



Introduction
Wheat (Triticum aestivum L.) is the most widely grown crop globally, providing 20% of the daily consumed calories and protein content around the world (Huang et al. Reference Huang, Wang, Su, Yu, Shen, He and Gao2022; Luo et al. Reference Luo, Yang, Lin and Xiao2023). With the growing global population and frequent occurrence of extreme weather caused by climate change, ensuring wheat production is essential for food security (Huang et al. Reference Huang, Wang, Su, Yu, Shen, He and Gao2022; Luo et al. Reference Luo, Yang, Lin and Xiao2023). Herbicides are important tools for controlling weeds in farmland and have been widely used globally. From 1990 to 2015, a steady, linear trend of an increasing number of herbicide area treatments was observed for all crops except soybean [Glycine max (L.) Merr.] in the United States (Kniss Reference Kniss2017). Although agricultural production around the world is increasingly reliant on herbicides, we cannot ignore the phytotoxicity of herbicides to non-target plants. The phytotoxicity of herbicides to non-target crops is one of the main obstacles to high-yield cultivation of crops (Varshney et al. Reference Varshney, Hayat, Alyemeni and Ahmad2012). The negative impact of improper use of herbicides on agricultural production is sometimes more severe than that of weeds (de Castro Marcato et al. Reference de Castro Marcato, de Souza and Fontanetti2017; Shaner Reference Shaner2004).
Isoproturon is a phenylurea-derived herbicide that has been widely applied for weed control in wheat fields (Sorensen et al. Reference Sorensen, Ronen and Aamand2001). The phytotoxicity of isoproturon acts through inhibition of photosynthesis, reactive oxygen species (ROS) generation, and damage to defense systems (Hassan and Nemat Alla Reference Hassan and Nemat Alla2022). With increasing weed resistance in wheat fields becomes increasingly severe (Nakka et al. Reference Nakka, Jugulam, Peterson and Asif2019), some farmers may respond to the pressure of controlling resistant weeds by increasing the dosage of isoproturon being used. This undoubtedly increases the risk of herbicide phytotoxicity. If wheat encounters severe weather conditions such as continuous cloudy or low temperatures after isoproturon spraying, even if the application amount is within the recommended dosage range, the impact of low light and low temperature stress on wheat will significantly increase the risk of herbicide phytotoxicity (Gu et al. Reference Gu, Li, Sun, Luo, Liu, Zhang, Qi, Zhao, Tang, Zhang, Shao, Xiong, Si and Zhao2017; Li et al. Reference Li, Cai, Liu, Dai, Cao and Jiang2014; Xiao et al. Reference Xiao, Asseng, Wang, Xia, Zhang, Liu, Tang, Cao, Zhu and Liu2022). In China, herbicide phytotoxicity often occurs in winter wheat fields after isoproturon spraying. Therefore, it makes sense to explore simple and feasible ways to alleviate isoproturon phytotoxicity in wheat.
Plant growth regulators (PGRs) can be defined as natural or synthetic compounds that affect the development or metabolic processes of higher plants and are widely used in modern agricultural production (Rademacher Reference Rademacher2015). Many PGRs or exogenous compounds with plant growth regulatory effects have also been reported to enhance crop resistance to stresses such as salt stress, temperature stress, and water stress, reducing stress-induced damage. They usually promote crop photosynthesis on the one hand and maintain a lower ROS state by increasing the activity of antioxidant enzymes in the crop on the other hand (He et al. Reference He, He, Xu, Zhang, Zhang, Ren, Li, Zhou and Duan2023; Wang et al. Reference Wang, Zhao, Zhao, Gu, Na, Xie, Cheng and Pan2020; Yan et al. Reference Yan, Wei, Ding, Li, Liu, Chen, Tang, Ding, Jiang and Li2021a). Iron chlorin e6 (ICe6) is a new PGR patented by Nanjing Better Bioengineering. As shown in Figure 1, it is formed by chelating pyropheophorbide, purpurin, and dihydroporphyrin as main ligands and different acid radicals or hydroxy radicals serving as axial ligands (X) with the transitional trivalent iron ions (Cao et al. Reference Cao, Yu, Zhao, Wei, Song, Chen and Wang2016). At present, ICe6 is registered in China on crops such as rice (Oryza sativa