlusal pits. The Tainan materials (both NSMT and the Waseda specimens) exhibit those pits near the anterior (2nd–7th) maxillary teeth (Fig. 8.1, 8.2), whereas in Toyotamaphimeia machikanensis these are positioned farther posteriorly, near the 8th–13th teeth (Fig. 8.3).

Figure 6. Comparison of the interorbital regions of studied specimens in dorsal view. (1) Toyotamaphimeia taiwanicus n. comb. (NSMT-P-9122). (2) Toyotamaphimeia machikanensis (NSMT-P-20510). (3) Toyotamaphimeia cf. T. machikanensis from Kishiwada (KSNHM-F7-6). (4) Penghusuchus pani (NMNS-005645). (5) Tomistoma schlegelii (NSMT-H-5870). Solid lines indicate sutures; dashed lines represent proposed borders. All scale bars = 5 cm.

Figure 7. Comparison of the orbital regions of studied specimens in anterior view. (1) Toyotamaphimeia taiwanicus n. comb. (NSMT-P-9122). (2) Toyotamaphimeia machikanensis (NSMT-P-20510). (3) Toyotamaphimeia cf. T. machikanensis from Kishiwada (KSNHM-F7-6). (4) Penghusuchus pani (NMNS-005645). (5) Tomistoma schlegelii (NSMT-H-5870). Dashed lines indicate the dorsal orbital margins. All scale bars = 5 cm.

Figure 8. Comparison of the occlusal pit positions and anterior rostral shapes (the portion of 1st–7th alveoli). (1) Toyotamaphimeia taiwanicus n. comb. (NSMT-P-9124). (2) Toyotamaphimeia taiwanicus n. comb. (WU-HPAC-D 007). (3) Toyotamaphimeia machikanensis (NSMT-P-20510). (4) Penghusuchus pani (NMNS-005645) (5) Hanyusuchus sinensis (SM S01812; from the supplementary material of Iijima et al., Reference Iijima, Qiao, Lin, Peng, Yoneda and Liu2022, under the license of CC-BY 4.0). White translucent areas indicate occlusal pit positions; dashed lines represent proposed borders. All scale bars = 10 cm.
Our morphological comparisons allow a reliable assignment of Tomistoma taiwanicus to Toyotamaphimeia, creating Toyotamaphimeia taiwanicus n. comb. Of note, based on the morphological similarity between the Waseda specimen (WU-HPAC-D 007) and the NSMT materials, we assign WU-HPAC-D 007 to Toyotamaphimeia cf. T. taiwanicus n. comb., because no definite morphological differences exist between WU-HPAC-D 007 and NSMT-P-9126, and both specimens likely come from the same geological horizon. In addition, both the rostral outlines of Toyotamaphimeia taiwanicus n. comb. represented by WU-HPAC-D 007 and Toyotamaphimeia machikanensis show a curved and prominent expansion at the level of 7th alveolus (Fig. 8), in contrast to the more gradual and moderate expansion of the rostral outlines of Hanyusuchus sinensis and Penghusuchus pani (Fig. 8). All the fossil materials described here (except NSMT-P-9123) likely belong to physically and sexually mature individuals (>4 m and up to 7 m long; Iijima and Kubo, Reference Iijima and Kubo2020; Iijima et al., Reference Iijima, Qiao, Lin, Peng, Yoneda and Liu2022), eliminating the likelihood of ontogenetic differences (Iijima, Reference Iijima2017).
Description of the type specimens
Both the NSMT and Waseda materials have been described in detail by Shikama (Reference Shikama1972) and Ito et al. (Reference Ito, Aoki, Hirayama, Yoshida, Kon and Endo2018), respectively. We aim to describe new morphological observations and conduct further comparisons with other relevant and newly discovered fossils. One of the most critical re-identifications is that NSMT-P-9123, originally recognized as the parietal by Shikama (Reference Shikama1972), is in fact a frontal, because of the presence of a narrow crista cranii frontalis in ventral view (Fig. 3.4). The posterolateral margin of the orbit in NSMT-P-9123 is circular, and its dorsal orbital margin is nearly smooth (Fig. 3.3), similar to that of NSMT-P-9122. The size disparity between NSMT-P-9122 and NSMT-P-9123 suggests that they represent a large individual and a young juvenile.
