



Introduction
Old man’s beard (also called traveller’s joy; Clematis vitalba L.) is a liana species native to warm temperate regions in western, central, and southern Europe, northern Africa, and western Asia (Tutin et al. Reference Tutin, Heywood, Burges, Valentine, Walters and Webb1964; USDA-ARS 2003), but has been introduced as an ornamental to many parts of the world, and is now found as a naturalized species in northern and eastern Europe (Danielewicz and Wiatrowska Reference Danielewicz and Wiatrowska2014; Fitter Reference Fitter1978; Möllerová Reference Möllerová2005), North America (USDA-NRCS 2006), Australia (Biosecurity Queensland 2016), and New Zealand (Allan Reference Allan1940). Twining leaf petioles allow C. vitalba stems to rapidly climb nearby vegetation like a trellis and blanket their hosts in the upper canopy, depriving them of sunlight, thus reducing photosynthesis and eventually causing tree collapse in some cases (Atkinson Reference Atkinson1984; Gourlay et al. Reference Gourlay, Wittenberg, Hill, Spiers and Fowler1999; Ogle et al. Reference Ogle, Cock, Arnold and Mickleson2000). Despite extensive ongoing efforts to control it, C. vitalba remains one of the most problematic invasive plant species in New Zealand and is recognized as a threat to native plant diversity and abundance, as well as native forest structure (Environmental Protection Authority 2018; Ogle et al. Reference Ogle, Cock, Arnold and Mickleson2000; Redmond and Stout Reference Redmond and Stout2018). It is also becoming an increasing problem in production forestry and agricultural riparian plantings (Te Uru Rākau–New Zealand Forest Service 2024).
Most research on liana competition has studied relationships between lianas and trees or other woody plants. For example, previous research in mature secondary forest in Panama showed that seedlings of all liana species studied outperform tree seedlings in low-light situations (Pasquini et al. Reference Pasquini, Wright and Santiago2015). Also, the combined effect of underground and aboveground competition with the liana ngobop (Acacia kamerunensis Gand.) has been shown to reduce biomass of three tropical tree species by 42% (Toledo-Aceves and Swain Reference Toledo-Aceves and Swain2008).
Successful plant invasion is contingent on favorable interactions with the biotic and abiotic elements in the environment, which provide satisfactory levels of nutrients, water, and light (Gurevitch Reference Gurevitch2011; Wilson et al. Reference Wilson, Steel and Steel2007). Although it has shown tolerance of poor-quality soils in New Zealand, C. vitalba generally grows best in soils that are moderately to well drained, are not highly acidic, and do not have severe nutrient deficiencies (Hume et al. Reference Hume, West and Watts1995). The species also has a positive response to calcium, phosphorous, and nitrates (Bungard et al. Reference Bungard, Morton, McNeil and Daly1998; Hume et al. Reference Hume, West and Watts1995). Zhang et al. (Reference Zhang, Jiang, Wang, Zhang, Gai, Peng, Zhang, Mao, Lou, Mo, Feng and Ming2021) demonstrated that mature C. vitalba plants are resistant to heat shock, which may be advantageous during a particularly hot summer or when growing in open areas with direct sunlight. Shade tolerance of mature C. vitalba has been shown, yet higher irradiance levels stimulate rapid growth (Baars and Kelly Reference Baars and Kelly1996; Bungard et al. Reference Bungard, Morton, McNeil and Daly1998; Paliwal et al. Reference Paliwal, Kuppers and Schneider1994). Williams (Reference Williams2009, p. 12) reported that once mature, C. vitalba is “highly competitive with all associated vegetation.” Ogle et al. (Reference Ogle, Cock, Arnold and Mickleson2000) attributed the disappearance of 24% of previously recorded shrub and small tree species and 37% herbaceous plant species in Taihape, New Zealand, reserves over a 26-yr period to the increasing cover of C. vitalba. Yet no controlled studies looking at the effects of competition on C. vitalba have been published, at any stage of life, but especially at the seedling stage.
Clematis vitalba is known to produce up to 50,000 viable seeds m−2 yr−1 in heavily infested areas (Jarvis-Lowry Reference Jarvis-Lowry2023). However, the potential of those seeds to cause new infestations after dispersal has not been clear. Germination and the young seedling phase are particularly vulnerable periods in a plant’s life cycle (Batlla and Benech-Arnold Reference Batlla and Benech-Arnold2014; Silvertown et al. Reference Silvertown, Franco, Pisanty and Mendoza1993). West (Reference West1992) posited that C. vitalba seedlings in New Zealand have high mortality and are negatively density dependent. Also, in contrast to mature plants, C. vitalba seedlings in southwestern Russia have been reported to be intolerant of dry conditions (Kozlovskiy et al. Reference Kozlovskiy, Fedorinova, Kuropyatnikov and Dmitriev2017). When studying the effects of light and nitrogen levels on C. vitalba, Bungard et al. (Reference Bungard, Morton, McNeil and Daly1998) found that despite the wide tolerance of C. vitalba to low light levels, seedling transplants did not survive in a deeply shaded forest in New Zealand. However, another New Zealand study focused on light levels observed that, when grown in situ from seed, 20 times more C. vitalba seedlings emerged in open sites than in forested sites, yet seedling survival after emergence was proportionally higher in the forested sites (Nikoloff Reference Nikoloff2011). Thus, open habitats supported more seedlings, even though many did not survive in such settings (Nikoloff Reference Nikoloff2011).
