



1. Arumberia: a globally recognized sedimentary surface texture
Arumberia is a sedimentary surface texture comprising a series of low-relief, sharply defined sub-parallel lines (Fig. 1), which is often recognized on Precambrian and lower Palaeozoic bedding planes. It was originally reported from the Ediacaran Arumbera Sandstone of Australia by Glaessner & Walter (Reference Glaessner and Walter1975), who interpreted it as an impression fossil of a small cup-shaped metazoan. Subsequent Arumberia discoveries have shown that the cup-shaped gross morphology of the original specimens is not indicative of its true form, which can extend over multiple square metres on planar or rippled bedding planes (Fig. 1i; Table 1). The characteristic features of Arumberia have been explained in several mutually exclusive ways. Dominant among these are hypotheses that the texture is either the ‘body-fossil’ impression of a mat-like organism or community attached to a sedimentary substrate (Bland, Reference Bland1984; Kumar & Pandey, Reference Kumar and Pandey2008, Reference Kumar and Pandey2009; Kolesnikov et al. Reference Kolesnikov, Grazhdankin and Maslov2012, Reference Kolesnikov, Marusin, Nagovitsin, Maslov and Grazhdankin2015; Arrouy et al. Reference Arrouy, Warren, Quaglio, Poiré, Simões, Rosa and Peral2016), or a physical sedimentary structure formed with or without a microbial mat covering (Brasier, Reference Brasier and House1979; McIlroy & Walter, Reference McIlroy and Walter1997; Jensen et al. Reference Jensen, Droser and Gehling2005; McIlroy et al. Reference McIlroy, Crimes and Pauley2005; Allen & Leather, Reference Allen and Leather2006; Seilacher, Reference Seilacher2007; Netto, Reference Netto, Netto, Carmona and Tognoli2012; Liu et al. Reference Liu, Brasier, Bogolepova, Raevskaya and Gubanov2013; Sharma & Mathur, Reference Sharma, Mathur, Shrivastava and Kumar2014; Becker-Kerber et al. Reference Becker-Kerber, Paim, Junior, Girelli, da Rosa, El Albani, Osés, Prado, Figueiredo, Simões and Pacheco2020). Notable among previous work is the 1984 review paper by Bland, which described and reviewed all prior known Arumberia occurrences. Bland concluded that only a fraction of reported Arumberia specimens exhibited the original diagnostic criteria of Glaessner & Walter (Reference Glaessner and Walter1975), and that they represented large impression fossils restricted to uppermost Precambrian and lower Cambrian strata deposited in very shallow-marine to non-marine environments.

Fig. 1. Global Arumberia examples. (a) Maturin Ponds Formation, Canada. Note frequent bifurcations. (b) Synalds Formation, United Kingdom. Note the lack of intermittent bifurcations. Coin has diameter of 19 mm. (c) Ferryland Head Formation, Canada. (d) Maturin Ponds Formation, Canada. Coin has diameter of 16.25 mm. (e) Synalds Formation, United Kingdom. (f) Crown Hill Formation, Canada. In this instance Arumberia lines follow the direction indicated by the underlying linguoid ripple marks. Coin has diameter of 16.25 mm. (g) Arumbera Sandstone, Australia. (h) Maturin Ponds Formation, Canada. White arrow highlights an instance where multiple fine, superimposed, Arumberia cords are observed to comprise an ‘individual’ Arumberia line. (i) Ferryland Head Formation, Canada, an instance where contiguous Arumberia lines extend across a large surface area (exceeding 3 m2). Contiguous Arumberia examples rule out the original hypothesis that Arumberia records the remains of a small cup-shaped organism (Glaessner & Walter, Reference Glaessner and Walter1975). (j) Crown Hill Formation. Arumberia are overprinted by raindrop marks (implying Arumberia was an ineffective buffer against droplet impact; Davies et al. Reference Davies, Liu, Gibling and Miller2016). (k) Maturin Ponds Formation. (l) Gibbett Hill Formation, Canada. (m) Crown Hill Formation, Canada. (n) Ferryland Head Formation. (o) Crown Hill Formation, Canada. (p) Maturin Ponds Formation, Canada.
Table 1. Details of Arumberia morphologies documented by the present authors (in bold) or by previous workers (listed in Table 2). We searched for Arumberia in the Erquy Formation, Lightspout Formation, Rozel Conglomerate and Portway Formation, but without success. Features previously reported as Arumberia in the Diabaig Formation have been studied by us and are not considered to be equivalent (Section 5). Epi – epirelief; Hypo – hyporelief; +/− – positive/negative.

In this study we critically assess the nature of Arumberia using new evidence from the largest reported instance so far discovered in the global rock record: a 300 m2 bedding plane in the lower Cambrian Port Lazo Formation of Brittany, France (Fig. 2). Isolated Arumberia have previously been reported from this locality (Bland, Reference Bland1984), but the extensive low-lying coastal outcrop we describe here was fortuitously exhumed during an interval when the modern beach profile was lowered. Supporting this case study, we revisit Bland’s (Reference Bland1984) assessment of Arumberia in light of 22 subsequent reports, in addition to the 18 instances that he previously reviewed (Tables 1, 2). Our literature survey has been ground-truthed by field visits to six of these Arumberia-bearing formations (Table 1; the Arumbera Sandstone of Northern Territory, Australia; the Crown Hill, Ferryland Head, Gibbett Hill and Maturin Ponds formations of Newfoundland, Canada; and the Synalds Formation of Shropshire, England). These sites have provided further observations to supplement evidence from the Port Lazo bedding plane and support our interpretation of Arumberia.