NSMT-P-9121 is an isolated tooth (Fig. 2.4). NSMT-P-9126, a middle section of the lower jaw, accommodates another partial tooth (Fig. 2.1, 2.3). Both teeth are large and robust, slightly curved, and with a blunt apex. Two carinae are seen on the tooth surface, forming an angle of almost 90° to its curving axis (Fig. 2.2). The tooth crown exhibits a series of minor striations. The overall morphology of the tooth is similar to the teeth of Toyotamaphimeia machikanensis and different from the elongated and strongly curving teeth with a sharp apex of the extant Tomistoma schlegelii.
The NSMT-P-9122 partial frontal/prefrontal complex (Fig. 3.1, 3.2) is large and comparable to that of the holotype of Toyotamaphimeia machikanensis. Shikama (Reference Shikama1972) oriented NSMT-P-9122 in a reverse anteroposterior direction and missed the existence of the prefrontal. Our re-examination confirms a brief notice by Shan et al. (Reference Shan, Wu, Cheng and Sato2009) that NSMT-P-9122 includes both the frontal and prefrontal. The inner orbital margin of NSMT-P-9122 is linear, whereas both the extant Tomistoma schlegelii and the Miocene Penghusuchus pani show an apparent “inflection point” at the level of the narrowest part of the interorbital (Fig. 6). The posterolateral margin of the prefrontal in NSMT-P-9122 is medially curved and forms the anteromedial margin of the orbit, making a circular margin along the inner orbit (Figs. 2.1, 6.1). This condition also occurs in Toyotamaphimeia machikanensis, Hanyusuchus sinensis, and Penghusuchus pani, but the inner orbital margin of Penghusuchus pani is more angular (Fig. 6). The posterolateral margin of the prefrontal is straight instead of medially curving, as in the extant Tomistoma schlegelii in which the general orbital shape is like a capital D (Fig. 6). In addition, the dorsal margin of the orbit is fairly smooth, similar to that of Toyotamaphimeia machikanensis, Hanyusuchus sinensis, and Penghusuchus pani, but different from the prominent upturned ridge of Tomistoma schlegelii (Fig. 7).
The dorsal surface of a partial rostrum (NSMT-P-9124; Fig. 4) is covered by a thin sediment layer, hampering identification of the premaxilla–maxilla. The suture is likely anteroposteriorly linear, gradually curving anterolaterally. The rostrum is lateromedially narrow, with the lateral margins of the anterior portion parallel in dorsal view with a gentle posterior expansion. Ventrally, no trace of premaxilla or palatine is preserved. The left maxilla preserves one intact alveolus in NSMT-P-9124, and the alveoli increase slightly in size posteriorly, along with a gradual lateral expansion of the maxilla. Prominent concavities along the lateral margin of the maxilla between consecutive alveoli represent occlusal pits for the lower dentition. The equal size of the alveoli and the parallel margins of the preserved maxilla suggest that NSMT-P-9124 is more likely an anterior-most portion of the maxilla preserving the 1st–3rd alveoli.
NSMT-P-9125, recognized as the anterior portion of the lower jaw by Shikama (Reference Shikama1972), seems to belong to the middle segment of the lower jaw, given the presence of a primordial canal extending horizontally along its dorsoventral center instead of being displaced dorsally or on the dorsal half (Fig. 5). The position of the primordial canal remains unknown in the Toyotamaphimeia lineage, but future finds and CT data should help to test if it is like the extant Tomistoma schlegelii (Fig. 5.6). Based on our close examination, the narrow anterior tip of NSMT-P-9125 likely results from post-mortem erosion and damage, making the overall morphology like the anterior part of the dentary. In addition, the size of the alveoli in both NSMT-P-9125 and NSMT-P-9126 is similar and shows no enlargement along the lower jaws. We also found no remains of the splenial in both specimens, suggesting that NSMT-P-9125 and NSMT-P-9126 belong to lower jaw sections anterior to the splenial symphysis and posterior to the enlarged 4th alveoli.
Referred specimen
WU-HPAC-D 007, a partial rostrum, including the anterior portion of the maxilla and an uncertain piece of the premaxilla. This historic specimen has long been considered to have been lost during World War II after a brief report (Tokunaga, Reference Tokunaga1936), but fortunately was rediscovered in the Waseda collection (Ito et al., Reference Ito, Aoki, Hirayama, Yoshida, Kon and Endo2018).