L.), wheat, and canola (Brassica napus L.), to increase crop yield and improve stress resistance. It has been reported that it can alleviate the damage caused by salt stress by improving the photosynthetic capacity of soybeans and canola, as well as the activity of antioxidant enzymes (Cao et al. Reference Cao, Yu, Zhao, Wei, Song, Chen and Wang2016, Reference Cao, Li, Yu, Shi, Chen and Yu2018). These results fully demonstrate its enormous potential in improving crop stress resistance. Wheat sprayed with isoproturon can be considered to be under a chemical stress (Baillard et al. Reference Baillard, Sulmon, Bittebiere, Mony, Couée, Gouesbet, Delignette-Muller, Devin and Billoir2020). Isoproturon disrupts the photosynthetic system of wheat, weakens its photosynthesis, and reduces the activity of antioxidant enzymes in wheat, inducing oxidative stress and leading to a large accumulation of ROS that ultimately results in growth inhibition and even death (Yin et al. Reference Yin, Jiang, Song and Yang2008). This is very similar to the damage caused by many abiotic stresses to crops, so it is feasible to alleviate the herbicide phytotoxicity of isoproturon to wheat by spraying PGRs or compounds with plant growth regulatory effects. Moreover, previous research reports exogenous application of glutathione, jasmonic acid, or salicylic acid can reduce the phytotoxicity of isoproturon to wheat (Liang et al. Reference Liang, Lu and Yang2012; Ma et al. Reference Ma, Zhang, Zhang, Zhang, Li, Wang, Yu and Yang2018; Nemat Alla and Hassan Reference Nemat Alla and Hassan2014). However, there is currently no research on the use of ICe6 in alleviating herbicide phytotoxicity. Therefore, the aim of this study is to clarify the alleviative effect of ICe6 on isoproturon phytotoxicity to wheat; specifically, to study the effects of ICe6 on the growth parameters, photosynthetic pigment content, photosynthesis, and antioxidant capacity of wheat damaged by isoproturon.

Figure 1. Chemical structure of iron chlorin e6 (ICe6).
Materials and Methods
Plant Materials and Chemicals
Wheat seeds (‘Ning 13’) were purchased from Jiangsu Mingtian Seeds Science and Technology (Nanjing, Jiangsu, China). Isoproturon (50% wettable powder) was purchased from Jiangsu Kuaida Agrochemical (Nantong, Jiangsu, China). ICe6 (purity = 2%) was provided by Nanjing Better Bioengineering (Nanjing, Jiangsu, China) (Figure 1). All other chemicals were purchased from Solarbio Science and Technology (Beijing, China).
Experimental Design
Wheat seeds were sown in square plastic pots (7×7cm) filled with a soil mixture (pure soil:organic substrate = 1:1; organic substrate purchased from Jiang Su Xing Nong Substrate & Technology, Zhenjiang, Jiangsu, China), with 15 seeds per pot. All wheat plants were cultivated in a greenhouse with a temperature of 15/20 C (night/day), a relative humidity of 85%, illumination of 8,000 lux, and a 12/12-h photoperiod. When wheat seedlings grow to the 1-leaf stage, only 10 wheat seedlings of uniform growth were retained in each pot. The pots were divided into 7 groups of 10 pots each. Our preliminary experiment (data not shown) determined that wheat would be susceptible to isoproturon damage at a dose of 2,250 g ai ha−1. When wheat seedlings grow to the 2-leaf stage, they were sprayed with isoproturon (dose of 2,250 g ai ha−1); wheat seedlings sprayed with water were blank control groups. After 7 d, wheat seedlings that were damaged by isoproturon were sprayed with 0, 0.00225, 0.0045, 0.009, 0.018, and 0.036 g ai ha−1 of ICe6. The spraying equipment we used was the 3WP-2000 walking spray tower produced by the Nanjing Mechanization Research Institute of the Ministry of Agriculture in China (spindle speed: 96 mm r−1; spray height: 300 mm; effective spray width of the nozzle: 350 mm; flow rate of the nozzle: 390 ml min−1; travel distance: 1,340 mm; volume of liquid solution: 30 ml). Of the 10 wheat pots from each treatment, 5 pots were randomly selected for sample collection at 0, 7, and 14 d after spraying with ICe6, and then 3 pots were randomly selected for the determination of growth parameters, and 5 randomly selected wheat plants from the remaining 2 pots were used for the measurement of photosynthesis.