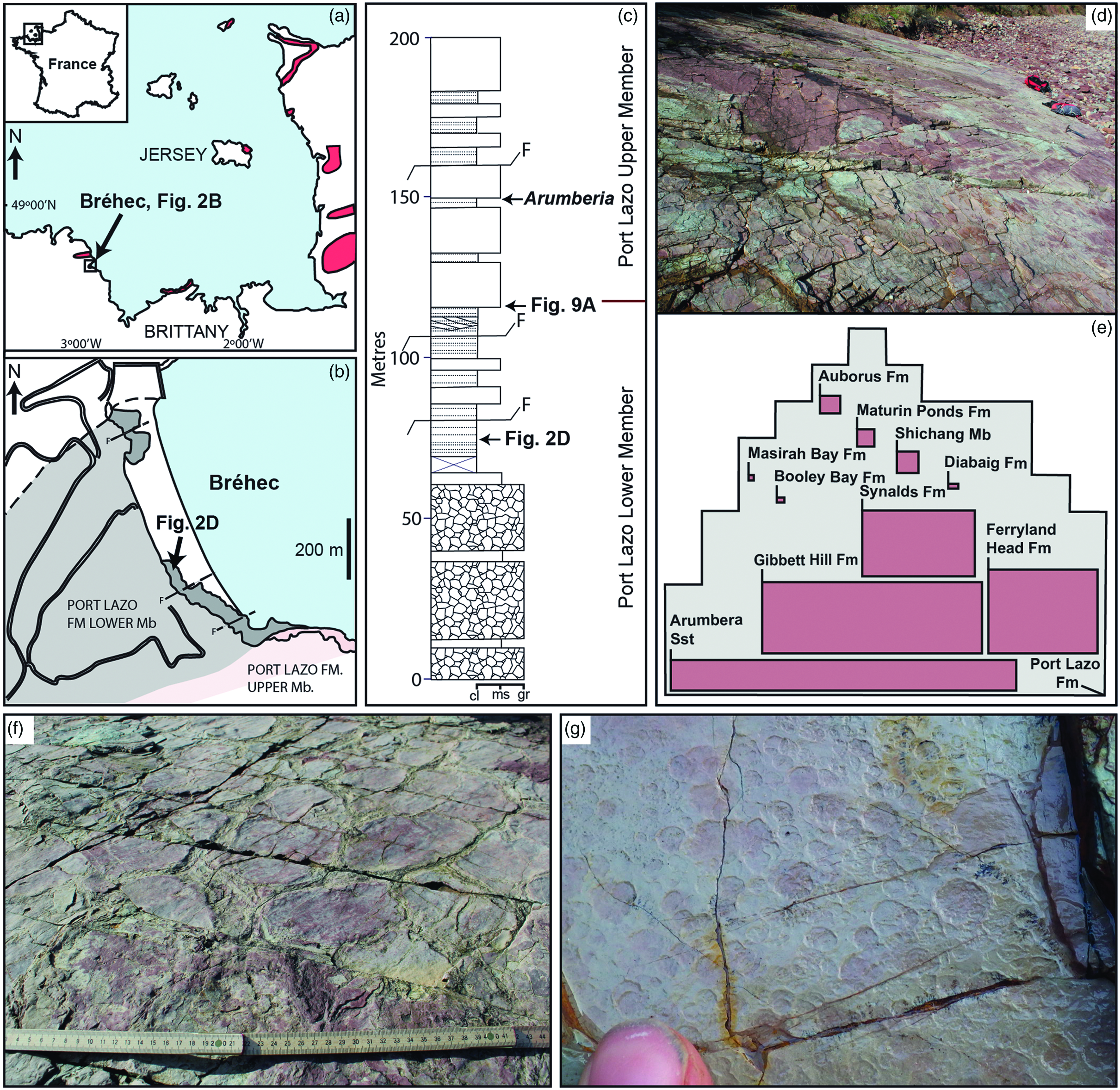
Fig. 2. Geographic, stratigraphic and spatial occurrence of Arumberia in the Port Lazo Formation, France. (a) Red outline marks outcrop locations of the Series Rouge (McMahon et al. Reference McMahon, Davies and Went2017; Went, Reference Went2021) that hosts the Arumberia-bearing Port Lazo Formation. (b) Location of Arumberia bedding plane at Bréhec Bay (amended from Went, Reference Went2017). (c) Measured section through the Port Lazo Formation indicating the position of the studied Arumberia-bedding plane. Cl – clay; ms – medium-grained sandstone; gr – granular sandstone/pebbly conglomerate (amended from Went, Reference Went2017). (d) The Arumberia-bearing bedding plane located at Bréhec Bay. (e) Grey outline illustrates the greater dimension of the analysed Port Lazo bedding plane in comparison to other previous Arumberia reports (drawn to scale, as red rectangles inset to the mapped grey outline). The total size of the Port Lazo bedding plane is c. 300 m2, but the graphic illustration is limited to the spatial coverage (86 m2) within which Arumberia was subjected by the authors to detailed scrutiny (measurements of line width, length, spacing, sinuosity and orientation). References detailing bedding plane dimensions of other Arumberia locations are listed in Table 2. (f) Desiccation cracks and (g) circular raindrop impressions identified on the studied red mudstone bed.
Table 2. Geographic and stratigraphic distribution of globally recognized Arumberia forms.

aAge assumed by the original authors based on presence of Arumberia.
2. Existing challenges and purpose of this paper
Universal diagnostic criteria for Arumberia have proved challenging to isolate (Glaessner & Walter, Reference Glaessner and Walter1975; Bland, Reference Bland1984; McIlroy et al. Reference McIlroy, Crimes and Pauley2005; Kumar & Pandey, Reference Kumar and Pandey2008, Reference Kumar and Pandey2009; Kolesnikov et al. Reference Kolesnikov, Grazhdankin and Maslov2012; Retallack & Broz, Reference Retallack and Broz2020), resulting in continuing uncertainty surrounding its affinities and origin (Table 3). Three factors have hampered understanding, described in the following.
-
(1) Preservation and ease of observation: it is challenging to describe individual specimens of Arumberia because the lateral margins of the texture are commonly either indistinct, or incomplete at the scale of observation. This makes it difficult to accurately define its areal extent. Additionally, while sub-parallel to parallel lines constitute all reported Arumberia (Fig. 1), the morphology and relief of these can be highly variable even over short distances, with spatially patchy expressions of branching, spacing, curvature and reticulation. Compounding these practical issues of description, Arumberia frequently has a topographic relief of < 1 mm, meaning that its visibility in natural outcrops can be hampered by poor light conditions, a lack of magnification and degradation through weathering.
-
(2) Changing diagnostic criteria and loaded terminology: the original type material of Arumberia (Glaessner & Walter, Reference Glaessner and Walter1975, redescribed by Retallack & Broz, Reference Retallack and Broz2020) lacks the full range of characteristics and spatial extents that have been revealed and incorporated into taxonomic diagnoses by later discoveries. Furthermore, some characteristics originally thought to be universal (e.g. conical forms with blunt apices, regular radial patterns of lines) are now known to be ancillary features, present in only a subset of instances. Arumberia has been reinterpreted several times as a body fossil (e.g. Retallack & Broz, Reference Retallack and Broz2020) or a pseudofossil (e.g. McIlroy et al. Reference McIlroy, Crimes and Pauley2005), with resultant emended diagnoses changing to reflect those interpretations through the addition or removal of diagnostic features. As a result, some of these criteria employ terminology that is loaded with an implication of biogenicity, rather than passive descriptive terms (e.g. description of the constituent lines as ‘rugae’; Kolesnikov et al. Reference Kolesnikov, Marusin, Nagovitsin, Maslov and Grazhdankin2015, Reference Kolesnikov, Danelian, Gommeaux, Maslov and Grazhdankin2017; McMahon et al. Reference McMahon, Davies and Went2017), further hindering objective interpretation.