Body size
Based on the size estimation of Toyotamaphimeia machikanensis (Iijima and Kubo, Reference Iijima and Kubo2020), the body size of Toyotamaphimeia taiwanicus n. comb. is comparable (i.e., >6 m and up to 7 m long). The sizes of given anatomical parts of NSMT-P-9122, 9124, 9125, and 9126 are almost identical to those of the holotype of Toyotamaphimeia machikanensis. For example, the interorbital width is 49 mm in NMST-P-9122 and 49.6 mm in Toyotamaphimeia machikanensis. In addition, the much smaller size of NSMT-P-9123, re-identified here as a frontal, compared to NSMT-P-9122, plus our comparison with extant crocodylians, suggests that NSMT-P-9123 belongs to a juvenile individual likely ranging from 2–3 m long.
Remarks
The original diagnosis of “Tomistoma (?) taiwanicus” of Shikama (Reference Shikama1972, p. 126) is ambiguous and unfounded for the following reasons: (1) because preservation of the anterior-most maxilla NSMT-P-9124 is incomplete, we cannot be sure that its snout is “more elongate” than that of Toyotamaphimeia machikanensis; (2) NSMT-P-9124 represents the anterior portion of the upper jaw, showing the suture between the maxilla and premaxilla instead of occurring more posteriorly between the maxilla and nasal, hence we cannot confirm the presence of an “obsolete suture between the maxilla and nasal”; (3) the isolated tooth (NSMT-P-9121) and preserved alveoli of the rostrum fall within the size range of Toyotamaphimeia machikanensis teeth, and are not “relatively large”; (4) the diastema between consecutive teeth also are similar to those in Toyotamaphimeia machikanensis; (5) we found the crista cranii frontalis on the ventral side of NSMT-P-9123, indicating it is a frontal instead of a parietal, so that the “very narrow parietal” is an illegitimate diagnostic trait; (6) the distinctly angulated “inner margin of orbital foramen” seems a more probable diagnostic feature, but comparison with other tomistomines (Fig. 6) shows that the orbit of NSMT-P-9123 has a circular and smooth inner margin. In the end, our comprehensive re-examination clearly demonstrates that the previous diagnosis of “Tomistoma (?) taiwanicus” is rather problematic.
Phylogenetic results
In the first round of our cladistic analysis (including all characters), we recovered 710 most parsimonious trees (MPTs) with 974 steps (see details in the Supplementary material). The 50% majority-rule consensus of those trees (time-calibrated, after Nicholl et al., Reference Nicholl, Rio, Mannion and Delfino2020, and Iijima et al., Reference Iijima, Qiao, Lin, Peng, Yoneda and Liu2022; Fig. 9) shows Toyotamaphimeia taiwanicus n. comb. (NSMT materials + WU-HPAC-D 007) in a polytomy with two clades, one formed by Hanyusuchus sinensis plus an undescribed tomistomine (NTUM-VP 200702, currently under study by YYC and CHT), and one formed by the three Toyotamaphimeia specimens from Japan. The sister taxon of this polytomy is the Miocene Penghusuchus pani from Taiwan. The next more externally branching lineage includes two species of Thecachampsa from the Miocene of North America.

Figure 9. Phylogenetic assessment of Toyotamaphimeia taiwanicus n. comb. 50% majority tree under the all-character analysis (254 characters and 77 OTUs) with time-calibrated information after Nicholl et al. (Reference Nicholl, Rio, Mannion and Delfino2020) and Iijima et al. (Reference Iijima, Qiao, Lin, Peng, Yoneda and Liu2022). Toyotamaphimeia silhouette downloaded from phylopic.org and illustrated by Armin Reindl under the license CC BY-NC 3.0.
We conducted an additional skull-only analysis (inactivating the post-cranial characters) and recovered 2760 MPTs of 814 steps (see details in the Supplementary material). Their 50% majority-rule consensus tree shows polytomy with five lineages (Toyotamaphimeia taiwanicus n. comb., Hanyusuchus sinensis, Penghusuchus pani, NTUM-VP 200702, and a clade including the three Toyotamaphimeia from Japan). Surprisingly, this skull-only analysis generated a topology in which tomistomines are included within Gavialoidea, as reported in a more recent morphology-based phylogeny (Rio and Mannion, Reference Rio and Mannion2021).