Determination of Growth Parameters
Plant height was measured, aboveground biomass was collected, and fresh weight (FW) was recorded at 2 wk after ICe6 application. This experiment was a completely randomized design with three replicates (3 pots each containing 10 wheat plants) set up for each treatment.
Determination of Photosynthetic Parameters
Photosynthetic parameters were measured at 14 d after ICe6 treatment. Photosynthetic parameters such as photosynthetic rate (P n), stomatal conductance (G s), transpiration rate (T r), intercellular CO2 concentration (C i), photosystem II (PSII) maximum photochemical quantum yield (F v/F m), and PSII quantum efficiency (ΦPSII) were evaluated using an automatic measurement system for photosynthesis (LI-6800, Li-Cor, Lincoln, NE, USA). The LI-6800 environmental conditions were set as flow: 500 μmol s−1; H2O: 50%; CO2: 400 μmol mol−1; fan: 10,000 rpm; temperature: 20 C; light: 1,200 μmol m−2 s−1. The unit of measurement for photosynthetic parameters was the penultimate leaf of each wheat plant. This experiment was a completely randomized design with five replicates set up for each treatment.
Determination of Chlorophyll Content and Carotenoid Content
Wheat samples (0.2 g) were collected at 0, 7, and 14 d after ICe6 treatment and were placed in 10 ml of ethanol acetone mixture (ethanol:acetone = 1) for chlorophyll extraction. OD633, OD470, and OD645 of the mixture were measured after 7 d. The concentrations of extracted photosynthetic pigments were calculated based on the three absorbances, according to Szwed et al. (Reference Szwed, Mitrus, Wiczkowski, Dębski and Horbowicz2020). This experiment was a completely randomized design with three replicates set up for each treatment.
Chlorophyll content was calculated using the following equations:
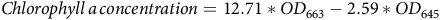 $Chlorophyll \,a \,concentration=12.71*OD_{663}-2.59*OD_{645}$
$Chlorophyll \,a \,concentration=12.71*OD_{663}-2.59*OD_{645}$
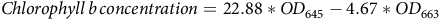 $Chlorophyll \,b \,concentration=22.88*OD_{645}-4.67*OD_{663}$
$Chlorophyll \,b \,concentration=22.88*OD_{645}-4.67*OD_{663}$
 $$\begin{gathered}Total{\,}chlorophyll{\,}content\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\hfill \\ = (8.04*O{D_{663}} + 20.29 * O{D_{645}}) * V/1,000W \hfill \\ \end{gathered}$$
$$\begin{gathered}Total{\,}chlorophyll{\,}content\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\hfill \\ = (8.04*O{D_{663}} + 20.29 * O{D_{645}}) * V/1,000W \hfill \\ \end{gathered}$$
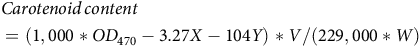 $$\begin{gathered}Carotenoid{\,} content \,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\hfill \\ = (1,000*O{D_{470}} - 3.27X - 104Y)*V/(229,000 * W) \hfill \\ \end{gathered}$$
$$\begin{gathered}Carotenoid{\,} content \,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\,\hfill \\ = (1,000*O{D_{470}} - 3.27X - 104Y)*V/(229,000 * W) \hfill \\ \end{gathered}$$
where V is the volume of the supernatant, W is the fresh weight (g) of the sample, X is the chlorophyll a concentration, and Y is the chlorophyll b concentration.