-
(3) ‘Arumberia’ as a bucket-term: the most frequently observed characteristic of Arumberia is a set of closely spaced sub-parallel to parallel lines on a bedding plane (Fig. 1). Interpreting such a simple form in isolation encounters the challenge of equifinality: such phenomena may have plural potential explanations, both biotic and abiotic, and the cause behind a specific feature may be underdetermined by available evidence (e.g. Davies et al. Reference Davies, Liu, Gibling and Miller2016, Reference Davies, Shillito, Slater, Liu and McMahon2020). There appear to be instances in existing literature where the term ‘Arumberia’ has been liberally used to describe any observations of (sub)parallel lines: in some of these instances, the forms share no further characteristics in common with other described Arumberia (e.g. in terms of physical dimensions, accessory characteristics or host facies) (Table 1).
Table 3. Previous interpretations of Arumberia.

Below, we attempt to redress these outstanding issues by presenting observations of exceptionally extensive Arumberia from a newly discovered bedding plane in the lower Cambrian Port Lazo Formation in France, augmented with observations of well-preserved examples from elsewhere in the rock record. We use these observations to amend the diagnostic criteria of Glaessner & Walter (Reference Glaessner and Walter1975) in light of those characteristics that are universal, common or infrequent attributes of Arumberia, and highlight previously reported instances of ‘Arumberia’ that can be considered doubtful. Our reassessment shows that the least equivocal instances of Arumberia exhibit a narrow stratigraphic and palaeoenvironmental range (cf. Bland, Reference Bland1984).
3. Arumberia in the Port Lazo Formation
Previous reports of Arumberia from the Port Lazo Formation in Brittany, France (Bland, Reference Bland1984; Davies et al. Reference Davies, Liu, Gibling and Miller2016; McMahon et al. Reference McMahon, Davies and Went2017; Went, Reference Went2017, Reference Went2021) have not described the feature in detail. The Port Lazo Formation is a constituent of the ‘Series Rouge’, a predominantly red-bed succession that crops out in coastal exposures across NW France and the neighbouring Channel Islands (Fig. 2a) (McMahon et al. Reference McMahon, Davies and Went2017). Multiple potential ages have been suggested for the Port Lazo Formation. The French Geological Survey maps it as Early Ordovician in age, based on correlation to stratigraphy at Crozon that lies 125 km to the south across the North Armorican Shear Zone in the central Armorican domain (Egal et al. Reference Egal, Le Goff, Guennoc, Lebret, Thiéblemont, Hallégouët, Houlgatte, Callier and Carn1995). This correlation is consistent with the Port Lazo Formation being bounded by underlying Brioverian metasediments, which have recently yielded zircon U–Pb radiometric dates of 551 ± 7 Ma and 540 ± 5 Ma (Gougeon et al. Reference Gougeon, Néraudeau, Loi and Poujol2021), and a 472 ± 5 Ma age from overlying andesites (Fig. 2a) (Auvray et al. Reference Auvray, Mace, Vidal and Vander Voo1980). However, regional variability in tectonic setting and sedimentary environments favours an early Cambrian age for the Port Lazo Formation, with deposition having commenced in extensional basins shortly after the Cadomian orogenic climax c. 540 Ma (Peucat, Reference Peucat1986). Three lines of evidence support this contention. (1) The strata had a source area to the west (McMahon et al. Reference McMahon, Davies and Went2017), which is consistent with regional palaeogeography during the Cambrian Period (Went & McMahon, Reference Went and McMahon2018; Went, Reference Went2021), but unlikely during the Early Ordovician Epoch when surrounding areas witnessed the deposition of the well dated marine ‘Grès Armoricain’ (Paris et al. Reference Paris, Robardet, Dabard, Feist, Ghienne, Guillocheau, Le Hérissé, Loi, Mélou, Servais, Shergold, Vidal and Vizcaïno1999; Dabard et al. Reference Dabard, Loi and Paris2007). (2) Red-bed sequences in Normandy, which may be correlated with the Port Lazo Formation (Went, Reference Went2021), underlie Cambrian Stage 3 (521–514 Ma) limestones (dated by the presence of the trilobite Bigotina Pillola, Reference Pillola1993) and Cambrian Stage 2 (529–521 Ma) siliciclastic strata. (3) Radiometric ages from overlying andesites (Auvray et al. Reference Auvray, Mace, Vidal and Vander Voo1980), which provide crucial support for a posited Ordovician age, are reliant on the igneous bodies being extrusive; however, the andesites have more recently been demonstrated to be a mixture of disconformable lavas and intrusive bodies (Went, Reference Went2017). Furthermore, the presence of Arumberia itself may provide circumstantial evidence for a Cambrian age once the global stratigraphic record of this feature is considered (see Section 5).
The majority of Series Rouge outcrops are exposed within the limbs of basin-wide synclines (Fig. 2a), but the studied section of the Port Lazo Formation occurs as an outlier to these, cropping out at Bréhec Bay (Fig. 2b). Sedimentological observations made over the course of this study, and previously, have shown that Port Lazo Formation specimens of Arumberia are restricted to very shallow-water facies that were intermittently subject to sub-aerial exposure (Davies et al. Reference Davies, Liu, Gibling and Miller2016; McMahon et al. Reference McMahon, Davies and Went2017; Went, Reference Went2017, Reference Went2021).
The Port Lazo Formation is separated into two distinct members (Went, Reference Went2017), with the bedding plane central to this study occurring in the Port Lazo Lower Member (Fig. 2c, d). The surface has a pervasive covering of polygonal mud cracks (Fig. 2f), in addition to raindrop impressions and wave-ripple marks (Fig. 2g). These characteristics indicate that Arumberia occurs on a true substrate, that is, a bedding plane that faithfully records a synoptic topography that formed at the interface of sediment and water or air (Davies & Shillito, Reference Davies and Shillito2018, Reference Davies and Shillito2021). Its c. 300 m2 surface area provides an opportunity to assess Arumberia morphology across a far larger area than has previously proved possible (Fig. 2e). Approximately 50% of the bedding plane is covered by Arumberia (although contiguous individual Arumberia are likely less extensive than this, due to their occurrence within multiple successive < 1 mm thick clay laminae that are impractical to correlate across the entire surface). Additional surfaces bearing Arumberia are also present at higher stratigraphic levels in the Port Lazo Formation Upper Member (Fig. 2c). These examples, which are far more limited in spatial extent, were preserved in shallow subaqueous, nearshore marine settings (Went, Reference Went2017; Table 2).
3.a. Arumberia lines
The Port Lazo Arumberia consist of a series of parallel or sub-parallel lines, usually < 2 mm in relief, 0.5–1.5 mm wide and spaced 1–3 mm apart (Fig. 3). Line relief and spacing can be variable even across individual specimens. The lines are most frequently low sinuosity and can exhibit intermittent bifurcations (Fig. 3a–d, g, h). A subordinate expression is present where Arumberia specimens are radially arranged, curving gently away from a single apex through an angle of up to 40° (Fig. 3e, f). Arumberia lines can sometimes be seen to transition along their length into a reticulated pattern (Fig. 3i, j). Reticulated patterns cover areas of up to 30 cm2 and consist of intersecting lines that form irregularly shaped, sharp-edged polygons < 1 cm in diameter.