As with many lianas, initial establishment of C. vitalba often occurs in forest gaps or edges, such as the open habitats noted previously. Clematis vitalba is also common in riparian zones. In such settings, competition for resources with grasses and other ground cover vegetation is a major factor in limiting establishment success. Established perennial grass swards have been shown to prevent the emergence of the woody shrub species Scotch broom [Cytisus scoparius (L.) Link] in New Zealand production forests (Tran et al. Reference Tran, Harrington, Ghanizadeh, Robertson and Watt2018). Also, perennial grasses were highly effective at limiting the establishment of woody plants in a sub-Mediterranean ecosystem in British Columbia, Canada (MacDougall and Turkington Reference MacDougall and Turkington2005). Seedling establishment in dense ground cover vegetation thus may be quite limited, but recent work has determined that C. vitalba plants can spread vegetatively and vigorously in dense networks by creeping along the ground (Jarvis-Lowry et al. Reference Jarvis-Lowry, Harrington, Ghanizadeh and Robertson2024a).
Local land managers in New Zealand have identified what appear to be young C. vitalba in grasses of riparian zones and in production forests (A Gordon, personal communication, October 7, 2019; J Keast, personal communication, April 17, 2019), but it is uncertain whether the shoots present in these areas are young seedlings or new stems produced by creeping vines that had established in previous years. Also, in forestry settings, C. vitalba appears to be less of a problem in grass swards than in the soil directly around tree seedlings kept bare by herbicides to allow the seedlings to grow competition-free.
Research summarized by Jarvis-Lowry (Reference Jarvis-Lowry2023) has shown that C. vitalba seeds can remain viable following extended immersion, and thus could survive a flooding event. Whether seeds introduced via flooding are likely to produce seedlings is unknown. In addition, germination of seeds in the presence of established grasses has not been studied, and no formal research has investigated the risk of invasion of grassy habitats by the species. The objective of this experiment was to assess the ability of C. vitalba seeds to germinate, emerge through the soil, and establish (survive 1 yr) within grassy swards, which are common in riparian zones or exotic forests.
Materials and Methods
Study Sites
A preliminary experiment was conducted from November 2019 to November 2020, and then was expanded and repeated, with two experiments begun in September 2020, one of which ran for 1 yr and the other for 2 yr (until September 2021 and 2022, respectively). The preliminary experiment, M1 (−40.387806, 175.612194; Site Mānuka 1), began in late spring on a portion of the Massey University Crop and Pasture Research Unit near Palmerston North, New Zealand. Plots were established on slopes of 25° to 30° in large 5 by 5 m grassy blocks with rows of young, 1.5 m-tall mānuka (Leptospermum scoparium J.R. Forst. & G. Forst.) bushes at the edges. The blocks had been fenced off to prevent grazing for 10 yr before the experiment commenced. Ground-level vegetation in the blocks was dense and vigorous. Apart from the L. scoparium, vegetative composition in these blocks consisted of approximately 50% Yorkshire fog (Holcus lanatus L.), 25% creeping buttercup (Ranunculus repens L.), 10% rough-stalked meadow grass (Poa trivialis L.), 5% white clover (Trifolium repens L.), and 5% perennial ryegrass (Lolium perenne L.), and the remaining 5% was an assortment of other grasses and broadleaf weeds.
In anticipation of higher C. vitalba emergence and survival with earlier sowing, the subsequent two experiments were begun 2 mo earlier the next season (September 2020) at two locations within the Crop and Pasture Research Unit: (1) M2 (−40.388, 175.612389; Site Mānuka 2), adjacent to the original site, with nearly identical properties; and (2) TL (−40.386694, 175.608806; Site Treeline), approximately 500 m west of M2, located inside the dripline beneath a row of 20- to 30-m-tall exotic Monterey pine (Pinus radiata D. Don) trees planted on the edge of the research unit as a windbreak. The Treeline site was chosen to contrast in two important ways with the two Mānuka sites: (1) it had been recently grazed and was fenced to exclude grazing only 2 wk before the commencement of the experiment; and (2) the large number of shallow roots and the P. radiata cover produced microclimatic conditions at the ground level that modulated temperature and limited light and moisture. As such, it was much drier and ground-level vegetation was sparser than at the other two sites. None of the sites had a previous history of C. vitalba presence. Ground-level vegetative composition in the TL plots consisted of roughly 40% L. perenne, 15% soft brome (Bromus hordeaceus L.), 15% daisy (Bellis perennis L.), 15% T. repens, and 10% common chickweed [Stellaria media (L.) Vill.], with the remaining 5% an assortment of other grasses and broadleaf weeds. Apart from the native L. scoparium, all species in the research sites were exotics, introduced either for a shelterbelt (P. radiata) or pasture purposes (grasses, T. repens), or were introduced as inadvertent weeds (all others; R. repens is especially troublesome in pastures).
Soil type at all three sites is a silt loam and, according to S-Map online (Landcare Research NZ Ltd 2019), is moderately deep and poorly drained. Soil sample analysis results of soil organic matter, pH, phosphorous, calcium, magnesium, potassium, sodium, cation exchange capacity, base saturation, and texture are given in the Supplementary Information (Supplementary Table S1), along with brief notes on minor site differences.