Interestingly, both the all-character and skull-only analyses suggest the existence of an East Asian lineage, including the Miocene Penghusuchus pani (Taiwan), the Pleistocene Toyotamaphimeia taiwanicus n. comb. (Taiwan) and Toyotamaphimeia machikanensis (Japan), and the Holocene Hanyusuchus sinensis (southeastern China). Seven unambiguous synapomorphies support this lineage: (1) the largest maxillary alveolus in the “first wave” is the 7th (Cha. 89-7); (2) the dorsal premaxillary processes are long and extend beyond the 5th maxillary alveolus (Cha. 145-2); (3) the opening of the lateral eustachian canals is lateral to the medial eustachian canal (Cha. 147-1); (4) the surangular-articular suture is oriented anteroposteriorly (Cha. 162-0); (5) the ventral processes of the exoccipital are oriented lateroventrally in occipital view (Cha. 189-1); (6) the largest maxillary tooth in the second “wave” posterior to the 9th tooth is the 12th or a more posterior tooth (Cha. 227-1); and (7) the maxillary teeth are not widely spaced, with the 7th and 8th teeth not more spaced than other diastemas (Cha. 235-0).
Discussion
Our detailed morphological re-examination of the type specimens and phylogenetic analyses allow a confident reassignment of Tomistoma taiwanicus to Toyotamaphimeia taiwanicus n. comb., suggesting an East Asian, instead of Southeast Asian, influence for evolution of the Pleistocene crocodylians from Tainan, Taiwan. Similarly, our study also allowed assignment of the Waseda specimen to Toyotamaphimeia cf. T. taiwanicus n. comb. because it shares medial maxillary occlusal pits on the anterior part of the maxillary dentition (2nd–7th teeth) with the NSMT materials, rather than more posteriorly (8th–13th teeth), as in Toyotamaphimeia machikanensis. The Waseda specimen (WU-HPAC-D 007) and the NSMT materials (NSMT-P-9121, 9123, 9124, 9125, and 9126) firmly confirm the existence of Toyotamaphimeia in Taiwan.
Toyotamaphimeia taiwanicus n. comb. from the Pleistocene of Taiwan represents a tropical species of Toyotamaphimeia and provides new insights into a long-standing puzzle as to why and how Toyotamaphimeia machikanensis solely inhabited a relatively high-latitudinal region (Japan), given that crocodylian physiology generally conforms to low-latitudinal warmer areas. A mid-latitude occurrence is more likely, within the northern boundaries of the extant crocodylian distribution range (Iijima et al., Reference Iijima, Momohara, Kobayashi, Hayashi, Ikeda, Taruno, Watanabe, Tanimoto and Furui2018; de Celis et al., Reference de Celis, Narváez and Ortega2020), but not beyond 35° (Nicolaï and Matzke, Reference Nicolaï and Matzke2019). For instance, the fossil record of Gavialis gangeticus Gmelin, Reference Gmelin1789, indicates a farther northern distribution than today (Nanda et al., Reference Nanda, Schleich and Kotlia2016; Martin, Reference Martin2019). Similarly, the past distribution of Alligator sinensis Fauvel, Reference Fauvel1879, also ranges from Thailand to farther north in Japan (Iijima et al., Reference Iijima, Takahashi and Kobayashi2016). Crocodylus niloticus Laurenti, Reference Laurenti1768, is widely distributed across the entirety of Africa, including colder areas (Fergusson, Reference Fergusson, Manolis and Stevenson2010), and the extant largest Crocodylus porosus Schneider, Reference Schneider1801, also shows a wide distribution range (Webb et al., Reference Webb, Manolis, Brien, Manolis and Stevenson2010). This suggests a past distribution expansion to mid-latitude regions for Crocodylia (including the extant Alligatoridae, Crocodylidae, and Gavialidae). We then should expect to find a low-latitude member of the Toyotamaphimeia lineage, and our Toyotamaphimeia taiwanicus n. comb. from the tropical Pleistocene of Tainan resolves this paleogeographic conundrum.