Fig. 3. Arumberia from the Port Lazo Formation. (a) Arumberia lines with intermittent bifurcations and reliefs of < 1 mm. (b) Arumberia lines with reliefs of approximately 2 mm. (c) Arumberia lines superimposed on a desiccated plate margin (white arrow). Black arrow depicts small ruptured domes shown in Figure 5f. (d) Arumberia lines in the Port Lazo Formation Upper Member. (e) Gently curved Arumberia that likely meet at a central node in the subcrop. Coin is 23.25 mm in diameter. (f) White arrow marks the approximate apex from which Arumberia lines in the bottom right of the image radiate. (g) Arumberia lines cross-cutting multiple desiccated polygons with no apparent deformation. (h) Arumberia positive epirelief lines superimposed by millimetre-wide negative epirelief ‘dimples’ (Section 3.c). Coin is 23.25 mm in diameter. (i) Sub-parallel Arumberia lines transition laterally into a reticulated pattern. No change in line relief or thickness occurs across the transition. (j) Line drawing of Figure 3i.
Arumberia lines frequently exhibit three-dimensionality to their preservation, implying that they originally had a cord-like form (in agreement with Bland, Reference Bland1984; Becker-Kerber et al. Reference Becker-Kerber, El Albani, Konhauser, Abd Elmola, Fontaine, Paim, Mazurier, Prado, Galante, Kerber, da Rosa, Fairchild, Meunier and Pacheco2021; see also Fig. 1h). On bedding surfaces, lines can occur in both positive epirelief (ridges) and negative epirelief (grooves), and both expressions of this relief can be seen along an individual line (a ‘ridge–groove’ transition) (Fig. 4). With magnification, they can be seen to be an artefact of partial weathering: the ridges record positive epirelief on the upper surface of the line-hosting lamina, and the grooves reflect casts of positive hyporelief in an underlying lamina (Fig. 4b). Ridge–groove transitions may reflect instances where Arumberia lines have been preserved in full relief between successive, very thin (c. 0.3 mm) clay laminae, or they may reflect moulding by sub-millimetre-thick laminae. In order to preserve the original three-dimensional (3D) morphology, Arumberia colonized the substrate during a net interval of sedimentary stasis, with subsequent deposition (in this instance of clay) covering Arumberia’s external surface, preceded by little or no scour (e.g. Davies & Shillito, Reference Davies and Shillito2021).

Fig. 4. 3D form of Arumberia in the Port Lazo Formation. (a) Clay veneers containing both positive epirelief lines on the top and positive hyporelief lines on the bottom. The hyporelief lines compress negative epirelief forms into the underlying bed, and changes in relief along individual lines occur at points where clay laminae have eroded from the outcrop. (b) Line drawing of Figure 4a. (c) Schematic depicting observed relief of 3D Arumberia ‘cords’ and the terminology used in this manuscript.
The three-dimensionality of the lines, and transitions from directed linearity to reticulated patterns, contradict previous assertions that Arumberia could be a physical sedimentary structure arising from unidirectional flow (i.e. a surficial texture registered on the substrate, such as current rills or flute marks: Brasier, Reference Brasier and House1979; Jenkins et al. Reference Jenkins, Plummer and Moriarty1981; McIlroy & Walter, Reference McIlroy and Walter1997; Seilacher, Reference Seilacher2007). Instead, these combined characteristics indicate the prior existence of a feature that was external to the sediment in which it is now hosted, and which responded variably along its length. The reticulation bears similarities to the patterning that emerges when buoyant filaments of bacteria or algae become entangled in standing or draining water (Shepard & Sumner, Reference Shepard and Sumner2010; Davies et al. Reference Davies, Liu, Gibling and Miller2016; Cuadrado & Pan, Reference Cuadrado and Pan2018).
On the large bedding plane exhibiting Arumberia (Fig. 2d), the drainage of water and associated drying and emergence is indicated by the presence of polygonal mud cracks and raindrop impressions (Fig. 2f, g). The occurrence of raindrop impressions overprinted on Arumberia implies that Arumberia was an ineffective buffer against droplet impact (Davies et al. Reference Davies, Liu, Gibling and Miller2016) (see also Fig. 1j). When in proximity to a desiccated plate margin, Arumberia can be seen to have deformed alongside the shrinking cohesive muds, and individual lines are seen to curl to form irregular, broadly concentric circles (Fig. 5). These instances demonstrate that Arumberia was already present on the muddy surface before desiccation. Small positive epirelief ‘blisters’ were observed at the margins of the deformed Arumberia (Fig. 5f), which potentially also formed during emergence as water escaped from the surficial clay veneer or a microbial mat, producing transient bubbles that subsequently ruptured at the surface. That Arumberia ridges and grooves document discrete 3D forms is further supported by late Ediacaran specimens from Newfoundland, Canada, in which individual Arumberia lines are seen to consist of multiple, finer lines that can appear to be superimposed upon one another (Fig. 1d, h).