Vegetative Cover Treatments
Plots were arranged in a randomized complete block design with four replicates. The following vegetative cover treatments were applied in all three experiments: (1) following removal of existing vegetation, soil kept bare throughout the experiment with hand weeding (Bare); (2) vegetation cut to 4 cm, with no further trimming after sowing (Cut); and (3) uncut vegetation (Long). In addition, a fourth treatment was added to M2 and TL: bare soil initially, with no maintenance after sowing (IBare). Plots in all three trials measured 60 by 60 cm (marked with wooden pegs), with a 60-cm buffer between them, and a 1-m buffer from the rows of L. scoparium shrubs in M1 and M2, and a 6-m buffer from the trunks of the P. radiata at TL. Plots in each block were randomly assigned treatments, and plots for bare-soil treatments were sprayed with glyphosate (using 10 ml L−1 of Roundup 360 Pro (Bayer, 700 Chesterfield Parkway West, Chesterfield, MO 63017) [360 g ae L−1] applied to runoff, equivalent to at least 3.6 kg ae ha−1 of glyphosate) at 3 wk before sowing the seeds. All plant parts that remained after the herbicide treatment were removed in bare-soil plots 1 d before sowing. Also, vegetation in the cut-grass plots was trimmed to approximately 4 cm the day before sowing. Vegetation in buffers around the bare-soil and cut-grass plots was trimmed to approximately 30-cm height to ensure no border-effect shading occurred. On average, uncut vegetation in the Long plots was 60-cm tall at the time of sowing.
Seed Collection and Sowing Times
Achenes (one-seeded fruits, hereafter referred to as seeds) of C. vitalba were collected in the spring (September) before planting that same year from two different populations within 35 km of the trial site. Seeds that were flat, eaten, or smaller than 50% of the average size were discarded; the remaining seeds were stored dry at 5 C until the day of sowing. Seeds were sown into the prepared plots November 28, 2019, for M1 and approximately 2 mo earlier the following year (September 17, 2020) for M2 and TL. On the day of sowing, seeds from both populations were pooled. Average laboratory germinability of a sample of the pooled seeds collected in September 2019 and September 2020 was observed to be approximately 37% and 48%, respectively, in complete darkness (Jarvis-Lowry Reference Jarvis-Lowry2023). In each plot, 100 seeds were sown by hand, 5 mm under the soil surface in five lines 20-cm long and 5-cm apart (20 seeds per 20-cm line).
Data Collection
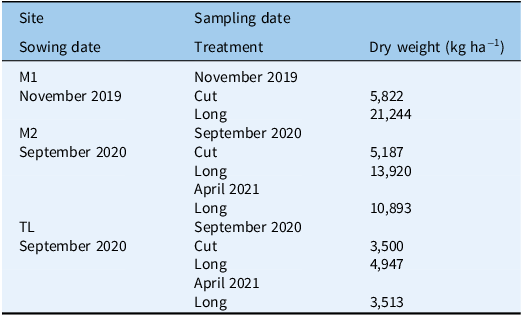
Vegetation samples were cut from the buffers outside the sown plots, using a 50 by 50 cm quadrat to estimate percent dry matter composition at the time of sowing: 18 samples at M1 and 12 each at M2 and TL (Table 1). Half of the biomass samples were taken to represent the initial vegetative cover level present in the Cut plots, which were trimmed to 4 cm before sampling. The other half were samples of uncut vegetation, representing conditions present in the Long plots. In Experiment M2, to get an indication of how the dry weight might have changed between spring 2020 and autumn 2021, six additional 50 by 50 cm quadrat samples were taken in the buffers in April 2021.
Table 1. Average dry weight of vegetation in kg ha−1 in plots cut to 4 cm (Cut) and uncut plots (Long) at three sites in Palmerston North, New Zealand: Mānuka 1, Mānuka 2, and Treeline (M1, M2, and TL, respectively). Samples were taken immediately before sowing in late-spring or early-spring (November 2019 or September 2020, respectively), and in mid-autumn (April 2021) following the September 2020 sowing date.
Plots were monitored regularly for emergence of C. vitalba seedlings, and Bare plots were kept bare with hand weeding. Number, size, and condition of seedlings present in each plot were recorded at 10, 15, 25, and 52 wk after sowing, with the last two data collections approximately 6 and 12 mo after sowing. Seedlings present in the 12-mo counts survived winter and were considered to have successfully established. At TL, new C. vitalba seedlings began emerging in some plots after 12 mo. Although the plots were not weeded after 12 mo, monitoring was extended at TL for another year into 2022 (at 61 and 104 wk after sowing) to assess the survivability of seedlings that establish the year after sowing.
Weather data from the AgResearch weather station 1 km away were recorded during the 2-yr experimentation period (Supplementary Figure S1). The maximum average daily temperature ranged from 24.6 C in February to 14.5 C in July. No supplemental watering was provided to the plots.
The electrified fencing surrounding the plots at M2 was inadvertently left off for a short period following sowing in September of 2020, and lamb (Ovis aries) hoofprints were found in the fourth block of the Bare and Cut treatments during the first monitoring/weeding session. Thereafter, no seedlings emerged in those plots, so no data could be obtained from them and Block 4 Bare and Cut plots were dropped from the analysis.
Data Analysis
All experimental analyses were performed using R statistical software v. 4.2.0 (R Core Team 2021). Seedling survival at each site was analyzed independently, with eight different generalized linear mixed-effects regression models for counts using the lme4 package (Bates et al. Reference Bates, Maechler, Bolker and Walker2015). Block was included as a random factor, with treatment as a fixed factor. For each of the three sites, one model was fit to the data collected 10 wk after sowing, and a separate model was fit to the data at the last monitoring event, 52 wk after sowing, at which time seedlings were considered to have survived to establish a perennial plant. For TL, an additional two models were fitted to data collected 61 and 104 wk after sowing (10 and 52 wk into the second year, respectively) to estimate treatment effects on seedling establishment in 2021 and subsequent seedling survival into 2022. Due to overdispersion, a negative binomial analysis was used for data from TL at 10 and 61 wk after sowing, but a Poisson was used for all other models. However, low seedling emergence and even lower survivorship made robust model estimates of block or treatment effects difficult to obtain. For example, because no seedlings ever established in the unmanipulated Long treatments at either M1 or M2, only the Bare and Cut treatments were used in those analyses. Also, all zeros in the IBare and Cut treatment data at M2 resulted in an uninformative dataset and a singular Hessian matrix for the Week 52 model. Failure of model convergence for Week 52 at TL was resolved by removing the all-zeros Long treatment data. Goodness of fit for each model was tested by plotting simulated residuals with the DHARMa package (Hartig Reference Hartig2022). Post hoc multiple pairwise comparisons were made using the multcomp package (Hothorn et al. Reference Hothorn, Bretz and Westfall2008).