Temperature is one of the most critical abiotic factors restricting crocodylian range and the distribution of crown-Crocodylia has been restricted to tropical or subtropical areas between 35°N and 35°S latitude after a global cooling event in the Late Miocene (Markwick, Reference Markwick1998a, Reference Markwickb; Mannion et al., Reference Mannion, Benson, Carrano, Tennant, Judd and Butler2015). The repeated glacial–interglacial cycles during the Pleistocene resulted in constant environmental change and forced large-scale dispersal events. Macro-plant fossils coeval with Toyotamaphimeia machikanensis suggest an environmental temperature close to the lowest boundary for crocodylian survival during the interglacial period in Japan, and the lineage may have dispersed farther south during the Pleistocene glaciations (Iijima et al., Reference Iijima, Momohara, Kobayashi, Hayashi, Ikeda, Taruno, Watanabe, Tanimoto and Furui2018). However, our recognition of Toyotamaphimeia, which is larger and geologically older than Toyotamaphimeia machikanensis (T. taiwanicus n. comb.: 0.8–0.4 Ma; T. machikanensis: 0.6–0.5 Ma), in a Pleistocene tropical site (Tainan, Taiwan), provides a novel interpretation. We propose that Toyotamaphimeia originated in Taiwan and evolved to a large body size (i.e., reaching 7 m long), with a thermoregulation likely similar to that of the “saltwater crocodile” Crocodylus porosus. This is the largest extant crocodylian (>6 m long) with a gigantothermic physiology that allows maintaining its body temperature under cold conditions (Grigg et al., Reference Grigg, Seebacherd, Beard and Morris1998; Seebacher et al., Reference Seebacher, Grigg and Beard1999). The gigantic Toyotamaphimeia taiwanicus n. comb. also may have been equipped with a gigantothermic physiology, allowing an out-of-Taiwan migration, dispersing to higher latitudinal regions, such as Japan.
Surprisingly, both our all-character and skull-only phylogenetic analyses generated a consistent “East Asian tomistomine clade,” including Penghusuchus pani (Taiwan), Toyotamaphimeia machikanensis (Japan), Toyotamaphimeia taiwanicus n. comb. (Taiwan), Hanyusuchus sinensis (southeastern China), and an undescribed specimen (NTUM-VP 200702, Taiwan), but under disparate large-scale phylogenetic scenarios—tomistomines nested either in the crocodyloid or in the gavialoid clade (Fig. 9; see Supplementary material for all topologies). This emphasizes the phylogenetic instability with different taxa and character data, highlighting the gharial problem. A recent extensive morphology-based phylogeny (Rio and Mannion, Reference Rio and Mannion2021) recovered a tomistomine lineage nested within gavialoids, as in our skull-only analysis. Nevertheless, Penghusuchus pani and Toyotamaphimeia form a ladder topology towards the extant Gavialis (Rio and Mannion, Reference Rio and Mannion2021) rather than an “East Asian tomistomine clade,” as both of our analyses suggest. More fossil crocodylians with well-preserved materials and critical assessment of character descriptions should be key to resolving such phylogenetic inconsistencies. In addition, detailed description and phylogenetic assessment of the undescribed NTMU-VP 200702 (or more-complete specimens of Toyotamaphimeia taiwanicus n. comb. and Penghusuchus pani) from Taiwan should help illuminate hidden diversity, perhaps revealing a more robust phylogenetic structure.
Interestingly, our re-examination of Toyotamaphimeia taiwanicus n. comb. resulted in the discovery of size disparity between individuals present among the type specimens. NSMT-P-9122 represents a large adult reaching ~7 m long, similar in size to the holotype of Toyotamaphimeia machikanensis, whereas NSMT-P-9123 is a younger juvenile, likely 2–3 m long. Prior to our present study, two crocodylian lineages were reported from the same locality and horizon (Chiting Formation of Tainan): Tomistoma taiwanicus Shikama, Reference Shikama1972, and Toyotamaphimeia sp. (Ito et al., Reference Ito, Aoki, Hirayama, Yoshida, Kon and Endo2018). After examining all relevant specimens (NSMT and Waseda materials), we conclude that they all belong to Toyotamaphimeia taiwanicus n. comb. However, given that the oldest crocodylian (the tomistomine Penghusuchus pani) from Taiwan can be traced back to the Middle Miocene, a much higher crocodylian or tomistomine diversity may be anticipated for the Pleistocene of Tainan, because coexistence and niche partitioning among crocodylians are fairly common (Young et al., Reference Young, Bell, de Andrade and Brusatte2011; Salas-Gismondi et al., Reference Salas-Gismondi, Flynn, Baby, Tejada-Lara, Wesselingh and Antoine2015; Staniewicz et al., Reference Staniewicz, Behler, Dharmasyah and Jones2018).