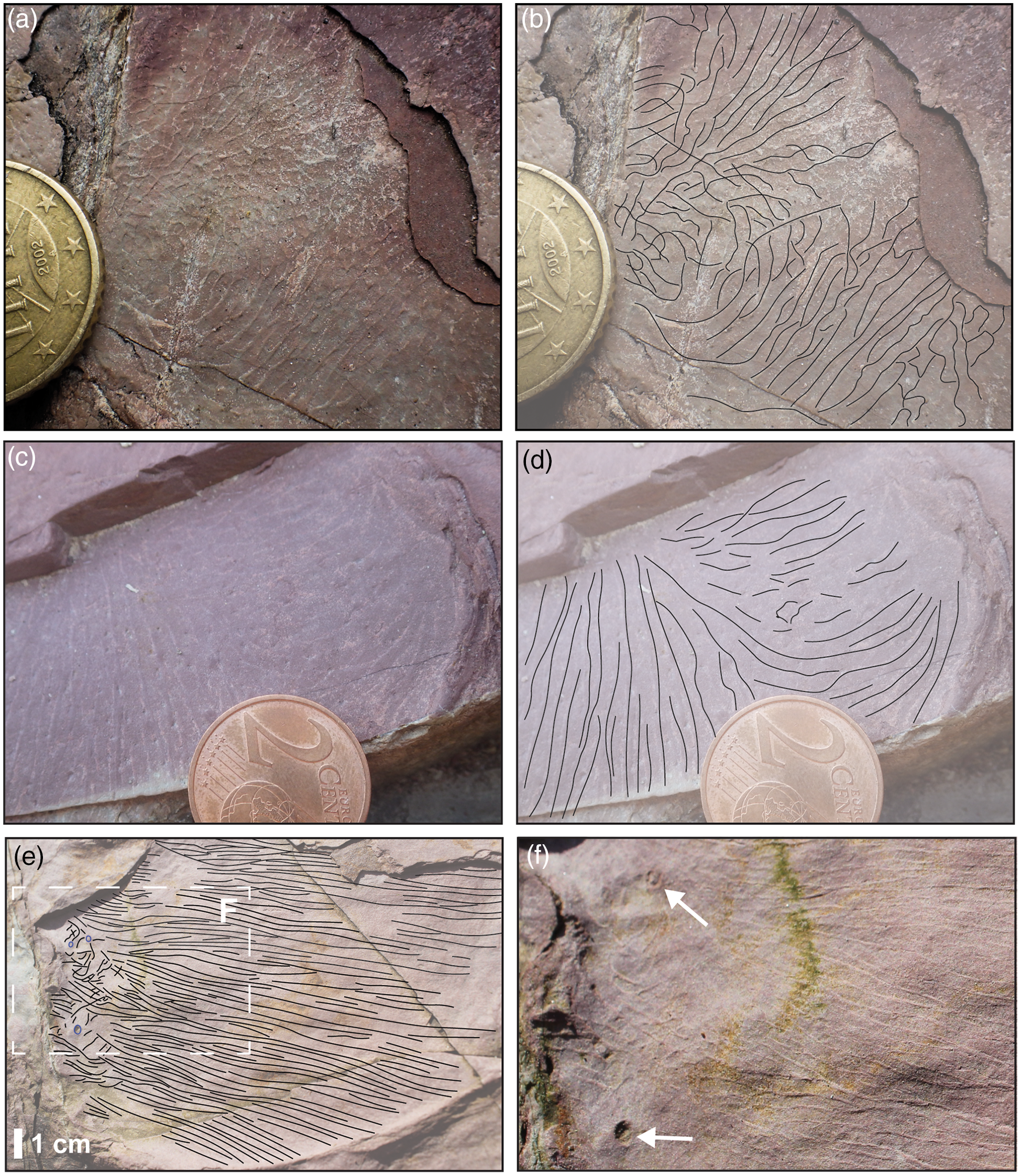
Fig. 5. Response to emergence displayed on Arumberia-bearing bedding planes in the Port Lazo Formation Lower Member. (a, c) Curled Arumberia lines in proximity to and in contact with desiccated plate margins. (b, d) Line drawings of Figures 5a and c, respectively. (e) Line drawing of linear Arumberia lines transitioning laterally into curled, discontinuous forms in proximity to a desiccated plate margin (original figure shown in Fig. 3c). (f) Inset of Figure 5e showing possible ruptured domes (‘blister structures’) occurring alongside deformed Arumberia adjacent to a desiccated plate margin. Similar discoidal ring-shaped bulges, no more than 30 mm in diameter and 3 mm in height, are present across the bedding plane. Coin in Figure 5a is 22.25 mm in diameter. Coin in Figure 5c is 18.75 mm in diameter.
3.b. Arumberia orientation
Directional measurements of Arumberia lines have a tight spread of WNW–ESE orientations over the total area of 300 m2 and through < 0.1 m of vertical succession (Fig. 6). Palaeocurrent measurements taken throughout the succession indicate that dominant current flow was near perpendicular to this axial orientation (n = 61), and symmetrical ripple mark crest lines on overlying beds have near-identical strikes (n = 14). The evident spatial arrangement of Arumberia lines with respect to measured palaeoflow implies a hydrodynamic role in morphogenesis.

Fig. 6. Map of Arumberia line orientations on a 300 m2 bedding plane in the Port Lazo Formation. Rose diagrams indicate predominant palaeoflow is approximately perpendicular to line orientation. Total mapped area 86 m2.
3.c. Association with dimple-pimple marks
Arumberia lines may be associated with 0.5–1.5 mm diameter circular features that lack internal structure (Figs 3d, h, 7). These features have reliefs of < 1 mm and can occur in dense populations consisting of hundreds of examples. They are generally found in negative epirelief (dimples) and positive epirelief (pimples) within < 1 mm thick clay veneers (reflecting full relief orbs, in the same way that Arumberia lines reflect full relief cords). One surface shows surficial dimples and basal pimples, effectively recording impression of clay laminae from above and below by two opposing hemispheres (Fig. 7d). Elsewhere, at least 10 fully illustrated and described global occurrences of Arumberia exhibit similar features (Table 1), which have variably been suggested to be resting cysts of Arumberia (Bland, Reference Bland1984), structures which mark a distinct stage in the development of Arumberia (Becker-Kerber et al. Reference Becker-Kerber, Paim, Junior, Girelli, da Rosa, El Albani, Osés, Prado, Figueiredo, Simões and Pacheco2020), independent body fossils assigned to the taxon ‘Beltanelliformis minutae’ (McIlroy et al. Reference McIlroy, Crimes and Pauley2005), pseudofossils associated with the perforation of microbial matgrounds (Menon et al. Reference Menon, McIlroy, Liu and Brasier2016) or volcanic lapilli (McIlroy & Walter, Reference McIlroy and Walter1997). While the features remain enigmatic, the local palaeogeographic context of the Port Lazo specimens eliminates the possibility of volcanic lapilli.

Fig. 7. ‘Dimple-pimple’ marks associated with Arumberia in the Port Lazo Formation. (a) Population of negative epirelief dimples on a plane directly underlying clay laminae hosting Arumberia lines. (b) Positive epirelief Arumberia lines superimposed by negative epirelief dimples. (c) Subcircular negative epirelief dimples occurring independently of Arumberia. Coin is 23.25 mm in diameter. (d) Positive epirelief pimples immediately overlain by negative epirelief dimples. (e) Interpretative sketch of 3D spheroidal orbs, and regularly viewed two-dimensional dimples and pimples.
4. Emended diagnostic criteria for identifying Arumberia
The original diagnosis of Arumberia provided by Glaessner & Walter (Reference Glaessner and Walter1975) was based on specimens from the Arumbera Sandstone, but 39 additional Arumberia reports from around the globe (Tables 1, 2), in addition to key characteristics recognized in the Port Lazo examples, necessitate modification of its diagnosis.

Fig. 8. Chronostratigraphic ranges of reported instances of Arumberia. Green bars mark Arumberia identifications that are fully described and photographed and shown to closely match either the original (Glaessner & Walter, Reference Glaessner and Walter1975) or emended (this study) Arumberia diagnosis. Orange bars mark instances where Arumberia has been photographed, but no other sedimentological or morphological details have been provided. Grey bars denote formations in which Arumberia have been stated to occur, but no informative details of morphology have been provided. Red bars indicate formations where Arumberia has been described but, following scrutiny of form (Section 5 ), likely represent features not equivalent to Arumberia (as originally diagnosed by Glaessner & Walter, Reference Glaessner and Walter1975). Yellow and purple columns indicate the chronostratigraphic restriction of Arumberia when dubious reports are excluded (680–520 Ma). Purple column alone shows a tighter stratigraphic range (560–520 Ma), evident when considering only overlapping ranges of posited ages.