Results and Discussion
Effect of Sowing Time and Seasonal Conditions
Higher invasion potential has been attributed to plants whose seeds germinate and seedlings emerge earlier in the growing season, because early access to finite resources can enhance their prospects for reaching maturity (Bakker et al. Reference Bakker, Dekker and De Vries1980; Guido et al. Reference Guido, Hoss and Pillar2017). In this research, time of sowing may have been an important factor in determining C. vitalba seedling emergence. Jarvis-Lowry (Reference Jarvis-Lowry2023) noted that under optimal laboratory circumstances, approximately 80% of C. vitalba seeds collected and incubated on soilless blotting paper in early spring will germinate when exposed to light, but only about 40% of seeds will germinate without light. In our study, fewer than 10% of seeds sown in late spring (M1, November 2019) in plots kept bare by weeding (Bare) produced a seedling (Figure 1A). In contrast, 36% of seeds sown in Bare plots earlier in the spring in 2020 (M2, September 2020) produced a seedling (Figure 1B), slightly lower than the reference laboratory germination results for seeds kept in darkness. As noted previously, one Bare M2 plot was trampled by sheep and did not support any seedling establishment, so the M2 results were averaged across only three of the four blocks for the Bare treatment. Lower emergence in the field compared with the high germination in the lab could be explained by more factors being involved in the successful germination of seeds and subsequent seedling emergence in a natural setting than in the laboratory, most importantly those having to do with soil, such as physical barriers to emergence through the soil, uncontrolled temperature and moisture, restricted nutrient and gas exchange (Brady and Weil Reference Brady and Weil1999), and the presence of soil fauna (Gioria and Pyšek Reference Gioria and Pyšek2017; Gurevitch Reference Gurevitch2011; Van Mourik et al. Reference Van Mourik, Stomph and Murdoch2005).
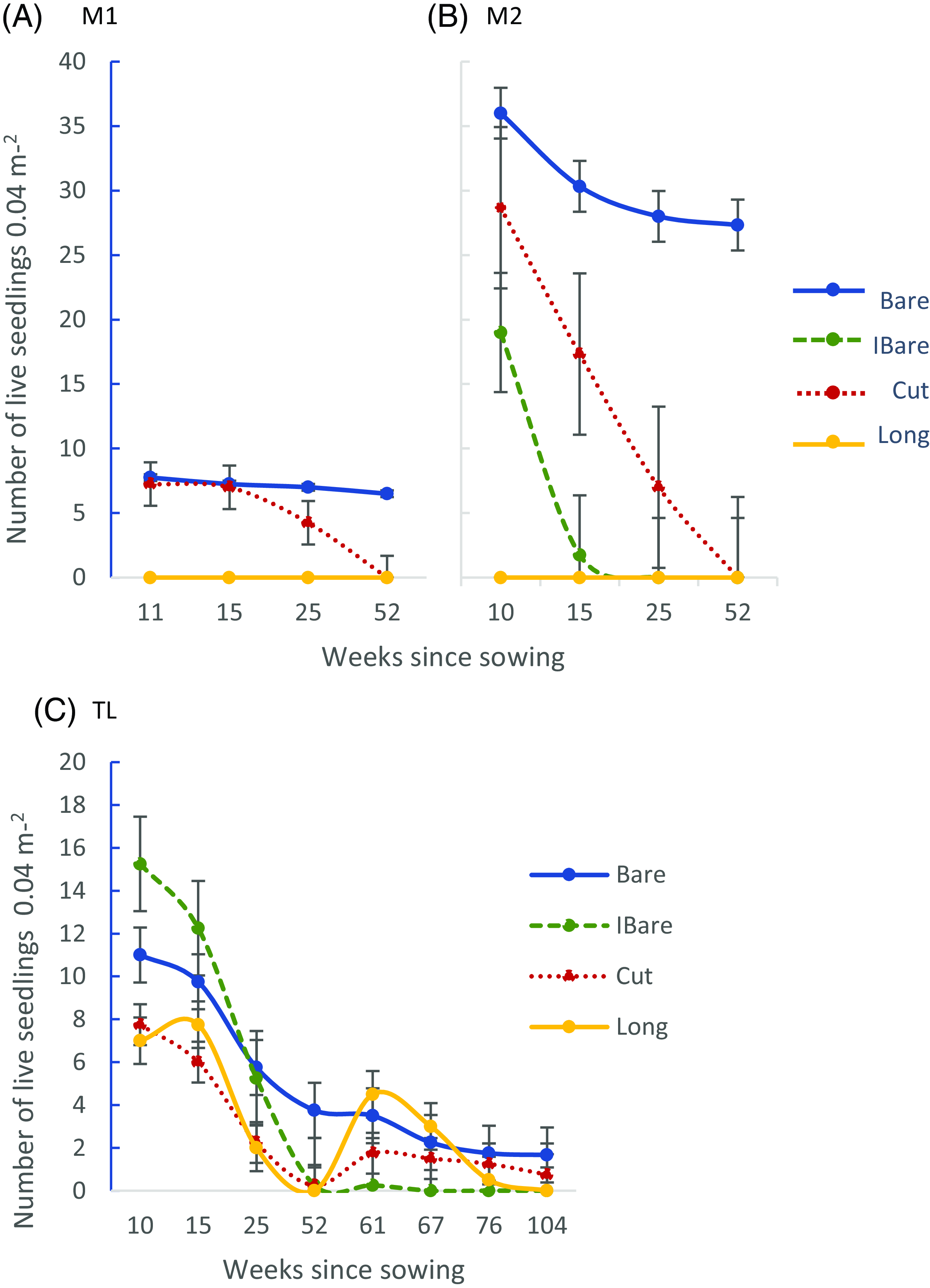
Figure 1. Average Clematis vitalba seedling emergence and survival (raw data) per plot after sowing (100 seeds plot−1): over a 1-yr period at two densely vegetated sites interspersed with Leptospermum scoparium plants (A) from November 2019 to November 2020 (late spring to late spring at the Mānuka 1 site [M1]) and (B) from September 2020 to September 2021 (early spring to early spring at Mānuka 2 site [M2]); and (C) over a 2-yr period (September 2020–September 2022, early spring to early spring) after sowing at a sparsely vegetated site beneath a row of Pinus radiata trees (Treeline, TL). Error bars based on standard errors of the arithmetic mean. Bare, plots maintained bare (bare at the time of sowing, kept weeded and trimmed throughout the experiment); IBare, plots initially bare (bare at the time of sowing, but not weeded or trimmed thereafter); Cut, vegetation in plots cut to ∼ 4 cm at the time of sowing, but not trimmed again; Long, seeds sown in plots with unmanipulated vegetation. M1 did not include an IBare treatment.