Determination of Malondialdehyde (MDA) Content
The MDA measurement method was based on previous work by Wang et al. (Reference Wang, Wang, Li, Yu, Lin and Dong2022) with slight modifications. Wheat samples (0.2 g) were collected 0, 7, and 14 d after ICe6 treatment. Wheat leaves (0.2 g) were homogenized in 1.5 ml of 50 g L–1 trichloroacetic acid (TCA) solution. The homogenate was centrifuged at 12,000 × g for 20 min at 4 C in a refrigerated centrifuge, and 0.5 ml of supernatant was added to 2 ml of 5 g L–1 thiobarbituric acid in 200 g L–1 TCA. The mixture was heated in a block bath at approximately 99 C for 30 min, and the reaction was stopped in an ice bath for 10 min. Following centrifugation at 12,000 × g for 5 min, the supernatant (0.2 ml) was collected and the absorbance was measured at 450, 532, and 600 nm. This experiment was a completely randomized design with three replicates set up for each treatment.
 $MDA \,concentration=6.45*(OD_{532}- OD_{600}) - 0.56*OD_{450}$
$MDA \,concentration=6.45*(OD_{532}- OD_{600}) - 0.56*OD_{450}$
 $MDA \,content = C*V/W $
$MDA \,content = C*V/W $
where C is the MDA concentration, V is the volume of the supernatant, and W is the fresh weight of the sample.
Determination of Hydrogen Peroxide (H 2 O 2 ) Content
Wheat samples (0.2 g) were collected 0, 7, and 14 d after ICe6 treatment. Wheat leaves (0.2 g) were homogenized in 1.5 ml of 1 g L–1 TCA solution. The homogenate was centrifuged at 12,000 × g for 15 min at 4 C in a refrigerated centrifuge, and 0.5 ml of supernatant was added to a mixture containing 0.5 ml of 10 mM phosphate buffer (pH 7.0) and 1 ml of 1 mol L−1 KI. The absorbance was measured at 390 nm after mixing. The H2O2 concentration was measured according to the standard curve (Liu et al. Reference Liu, Wu, Naeem, Liu, Deng, Xu, Zhang and Zhou2013). This experiment was a completely randomized design with three replicates set up for each treatment.
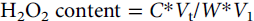 $${{\text{H}}_{\text{2}}}{{\text{O}}_{\text{2}}}{\text{ content} {= C^*}}{{{V}}_{{t}}}{{/W^*}}{{{V}}_{\text{1}}}$$
$${{\text{H}}_{\text{2}}}{{\text{O}}_{\text{2}}}{\text{ content} {= C^*}}{{{V}}_{{t}}}{{/W^*}}{{{V}}_{\text{1}}}$$
where C is the H2O2 concentration, V t is the total volume of sample extraction solution, V 1 is the volume of sample extraction solution used for measurement, and W is the fresh weight of the sample.
Determination of Antioxidant Enzyme Activity
Wheat samples (0.2 g) were collected 0, 7, and 14 d after ICe6 treatment. Enzyme extractions were performed according to the method used by Li et al. (Reference Li, Qiu, Zhang and Wang2010) with slight modifications. Wheat leaves (0.2 g) were lyophilized in liquid nitrogen and ground into powder by a plant tissue grinder and then added to 2-ml centrifuge tubes containing 2 ml of 50 mmol L−1 phosphate buffer (pH 7.8), 10 g L–1 polyvinylpyrrolidone, and 1 mM ethylenediaminetetraacetic acid disodium salt (EDTA·Na2). The homogenate was centrifuged at 12,000 rpm for 20 min at 4 C in a refrigerated centrifuge, and the resulting supernatant (0.2 ml) was used to assay of superoxide dismutase (SOD), catalase (CAT), peroxidase (POD), and ascorbate peroxidase (APX) activities. The following experiments were conducted in a completely randomized design with three replicates set up for each treatment.
The SOD activity was assayed using the photochemical nitrogen blue tetrazolium (NBT) method (Beauchamp and Fridovich Reference Beauchamp and Fridovich1971). The reaction mixture containing 50 mmol L−1 of phosphate buffer (pH 7.8), 130 mmol L−1 of methionine, 750 μmol L−1 of NBT, 20 μmol L−1 of riboflavin, 0.1 mmol L−1 of EDTA·Na2, and 25 μl of enzyme extract in a 1.5-ml volume was placed under light and reacted for 20 min. The absorbance of the solution was measured at 560 nm. One unit (U) of SOD was defined as the enzyme activity that inhibited the photoreduction of NBT to blue formazan by 50%.