Arumberia: Glaessner & Walter Reference Glaessner and Walter1975 (figs 6a–c, 7a–c, 8).
Emended diagnosis: Surface texture comprising parallel, sub-parallel or radiating lines 0.4–4.0 mm wide and spaced 0.25–30.0 mm apart. Lines can present in both positive and negative epirelief and hyporelief, with total line relief rarely exceeding 2.0 mm. Lines are commonly several centimetres long but may continue for tens of centimetres. Lines may bifurcate, but typically maintain constant width along their lateral extent. Edges are indistinct. Lines rarely overlap, but can transition laterally into reticulated networks.
Distribution: Arumberia has been described from every continent except Antarctica (see full list of localities in Table 2). Its temporal range is discussed in Section 5.
Discussion: Emended diagnoses for Arumberia and Arumberia banksi were recently presented by Retallack & Broz (Reference Retallack and Broz2020), following re-examination of the type material of Arumberia banksi and other specimens from Central Australia. They interpreted features including septae, chambers and radiating ribs within their studied material, and included these within their emended diagnosis. We find no evidence of septae or chambers in any of the Arumberia specimens we have studied, and also find no reason to consider the radiating pattern of ribs to be a diagnostic feature. As such, we do not include these features in our emended diagnosis.
There has been disagreement regarding whether Arumberia should be considered a genus (Glaessner & Walter, Reference Glaessner and Walter1975), arising from the uncertainty regarding its origin (see Section 6) and the absence of sufficient fossil material to permit more than a subjective hierarchical context. Recent formalization within the class Vendobionta (Seilacher, Reference Seilacher1992; Retallack & Broz, Reference Retallack and Broz2020) is not followed here, since: (1) a discrete macroorganism interpretation for Arumberia is seemingly incompatible with its ability to express quite different morphological forms (e.g. linear, Fig. 3a–h; reticulated, Fig. 3i, j; curled, Fig. 5) dependent on the prevailing environmental conditions; and (2) we do not consider Vendobionta to be a valid clade (Dunn & Liu, Reference Dunn and Liu2019). McIlroy et al. (Reference McIlroy, Crimes and Pauley2005) opted to classify it as a pseudofossil under ‘microbially modified sedimentary structures’. While we agree that there is the potential for a microbial role in the formation of this surface structure, we argue that the 3D morphology of individual lines indicates that they preserve casts of fossil organisms, rather than modifications of sedimentary surfaces. Our interpretation of Arumberia as the remains of a distinct organism or population of organisms requires that it retains its Linnaean terminology.
Glaessner & Walter’s (Reference Glaessner and Walter1975) type material was given the species name Arumberia banksi. Five additional Arumberia species have since been proposed (A. vindhyanensis, ‘A. usvaensis’, ‘A. multykensis’, ‘A. beckeri’ and ‘A. ollii’: Kumar & Pandey, Reference Kumar and Pandey2008; Kolesnikov et al. Reference Kolesnikov, Grazhdankin and Maslov2012). Presently, only A. banksi and A. vindhyanensis are considered valid (Kolesnikov et al. Reference Kolesnikov, Danelian, Gommeaux, Maslov and Grazhdankin2017), and these are differentiated by the frequency of line bifurcation. Arumberia banksi was most recently re-described by Retallack & Broz (Reference Retallack and Broz2020), while the systematic description of Arumberia vindhyanensis can be found in Kumar & Pandey (Reference Kumar and Pandey2008). In the Port Lazo Formation, specimens that could be attributed to both of these species can be seen to grade into one another (compare Fig. 3a and Fig. 3g), suggesting that subdivision to species level may have limited biological meaning (see Section 6.d). Morphological differences between forms may instead stem from external hydrodynamic or environmental factors.
5. Global stratigraphic range of true Arumberia
Arumberia has a narrow stratigraphic range, spanning the Neoproterozoic to lowermost Cambrian strata (Table 2), a trend that becomes particularly pronounced when equivocal reports of the surface texture are omitted (Fig. 8). These equivocal reports include 14 records where Arumberia is reported but not figured or described in a paper (i.e. with no evidence presented to verify or falsify the claim), and the following six instances that are here rejected as being Arumberia because they are either markedly dissimilar to both the original (Glaessner & Walter, Reference Glaessner and Walter1975) and emended diagnoses (Section 4), or are insufficiently well preserved to confirm their identification as Arumberia.
-
(1) Bland (Reference Bland1984) proposed that a sedimentary surface texture illustrated by Miller (Reference Miller1975, fig. 2) from the Mesoproterozoic Auborus Formation of Namibia may be Arumberia. However, the original figure lacks the definition to confirm this, and Miller (Reference Miller1975) interpreted the texture as forming abiotically due to aeolian adhesion on a mudflat.
-
(2) Bland (Reference Bland1984) suggested that a specimen illustrated by Kaudern (Reference Kaudern1932, fig. 5) from the Neoproterozoic Visingsö Formation of Sweden may represent poorly preserved casts of Arumberia. Kaudern’s (Reference Kaudern1932) original figure shows only a small hand specimen with no sedimentological context, and the sample is therefore an unconvincing example of Arumberia given its poor preservation.
-
(3) Callow et al. (Reference Callow, Battison and Brasier2011, fig. 10) reported and figured surface textures described as Arumberia from the early Neoproterozoic Diabaig Formation of Scotland. However, these surface textures differ from both the original and revised diagnoses of Arumberia, with lines being sporadically spaced, changing width along their lateral extent and showing a wide disparity of orientations even across small bedding planes. The interpretation of these specific structures by Callow et al. (Reference Callow, Battison and Brasier2011) as forming due to the moulding of entwined microbial filaments appears robust, but they are dissimilar to classic Arumberia.
-
(4) Reported Arumberia from the Ediacaran Masirah Bay Formation of Oman were figured by Allen & Leather (Reference Allen and Leather2006, figs 14c, d) and Brasier et al. (Reference Brasier, Antcliffe, Callow, Allison and Bottjer2011, fig. 11b). Brasier et al. (Reference Brasier, Antcliffe, Callow, Allison and Bottjer2011) noted that the features extended for up to “hundreds of square kilometres” and suggested that they reflected the flow of bottom currents interacting with possible filamentous microbes or algae. In both publications the small dimensions of the figured specimens make an assured diagnosis difficult, but their apparent and regular intertwining of sinuous ridges is not present in any other known Arumberia specimens (Table 1).
-
(5) Arumberia reported from the Upper Cambrian Booley Bay Formation of Ireland (MacGabhann et al. Reference MacGabhann, Murray and Nicholas2007, fig. 15) do not have a uniform line thickness across each bedding plane, unlike Arumberia examples described elsewhere (Table 1). Additionally, many individual lines exceed 0.5 cm in diameter, greater than any other previously described Arumberia.