Apart from the advantage of an early start, the September 2020 sowing benefited from more rainfall in the first 4 mo of the early-establishment phase than did those of the November 2019 sowings. Hotter late-spring temperatures by November may also have been less hospitable than September temperatures for emerging seedlings. Furthermore, differences between seed cohorts could have had a minor effect on the differences in outcome between M1 and M2 (Jarvis-Lowry Reference Jarvis-Lowry2023).
Effect of Vegetative Cover Treatments
Because of poor emergence, Poisson models showed very little variation among blocks and no vegetative cover treatment effect on C. vitalba seedling establishment in M1, at either Week 10 or 52 (Supplementary Table S2). At M2, where more C. vitalba seedlings emerged than at the other two sites, initial seedling establishment (Week 10) differed significantly among all treatments (P < 0.001; Supplementary Table S3), with Bare plots having the highest emergence, followed by Cut plots, then IBare plots and Long plots, with an average of 36%, 29%, 19%, and 0% of C. vitalba seedlings emerging, respectively (Figure 1B). By Week 52, no seedlings were still alive in any treatment other than the Bare treatment, where an average of 27% survived.
At the M1 and M2 sites, plots experienced some shading by L. scoparium plants, as some plots were within 2 m of the L. scoparium. However, crowding by the vegetation within plots likely played a greater role in restricting C. vitalba seedling access to limiting resources (light, water, nutrients). Previous research has shown that availability of light, soil moisture, and soil nitrogen to both shrubs (silverberry [Elaeagnus commutata Bernh. ex Rydb.]) and tussock grass [Bouteloua gracilis (Willd. ex Kunth) Lag. ex Griffiths] grown without other vegetation present is two to five times greater than for plants grown in the presence of other plants and that access to light declines linearly as neighboring plant biomass increases (Peltzer and Köchy Reference Peltzer and Köchy2001). In this study, of those C. vitalba seedlings at M1 and M2 that emerged in the treatments without other vegetation present (Bare plots), within 6 mo almost all seedlings had produced vigorous, multiple stems, each individual stem averaging more than 30 cm long (Figure 2A and 2B). More than 75% of seedlings in the Bare treatment at the Mānuka sites persisted to Week 52 (Figure 1A and 1B). By comparison, no recognizable seedlings were ever found in the treatment with dense, grassy vegetation (Long plots) at either of the two sites. Additionally, C. vitalba seedlings that emerged in treatments with managed vegetation levels (Cut and IBare) remained single-stemmed and small, with numbers that declined as regrowth of the grass and other weeds progressed. No seedlings in either Mānuka site survived the winter in plots with any vegetation present other than C. vitalba.