Causes for the complete extinction of crocodylians in Taiwan have never been discussed or explored. Our discussion here, based on the updated crocodylian taxonomy from Taiwan, lays a foundation for future endeavors. Environmental changes have been linked to the crocodylian extinction (e.g., Cosette et al., Reference Cossette, Adams, Drumheller, Nestler, Benefit, McCrossin, Manthi, Juma and Brochu2020; Hocknull et al., Reference Hocknull, Lewis, Arnold, Pietsch and Joannes-Boyau2020; Solórzano et al., Reference Solórzano, Núñez-Flores, Inostroza-Michael and Hernández2020), but the paleoecological aspects associated with crocodylian fossils in Taiwan remain virtually unknown, and this again highlights the long-ignored vertebrate paleontology of the country. For example, here we revised the taxonomy of historic crocodylian specimens—the first and the second fossil crocodylians in the history of Taiwan—which are properly curated in the NSMT and Waseda collections in Japan, making our study possible. On the contrary, some historic specimens from Taiwan should have been curated for future research, but the original fossils are lost, likely owing to the indifference to paleontological research (e.g., Liaw and Tsai, Reference Liaw and Tsai2022; Tsai and Tseng, Reference Tsai and Tseng2022). Our current efforts into the development of vertebrate paleontology in Taiwan aims to better understand the Pleistocene paleo-ecosystems and to further elucidate the driving factors of extinctions in the island. For instance, the coexistence of the huge Toyotamaphimeia and the carnivorous, machairodontine felid Homotherium (Tsai and Tseng, Reference Tsai and Tseng2022) in Tainan, should draw far-ranging attention and lead to more in-depth paleontological research in Taiwan.
Acknowledgments
We thank M. Manabe and T. Tsuihiji (NSMT, Japan; now known as National Museum of Nature and Science, Japan) for allowing us to borrow the NSMT type specimens and access the replicas (NSMT-P-20510) of MOUF00001; S. Kawada (NSMT) for access to extant an Tomistoma (NSMT-H-5870) specimen; R. Hirayama (Waseda University) for access to WU-HPAC-D 007; S. Hirata, K. Watanabe, T. Masahiro, and H. Tokugawa (Natural History Museum, Kishiwada City, Japan) for access to Toyotamaphimeia cf. T. machikanensis (KSNHM-F7-6); S.-W. Wang and H.-Y. Shan (National Museum of Natural Science, Taiwan) for access to Penghusuchus pani (NMNS-005645); M. Iijima and K. Watanabe for providing high-resolution photos of MOUF00001 and the Yage specimen (SPMN uncatalogued material, Museum of Natural and Environmental History, Shizuoka, Japan); I.-L. Lu and his 3DR INTERNATIONAL LTD for 3D scanning the NSMT materials; and the handling editor M. Langer and two anonymous reviewers for constructive suggestions and comments. This research was funded by the Taiwan Ministry of Science and Technology (MOST 107-2621-B-002-005, 108-2621-B-002-006-MY3, 111-2621-B-002-006; now known as National Science and Technology Council, Taiwan) and by public donations to the Lab of Evolution and Diversity of Fossil Vertebrates, National Taiwan University (NTU FD107028) to CHT. This research was also partly supported by National Taiwan Museum.
Author contributions
CHT conceived and designed this study; YYC and CHT collected the data, and YYC analyzed the dataset; both authors wrote and discussed the content of the manuscript; both authors approved the final submission for publication.
Declaration of competing interests
The authors declare no competing interests.
Data availability statement
Data available from the Zenodo Digital Repository: https://doi.org/10.5281/zenodo.7340097 and http://doi.org/10.5281/zenodo.7937137.










 Open access
Open access