The POD activity was measured with guaiacol as the substrate with some modifications (Kwak et al. Reference Kwak, Kim, Lee, Jung, Park and Liu1995). The reaction mixture (200 μl) consisted of 200 mmol L−1 of sodium acetate buffer (pH 6), 0.25% guaiacol, 0.3% H2O2, and 5 μl of enzyme extract. The increase in absorbance due to oxidation of guaiacol was measured at 470 nm for 3 min. One unit of POD activity was defined as a change in absorbance of 0.1 U min−1.
The CAT activity was assayed spectrophotometrically at 240 nm in a 200-μl reaction mixture containing 10 μl of enzyme extract, 100 μl of 150 mmol L−1 phosphate buffer (pH 7.0), and 90 μl of 30% H2O2 (Cakmak and Marschner Reference Cakmak and Marschner1992). The decomposition of H2O2 was measured by following the decrease in absorbance at 240 nm for 3 min. One unit of CAT activity was defined as a change in absorbance of 0.1 U min−1 caused by the addition of the enzyme extract.
The APX activity was assayed in a reaction mixture of 200 μl containing 175 μl of 100 mmol L−1 phosphate (pH 7.0), 1 μl of 100 mmol L−1 l-ascorbic acid, 19 μl of H2O2, and 5 μl of enzyme extract. The reaction was started by adding enzyme extract to the mixture. Enzyme activity was quantified by following the decrease in absorbance at 290 nm for 3 min (Jiang and Zhang Reference Jiang and Zhang2002). One unit of APX activity was defined as an absorbance change of 0.1 U min−1.
Data Analysis
All results are presented as the mean ± SE of at least three replicates, and all experiments were repeated at least once. All data were subjected to ANOVA, and Duncan’s multiple-range test (P < 0.05) was used in SPSS v. 21.0 (SPSS, Chicago, IL, USA). Data charts were made using SigmaPlot v. 12.0 (SigmaPlot Software, Chicago, IL, USA).
Results and Discussion
Alleviative Effect on Isoproturon Phytotoxicity to Wheat
The results of the whole-plant bioassay confirmed that ICe6 has an alleviative effect on isoproturon phytotoxicity to wheat (Figure 2; Table 1). Fourteen days after treatment with ICe6 (21 d after treatment with isoproturon), the phenotype of the isoproturon-treated wheat showed plant tissue wilting and leaf chlorosis, and the shoot fresh weight and shoot height of the isoproturon-treated wheat were significantly inhibited compared with the control, with a shoot fresh weight inhibition rate of 57.62% and a plant height inhibition rate of 26.87%. However, all the isoproturon+ICe6–treated wheat showed recovery in plant height and fresh weight compared with the isoproturon-treated wheat, with the isoproturon+ICe6–treated (0.018 g ai ha−1) wheat having the most significant recovery effect. The phenotype of the isoproturon+ICe6–treated (0.018 g ai ha−1) wheat did not show plant tissue wilting and leaf chlorosis, and the fresh weight inhibition rate was limited to 20.20%, and the plant height inhibition rate was limited to 8.20%.

Figure 2. (A) Phenotype of wheat seedlings after 7 d of treatment with isoproturon (2,250 g ai ha−1). (B and C) Phenotype of wheat seedlings after 14 d of treatment with iron chlorin e6 (ICe6) at a dose of 0.018 g ai ha−1; i.e., 21 d of treatment with isoproturon (2,250 g ai ha−1). The dose of ICe6 was 0.018 g ai ha−1. CK, control wheat seedlings treated with water; Isoproturon, wheat seedlings treated with isoproturon; and Isoproturon + ICe6, wheat seedlings treated with isoproturon followed by ICe6.