-
(6) Liu (Reference Liu1981) described Arumberia from a limestone bed of the Shichang Member in China, the only report from a carbonate lithology. The small figured specimen makes confirmation difficult, but the positive epirelief lines appear to be an order of magnitude greater in size than any other Arumberia reports (Table 1).
When these examples are excluded, all remaining reports of Arumberia-hosting strata date to 520–680 Ma in age (Fig. 8). When considering the overlapping ranges of posited ages, the stratigraphic range can be further refined to 520–560 Ma in age (this is also true for 12 of the 14 instances of reported Arumberia that were not figured in their original reports).
6. Abiotic or biotic?
The limited range of consistently expressed morphological characteristics of ‘true’ Arumberia, and the inclusion of a wide variety of instances of clustered lines on bedding planes within the genus in the past, have resulted in ongoing debate regarding the biogenicity of this feature. Three lines of evidence are discussed here to interrogate these competing claims: similarity to abiotic forms, the tight global stratigraphic range and the preferred orientation of specimens.
6.a. Similarity to abiotic forms
Features with a passing resemblance to some Arumberia specimens were created abiotically by Dżułyński & Walton (Reference Dżułyński and Walton1965) in flume tank experiments devised to study sedimentary features associated with deep marine flysches. Brasier (Reference Brasier and House1979) was the first to note this similarity, and it has subsequently been invoked to contend that Arumberia was, at least in part, hydrodynamic in origin (e.g. Kumar & Pandey, Reference Kumar and Pandey2008; Callow et al. Reference Callow, Battison and Brasier2011; Menon et al. Reference Menon, McIlroy, Liu and Brasier2016). We note that the structures developed by Dżułyński & Walton (Reference Dżułyński and Walton1965) occurred within turbidity-current-generated sand, while the majority of Arumberia worldwide come from mudstones and siltstones deposited in emergent and nearshore marine environments (Table 1; a notable exception is the Masirah Bay Formation ‘Arumberia’ from lower shoreface/offshore facies (Allen & Leather, Reference Allen and Leather2006) discussed in Section 5). Tangible characteristics of exposed Arumberia in the Port Lazo Formation (3D cord-like form, reticulated transitions, potential for cords to curl when subject to emergence) cannot, in concert, be explained by purely hydrodynamic mechanisms. Cohesive mud and granular sand have very different responses to hydrodynamic forces, and so the similarity to Dżułyński & Walton’s (Reference Dżułyński and Walton1965) sedimentary structures is most likely an artefact of equifinality. Furthermore, the observation that exceptionally well preserved specimens comprise finer linear structures (Fig. 1d, h) rules out abiotic formation mechanisms that produce only surface impressions. The bundling of these cords is consistent with them preserving discrete filamentous organisms such as bacteria or algae, and is inexplicable by fluid dynamics alone.
6.b. Stratigraphic range
McIlroy & Walter (Reference McIlroy and Walter1997) used the tight stratigraphic range of Arumberia to suggest that it is an anactualistic fabric on sedimentary surfaces resulting from the interplay of microbial mats with physical hydrodynamic processes. However, this interpretation cannot explain the absence of Arumberia from Phanerozoic or lower Precambrian strata. Both of these intervals host abundant sedimentary surface textures attributable to the interplay of fluids and microbial mats (Davies et al. Reference Davies, Liu, Gibling and Miller2016). Arumberia is absent from comparable Phanerozoic sedimentary facies where bedding planes have received intensive study: for example, there are no reports of Arumberia in voluminous desiccated red mudrocks of the Siluro-Devonian Old Red Sandstone and equivalent strata, despite detailed investigations of surficial trace fossils (e.g. Gordon, Reference Gordon1988; Smith et al. Reference Smith, Braddy, Marriott and Briggs2003; Shillito & Davies, Reference Shillito and Davies2017) and microbial sedimentary surface textures (Davies et al. Reference Davies, Sansom and Turner2006, Reference Davies, Liu, Gibling and Miller2016; Marriott et al. Reference Marriott, Hillier and Morrissey2013) in littoral facies. Similarly, Arumberia is also unknown from pre-Ediacaran successions that have been interrogated for microbial sedimentary structures (e.g. Eriksson et al. Reference Eriksson, Bartman, Catuneanu, Mazumder and Lenhardt2012; Lan et al. Reference Lan, Chen, Li and Kaiho2013; Beraldi-Campesi et al. Reference Beraldi-Campesi, Farmer and Garcia-Pichel2014).
Given that no physical hydrodynamic fluid processes are likely to have been unique to the Ediacaran–Cambrian transition, the most parsimonious explanation for the stratigraphic restriction of Arumberia (Fig. 8) is that there was a dominant biological component to its formation (Bland, Reference Bland1984; Kumar & Pandey, Reference Kumar and Pandey2008, Reference Kumar and Pandey2009; Kolesnikov et al. Reference Kolesnikov, Grazhdankin and Maslov2012, Reference Kolesnikov, Marusin, Nagovitsin, Maslov and Grazhdankin2015, Reference Kolesnikov, Danelian, Gommeaux, Maslov and Grazhdankin2017; Arrouy et al. Reference Arrouy, Warren, Quaglio, Poiré, Simões, Rosa and Peral2016). This is also implied by the variability in individual Arumberia linearity (Fig. 5), reticulation (Fig. 3i, j) and spacing (Fig. 3a–h) observed within the large bedding plane studied at Port Lazo. Such morphological diversity is challenging to account for solely through hydrodynamic processes, but can be explained as different manifestations of a living system.
Forms resembling Arumberia have been observed on the surface of biofilms in modern salterns, with radiating and curved macroscopic ridges interpreted to form as a microbial community responds to changing environmental pressures (Kolesnikov et al. Reference Kolesnikov, Danelian, Gommeaux, Maslov and Grazhdankin2017). While these extant ‘arumberiamorph structures’ demonstrate a further mechanism by which equifinal Arumberia-like morphologies can form, they are unlikely to be direct analogues for the ancient Arumberia described here, given the global abundance of such forms in Ediacaran–Cambrian strata and the total absence of verified instances in the post-Cambrian rock record. The absence of Arumberia in any Phanerozoic strata, despite directed searching of equivalent facies, is most prosaically explained as a faithful historic testimony of its disappearance. This is because a Cambrian to Recent gap in the record of any sedimentary or fossil feature is significant: the notion that it is explainable by a > 500 Ma duration failure of the sedimentary-stratigraphic record to archive a relatively mundane sedimentary environment is unsubstantiated.