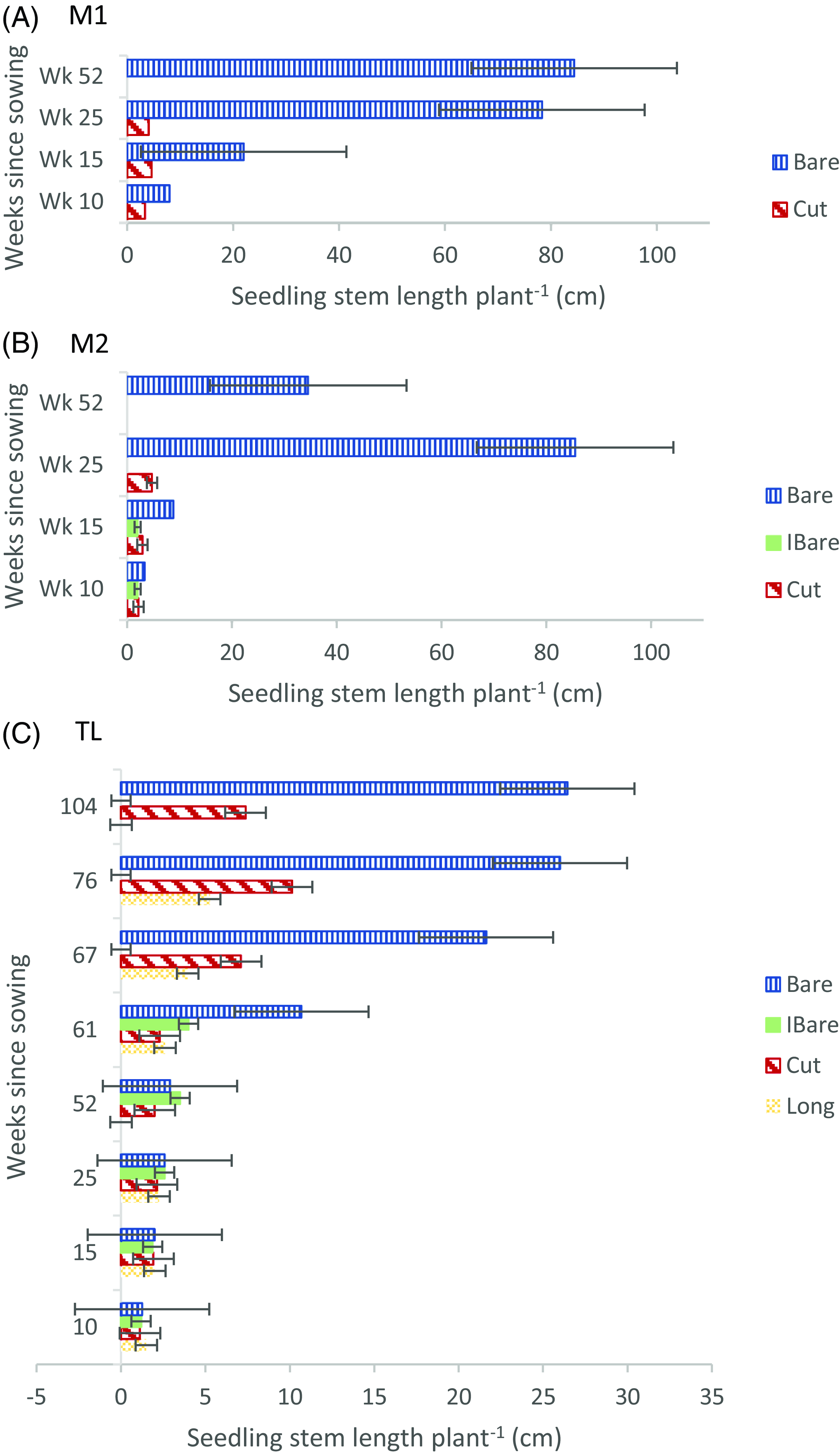
Figure 2. Average stem length of Clematis vitalba seedlings after sowing (A) in three levels of ground cover density at the Mānuka 1 site in late spring (M1) and four levels of ground cover density (B) at the Mānuka 2 site in early spring (M2), and (C) at the Treeline site (TL). At M1 and M2, no seedlings emerged in an additional treatment with unmanipulated vegetation, which is thus not shown. Seedlings in IBare and Cut treatments remained single-stemmed; Bare seedlings were multi-stemmed by 15 wk (WK 15). At TL, all seedlings between Wk (WK) 10-61 remained single-stemmed. At Week 67 (WK 67), seedlings in treatments Bare and Cut were multi-stemmed. Average seedling height multiplied by average number of stems to reflect average total stem length per plant. Bars represent standard error. Bare, plots maintained bare (bare at the time of sowing, kept weeded and trimmed throughout the experiment); IBare, plots initially bare (bare at the time of sowing, but not weeded or trimmed thereafter); Cut, vegetation in plots cut to ∼ 4 cm at the time of sowing, but not trimmed again; Long, seeds sown in plots with unmanipulated vegetation. M1 did not include an IBare treatment.
Each new stem of surviving C. vitalba plants represents a potential searching and/or climbing vine. In addition, according to Buru et al. (Reference Buru, Dhileepan, Osunkoya and Firn2016), development of multiple stems allows vines to maximize photosynthesis and improve competitive ability. Tanentzap et al. (Reference Tanentzap, Mountford, Cooke and Coomes2012) have also reported that forest understory trees with multiple stems have a greater chance of survival, due to the additive growth of each stem and the buffering effect of allocating resources among several stems in case of individual stem loss. Thus, reaching the stage where ancillary stems are produced may be critical for C. vitalba seedling survival to maturity, especially in densely vegetated habitats in combination with other stresses. In fact, only the smallest, single-stemmed seedlings in the Bare plots at the Mānuka sites died over the winter (7% at M1 and 2% at M2). Despite complete winter senescence of the smallest Bare seedlings and dieback of some stems on most seedlings, all Bare seedlings that did persist through winter had developed vigorous root systems when removed after Week 52 (Figure 3). These appeared to be well positioned to continue into maturity.

Figure 3. Vigorous root system of 1-yr-old Clematis vitalba seedling at Mānuka 2 (M2) site, grown in a plot kept bare by weeding for the first 6 mo of life.
The purpose of adding the IBare treatment at M2 and TL was to simulate what might happen in a situation where no intervention occurs after initially clearance of the land. It was also intended to be an intermediate level of vegetation density between the Bare and Cut treatments. At TL, the IBare plots supported significantly more seedlings than Cut plots at first (Figure 1C). However, at M2, not only did Cut plots support significantly more seedlings than IBare plots, but seedling numbers in the IBare plots also declined much more rapidly than those in the Cut plots (90% IBare mortality as opposed to 30% Cut mortality between Weeks 10 and 15; Figure 1B), suggesting that after the temporary bare phase, resources were more limiting in the IBare plots. More diverse types of weeds and more large-leaved weeds (curly dock [Rumex obtusifolius L.], bull thistle [Cirsium vulgare (Savi) Ten.], common sowthistle [Sonchus oleraceus L.], tall fleabane [Conyza sumatrensis (Retz. E. Walker)]) were found to have populated M2’s IBare plots after the first few weeks, whereas, aside from the height of the plants, the vegetative makeup of the Cut plots did not change over time. It may be that the difference between the two treatments was a result of the large broadleaf weeds that were able to establish in the IBare plots reducing the amount of light available to the C. vitalba seedlings in the plots. Regardless of the abundance of C. vitalba seedlings present within the first few months after sowing, no seedlings progressed past the single-stem stage, nor did any survive the winter in either the Cut or IBare treatment at M2. These results indicate that the removal of potentially competing species allowed the target species to develop vigorously, likely due to a greater availability of light and soil resources. Conversely, increasing vegetation density and consequent limitation of available light during the vulnerable young seedling stage prevented C. vitalba from surviving.