Table 1. The alleviating effect of different doses of iron chlorin e6 (ICe6) on isoproturon phytotoxicity to wheat a
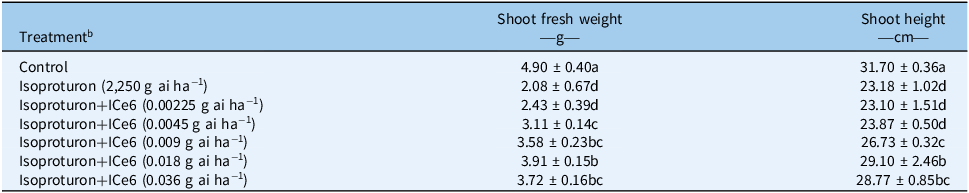
a Data in the table were collected 14 d after ICe6 treatment. Different letters in the same column indicate significant differences at the P < 0.05 level by Duncan’s test. Numbers after “±” indicate standard deviation.
b “Isoproturon + ICe6” means that wheat seedlings were treated with isoproturon (2,250 g ai ha−1) for 7 d followed by ICe6 for 14 d.
Herbicide damage to crops has always been a concern for farmers in chemical control of weeds in farmland. Previous studies have shown that high doses of isoproturon inhibit the growth of wheat seedlings (Ma et al. Reference Ma, Zhang, Zhang, Zhang, Li, Wang, Yu and Yang2018; Nemat Alla and Hassan Reference Nemat Alla and Hassan2014; Yin et al. Reference Yin, Jiang, Song and Yang2008). Isoproturon damage to winter wheat occurs more frequently in in China, so it is crucial to explore a convenient and feasible method to alleviate isoproturon phytotoxicity to wheat (Deng Hailing et al. Reference Deng Hailing, Zhang, Qiang and Song2022; Meng Dandan et al. Reference Meng Dandan, Guo, Ma, Chen, Peng and Li2019). ICe6, as a PGR, has been previously reported to enhance crop stress resistance under salt stress, but its potential for other stress resistance needs to be developed (Cao et al. Reference Cao, Yu, Zhao, Wei, Song, Chen and Wang2016, Reference Cao, Li, Yu, Shi, Chen and Yu2018). This study is the first report of the alleviative effect of ICe6 on isoproturon phytotoxicity to wheat and its optimal dosage for use. We found that ICe6 (0.018 g ai ha−1) significantly increased plant height and fresh weight of isoproturon-treated wheat (Table 1), and phenotypically, its alleviative effect on isoproturon phytotoxicity to wheat was significant (Figure 2). Although compounds such as glutathione and jasmonic acid found in previous studies are also very effective in alleviating the isoproturon phytotoxicity to wheat, they are not commercialized pesticides and may be difficult to promote in agricultural production at present (Ma et al. Reference Ma, Zhang, Zhang, Zhang, Li, Wang, Yu and Yang2018; Nemat Alla and Hassan Reference Nemat Alla and Hassan2014). In summary, ICe6 (0.018 g ai ha−1) has a significant alleviative effect on isoproturon phytotoxicity to wheat, and the findings of our study provide a feasible measure to alleviate the isoproturon phytotoxicity to wheat.
Effect of ICe6 on Antioxidant Capacity
MDA Content and H 2 O 2 Content
After 7, 14, and 21 d of treatment with isoproturon, the MDA content and H2O2 content of isoproturon-treated wheat gradually increased and were significantly higher than those of the control, while the MDA content and H2O2 content of the isoproturon+ICe6–treated (0.018 g ai ha−1) wheat did not continue to increase, and the H2O2 content and MDA content were significantly lower than those of the isoproturon-treated wheat at 14 d after ICe6 treatment and the difference with control was not significant (Figure 3).