6.c. Orientation of Arumberia lines
One consensus viewpoint regarding Arumberia is that there is a dominant directionality to its orientation on individual surfaces, which has been considered to result from either physical (e.g. Brasier, Reference Brasier and House1979; Jenkins et al. Reference Jenkins, Plummer and Moriarty1981; McIlroy & Walter, Reference McIlroy and Walter1997; Seilacher, Reference Seilacher2007) or biological (Glaessner & Walter, Reference Glaessner and Walter1975; Bland, Reference Bland1984) mechanisms of alignment. Of the 40 reports in Table 1, 17 consider Arumberia line orientations with respect to the local current (as measured by ripple mark strike-lines on bedding planes hosting Arumberia, or indirectly inferred from palaeoflow indicators measured throughout the host succession). These case studies reveal that Arumberia may in fact strike both parallel and perpendicular to the original flow direction (Fig. 9a–f; Table 1). Within the Port Lazo Formation studied here, a strong perpendicular arrangement is apparent (Section 3.b; Fig. 6).

Fig. 9. Relationships between Arumberia lines and original flow. (a) Flow-perpendicular Arumberia superimposed on top of symmetrical wave-ripple marks. Port Lazo Formation Upper Member. (b, c) Flow-parallel Arumberia situated on top of unidirectional linguoid ripple marks. Crown Hill Formation, Newfoundland. (d) Flow-parallel to oblique Arumberia situated above symmetrical ripple marks. Ferryland Head Formation, Newfoundland. (e) Flow-parallel Arumberia situated above asymmetrical ripples marks. Crown Hill Formation, Newfoundland. (f) Flow-perpendicular Arumberia situated above poorly developed wave-ripple marks. Gibbett Hill Formation, Newfoundland. Hand lens is 4 cm wide. Coin is 16.25 mm in diameter.
Kolesnikov et al. (Reference Kolesnikov, Danelian, Gommeaux, Maslov and Grazhdankin2017) suggested that the organization of Arumberia lines may not necessarily be current-induced, but instead archive a trophic response of a microbial community in competition for sunlight. However, if this were the case, line orientation should be expected to be random relative to flow, rather than preferentially oriented with or normal to the prevailing current direction. We note that in several cases, Arumberia is aligned with flow in those environments where there was a unidirectional component (as shown by preserved linguoid ripple marks with superimposed, parallel Arumberia lines; Fig. 9b, c), but lies perpendicular to flow in settings with dominantly oscillatory flow conditions (as shown by Arumberia lines striking alongside preserved symmetrical wave-ripple marks; Fig. 9a, f). This would imply that Arumberia orientation could be in flux when ‘active’, and that the orientation of preserved Arumberia lines was dependent on external influences at the chance instant of burial. Lines aligned with ripple crests under gently oscillating flow, but were streamlined in the direction of a steady flow (e.g. see Davies et al. Reference Davies, Shillito and McMahon2017). Reorganization in this way suggests that the Arumberia lines may record flexible cords that were pliant with hydrodynamic forcing. Such organization has analogue with extant marine algae, the flexible components of which reorganize and re-orientate under increasing unidirectionality and discharge of water flow (e.g. Denny & Gaylord, Reference Denny and Gaylord2002).
Perpendicular arrangements with the prevailing current flow, observed in the Port Lazo Formation (Fig. 6) and elsewhere (Table 1), require that there was a secondary, weaker current that was strong enough to re-orientate Arumberia but insufficient to reorganize the sediment grains on the colonized surface. One potential scenario arises when a subordinate current drains water away following an interval of elevated water level (e.g. in the receding waters of a tidal prism). Such conditions were likely frequently met in the deposition of Arumberia-bearing strata, which are composed of periodically emergent facies in the majority of unequivocal reports of the form (Table 1). In the Port Lazo Formation, evidence of water drainage and exposure is indicated by the occurrence of raindrop impressions (Fig. 2g) and desiccation cracks (Fig. 2f) alongside Arumberia, and possibly also through the reticulated Arumberia forms (Fig. 3i, j).
6.d. Arumberia is most likely a fossil organism
The arguments above strongly suggest that Arumberia records an extinct, environmentally restricted and sessile biological entity that was composed of non-biomineralized, 3D, flexible cords (see also Bland, Reference Bland1984, p. 630). The origins of some of its associated features (e.g. dimple-pimple marks) remain enigmatic, and its precise biological affinity remains uncertain, even though it is parsimonious to assume that it was algal or microbial. While this conclusion is similar to that reached by Kolesnikov et al. (Reference Kolesnikov, Grazhdankin and Maslov2012), who suggested that Arumberia records extinct, highly organized microbial colonies capable of producing 3D ‘rugae’, we caution against splitting the texture into biological ‘species’ given the propensity of these to grade into one another (Section 4). Differences between these forms (e.g. A. banksi, A. vindhyanensis and also non-Arumberia linear morphologies) may stem from external hydrodynamic or environmental factors, reflecting the form that happened to be adopted at the instant of burial. Clear transitions between linear (Fig. 3a–h), reticulated (Fig. 3i, j) and curled (Fig. 5) Arumberia rule out a recent ‘vendobiont’ fossil hypothesis (Retallack & Broz, Reference Retallack and Broz2020), since discrete macroorganisms would not have been expected to reorganize their form during emergence.
7. Conclusions
Arumberia consists of parallel, sub-parallel or radiating lines that present in both positive and negative epirelief. A detailed examination of the most contiguous Arumberia-bearing bedding plane so far discovered, from the Port Lazo Formation (NW France), combined with a literature survey of all known Arumberia reports, indicates:
-
the 3D morphology of Arumberia resemble cords or tubes distinct from the preserving sediment;
-
linear Arumberia cords transition laterally into reticulated networks, morphologies that bear strong resemblance to modern instances of reticulate marks formed as buoyant filaments of bacteria or algae become tangled in standing or draining water;
-
some Arumberia cords are demonstrably composed of finer, superimposed and overlapping threads;
-
Arumberia cords curl in proximity to desiccated plate margins, an apparent response to shrinking cohesive sediment undergoing emergence;
-
the orientation of Arumberia lines differs in different depositional settings, with lines apparently being streamlined in the direction of a unidirectional current, but oriented parallel to ripple crests when subject to oscillating flow; and
-
the significant majority of Arumberia occurrences worldwide are recorded from rocks dated to 520–560 Ma in age.
Together, these observations discredit the possibility that Arumberia is a purely physical sedimentary structure, and strongly favour a biological origin as the impression of an extinct, sessile Ediacaran–Cambrian organism or population of organisms composed of 3D, flexible cords.
Acknowledgements
Fieldwork in France was supported by Shell International Exploration and Production B.V under Research Framework agreement PT38181. Fieldwork in Canada was partially supported by Natural Environment Research Council grant NE/P002412/1, awarded to Dr Emily Mitchell, University of Cambridge. We thank Professor Sören Jensen and one anonymous reviewer for their constructive reviews of this paper, and editor Dr Bas Van de Schootbrugge for handling this paper.













 Open access
Open access