It is worth mentioning here that the Cut treatment was not intended as a simulation of a grazed pasture. Livestock readily graze C. vitalba. Therefore, seedlings that emerge in a consistently grazed pasture would be very unlikely to establish and reach maturity.
Site Effect
In the TL experiment, although this was not the only treatment with seedlings that survived to Week 52, the largest proportion of seedlings persisted in plots kept bare by weeding (Bare), and significantly more seedlings were still alive at the end of the study in the Bare treatment (P < 0.05) than in any other treatments. Only a small fraction of seedlings in the IBare and Cut treatments survived the winter, with no seedlings in the Long plots surviving. Initial emergence of seedlings in every treatment at TL, including the Long treatment, suggests that the lower ground cover density at this site than at either M1 or M2 was advantageous to C. vitalba in the emergence to early seedling stage. However, in addition to the ground cover, seedlings at TL were subject to drier soil conditions due to the prevalence of large P. radiata roots, shading and obstruction of rainfall from overhanging tree branches, shading from P. radiata needle litter, and possibly an allelopathic effect of the needles (Kimura et al. Reference Kimura, Sato and Kato-Noguchi2015). Thus, fewer seedlings emerged at TL than M2. Also, after the initial emergence, TL seedlings failed to grow taller than an average of 2.4 cm (Figure 2C) or produce multiple stems in the first year, even in the Bare treatment. Survival to 1 yr at TL was almost entirely limited to seedlings in the Bare treatment, which suggests that the presence of grasses and other plant species may still have had a more decisively negative effect on seedling survival than in the other treatments.
Higher mortality of seedlings occurred between 6 mo and 1 yr after sowing at TL than at M2 (27% vs. 76%), which may have been due to a failure to achieve a critical size to tolerate the winter. Notwithstanding their small size (no more than 3-cm tall), a few seedlings at TL did persist through the first winter (1% of all seeds sown; Figure 1C). In contrast to seedlings in M2, which did not survive in treatments where other vegetation was present, some of the persistent TL seedlings occurred in plots that had never been weeded. This was likely due to lower-density ground cover than at M2, which allowed for less crowding and more light available to the seedlings going into winter.
By Week 61 (early in the second year after sowing), new seedlings had emerged in the TL Cut plots (average 0.25 and 1.75 seedlings plot−1 at Week 52 and Week 61, respectively) and the Long plots (0 and 4.5 seedlings plot−1 at Week 52 and Week 61, respectively). Significantly more second-year seedlings were found in the Long plots (P < 0.05) than in the IBare plots at Week 61 (Supplementary Table S4), where all first-year seedlings had died and no new seedlings had emerged. By the end of the second year after sowing (Week 104, September 2022), there were again more seedlings in the Bare treatment than in any other, but so few seedlings persisted that there was no significant difference among treatments (Supplementary Table S5). Also at Week 104, Cut plots averaged 0.75 seedlings plot−1 and Long plots were again at 0. Despite the small flush of seedling emergence in the second year after sowing, 40% fewer seedlings persisted at TL at the end of that year (0.5% of all seeds sown) than at the end of the first year, demonstrating the difficulty of C. vitalba establishment in such an environment. However, even though no weeding was done in the second year after sowing at TL, seedlings began developing multiple stems, indicating that establishment in restrictive habitats can occur on a limited scale.
Zhang et al. (Reference Zhang, Jiang, Wang, Zhang, Gai, Peng, Zhang, Mao, Lou, Mo, Feng and Ming2021) have claimed that C. vitalba leaves are heat tolerant. Indeed, as the summer progressed, seedling leaves in Bare plots at the M1 and M2 sites and most at the TL site began developing reddish-purple coloring around the edges, an indication of anthocyanin production. Anthocyanins are known for helping protect plants against extreme temperatures, UV radiation, free radicals, drought, injury, and soil contamination (Chalker-Scott Reference Chalker-Scott2002; Gould Reference Gould2004; Kovinich et al. Reference Kovinich, Kayanja, Chanoca, Otegui and Grotewold2015; Laxa et al. Reference Laxa, Liebthal, Telman, Chibani and Dietz2019). At TL, overhanging P. radiata branches intercepted much of the direct rainfall TL seedlings would have received, and large P. radiata roots may also have made seedling access to soil resources more difficult. At M1 and M2, Bare seedlings were more exposed to direct, harsh sunlight than in any other treatments. Anthocyanin production may have helped these seedlings survive the accumulating stresses of limited water supply, nutrient stress, and high light intensity. In IBare plots at M2 and TL, as well as in Cut plots at all three sites, C. vitalba seedling numbers declined over the summer (Figure 1). It is possible that the anthocyanin production helped mitigate seedling loss during the summer months, but it could not protect seedlings also impacted by the effects of resource constraints caused by other plants.
Invasion Strategy
Plant establishment from seed in a new location requires successful seed germination, seedling survival, and growth to maturity (Richardson et al. Reference Richardson, Holmes, Esler, Galatowitsch, Stromberg, Kirkman, Pyšek and Hobbs2007). Nearly all C. vitalba seeds are dispersed by early spring, and viable seeds are most likely to successfully germinate in the same season (Jarvis-Lowry et al. Reference Jarvis-Lowry, Harrington, Ghanizadeh and Robertson2024b). However, despite the potential for an early start, even when seeds land in suitable sites for germination, obstacles to seedling emergence and restrictions to resources at the young seedling stage severely limit C. vitalba’s chances for successful establishment from seed. Indeed, even without other vegetation present, less than 40% of sown seeds emerged as seedlings. Therefore, risk of invasion by C. vitalba seed in dense, grassy habitats or other habitats with limited resources is not high.
Maximizing resource availability during vulnerable seedling stages by colonizing bare habitats is a strategy employed by many invasive plants (Gioria and Pyšek Reference Gioria and Pyšek2017), as seems to be the case for C. vitalba. A suitably vacant site may occur along rivers when silt is deposited during a flooding event or when gaps in the forest floor open due to treefalls or other disturbances. Clearing by machinery can also open sites for colonization. Yet if gaps or cleared land are allowed to be quickly recolonized with dense vegetation, C. vitalba seedlings that do emerge are unlikely to reach maturity. However, dense vegetation is not always achievable, and seedlings are capable of colonizing habitats with lower-density ground cover, even where the presence of large tree roots and shading makes establishment yet more problematic. In heavily infested sites, C. vitalba produces approximately 50,000 viable seeds m−2 yr−1 (Jarvis-Lowry Reference Jarvis-Lowry2023). If even as little as 0.5% to 1% of seeds are dispersed to suitably bare sites, and as in this study, fewer than 40% of those seeds germinate in soil that remains bare long enough for seedlings to reach a critical size, up to nearly 200 successful new recruits could still result. Also, dormant seeds in the soil could take advantage of newly bare habitats. Thus, the magnitude of its sexual propagule pressure may help C. vitalba compensate for its low investment in quality seed and establish itself in new locations. Importantly, because of climate change, more extreme and frequent storms and flooding events that generate more vacant sites may improve C. vitalba’s chances of establishing by seed.
Invasive plants often employ effective long-distance dispersal in combination with effective and rapid expansion over shorter distances (Schupp Reference Schupp2011). Other lianas are well known for spreading by clonal stems along the ground (Buru et al. Reference Buru, Dhileepan, Osunkoya and Firn2016; Ladwig and Meiners Reference Ladwig and Meiners2015; Ledo and Schnitzer Reference Ledo and Schnitzer2014; Mori et al. Reference Mori, Ueno, Matsumoto, Kamijo, Tsumura and Masaki2018; Prosperi et al. Reference Prosperi, Caballe and Caraglio2001; Schnitzer et al. Reference Schnitzer, Parren and Bongers2004; Yorke et al. Reference Yorke, Schnitzer, Mascaro, Letcher and Carson2013), and recent research has shown that vegetative spread in C. vitalba is common and vigorous (Jarvis-Lowry et al. Reference Jarvis-Lowry, Harrington, Ghanizadeh and Robertson2024a). Clematis vitalba seeds have the potential for long-distance dispersal via wind and water, which would help spread this invasive species. However, although cohort fitness and annual climate differences could modulate this, the outcome of the current study indicates that relatively few C. vitalba seedlings are expected to survive to maturity within established vegetation. Accordingly, for those few seedlings that do successfully colonize new habitats, vegetative spread may be the primary means of rapidly dominating new sites. That is, sexually reproduced diaspores may be the key to long-distance dispersal of the species, but once established, asexual growth likely allows the plant to entrench and become invasive.
Limitations
This study offers insight into the unexceptional vigor of early C. vitalba seedlings in the presence of established vegetative ground cover, although it did not evaluate the direct effects of competition between C. vitalba and other species, nor did it quantify changes in resource availability.
Summary of Findings
Sowing C. vitalba seeds in early spring resulted in more seedling emergence than sowing in late spring, although cohort fitness may have been a factor.
Survivorship of C. vitalba seedlings was highest, and seedlings were most vigorous, in plots kept bare of other vegetation. Increasing vegetative cover density resulted in lower survival and vigor of C. vitalba, and seedlings did not survive at all in the densest vegetative cover treatment.
-
Lower-density vegetative ground cover under mature P. radiata trees allowed C. vitalba seedlings to emerge in all treatments, yet survival and vigor were both low, due to limited rainfall, sunlight, and soil resources in that environment.
-
The best habitat for establishment of C. vitalba by seed is one in which soil resources and light are not limited, such as in newly bare sites along rivers or in forest gaps and edges. As C. vitalba is such an invasive species at many sites in New Zealand, limited seedling establishment is presumably compensated for by the vigorous vegetative growth from plants that do establish successfully.
Management Implications
Seedling recruitment can be reduced by limiting bare soil and encouraging a dense, desirable ground cover, such as healthy grasses beneath planted shrubs and trees in riparian zones and beneath pines in forestry blocks. In circumstances where seedlings establish in bare sites generated by natural or man-made disturbances, seedlings should be removed completely, so they cannot begin spreading vegetatively.
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.1017/wsc.2024.65
Funding statement
Financial support for this research was given by Massey University, Horizons Regional Council, New Zealand Plant Protection Society, New Zealand Biosecurity Institute, George Mason Charitable Trust, New Zealand Tree Crops Association, and Massey University Seed Tech Services.
Competing interests
The authors declare no conflicts of interest.







 Open access
Open access