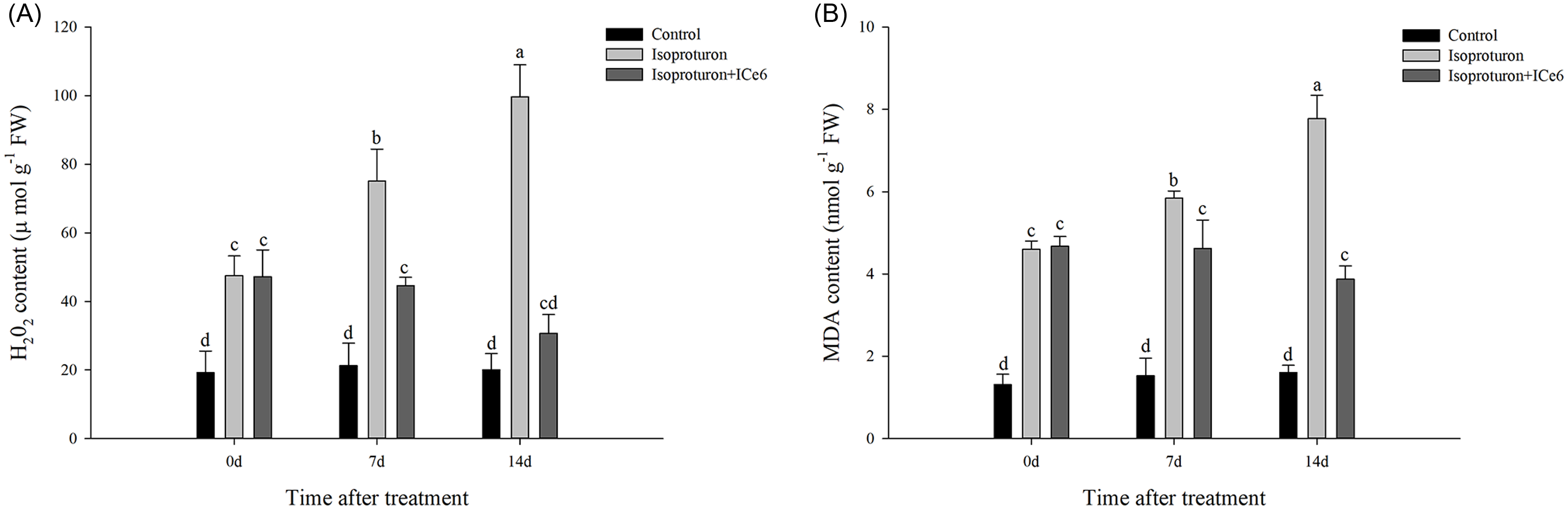
Figure 3. Effect of iron chlorin e6 (ICe6) treatment (0.018 g ai ha−1) on (A) hydrogen peroxide (H2O2) and (B) malondialdehyde (MDA) content of wheat damaged by isoproturon (2,250 g ai ha−1) during the subsequent 14 d. Points with the same letter are not significantly different at P < 0.05.
Isoproturon inhibits electron transfer in PSII, which prevents the absorption of light energy from being converted into electrochemical energy and produces triplet chlorophyll and singlet oxygen, leading to a large amount of ROS production. A large amount of ROS reacts with lipids, leading to membrane lipid peroxidation and metabolic disorders, as well as damage to cell structure (Fuerst and Norman Reference Fuerst and Norman2017; Gill and Tuteja Reference Gill and Tuteja2010; Rutherford and Krieger-Liszkay Reference Rutherford and Krieger-Liszkay2001). H2O2 is an important reactive oxygen component. MDA is the final product of membrane lipid peroxidation, and its content can reflect the level of membrane lipid peroxidation damage. In the present study, we found that high-dose isoproturon treatment for 21 d resulted in a significant accumulation of H2O2 and MDA in wheat, proving that isoproturon indeed induces oxidative stress and leads to membrane lipid peroxidation. However, the H2O2 and MDA content in wheat sprayed with ICe6 (0.018 g ai ha−1) after 7 d of treatment with isoproturon did not continue to increase and decreased slightly over time. In summary, ICe6 limited the generation of H2O2 and MDA and alleviated membrane lipid peroxidation damage in wheat cells.
Antioxidant Enzyme (SOD, POD, CAT, and APX) Activity
Seven days after treatment with isoproturon, the SOD activity of isoproturon-treated and isoproturon+ICe6–treated (0.018 g ai ha−1) wheat was higher than that of the control (Figure 4). There was no significant difference in POD and APX activity compared with the control, while CAT activity was lower than that of control. On the following 14 and 21 d, all antioxidant enzyme activity in the isoproturon-treated wheat decreased, while SOD activity in the isoproturon+ICe6–treated (0.018 g ai ha−1) wheat slowly decreased and remained higher than in the control. POD and CAT activity first increased and then decreased without significant difference from the control, while APX activity first decreased and then increased but also showed no significant difference from the control. After 7 and 14 d of treatment with ICe6, the activity of the four antioxidant enzymes in isoproturon+ICe6–treated (0.018 g ai ha−1) wheat was significantly higher than in isoproturon-treated wheat.
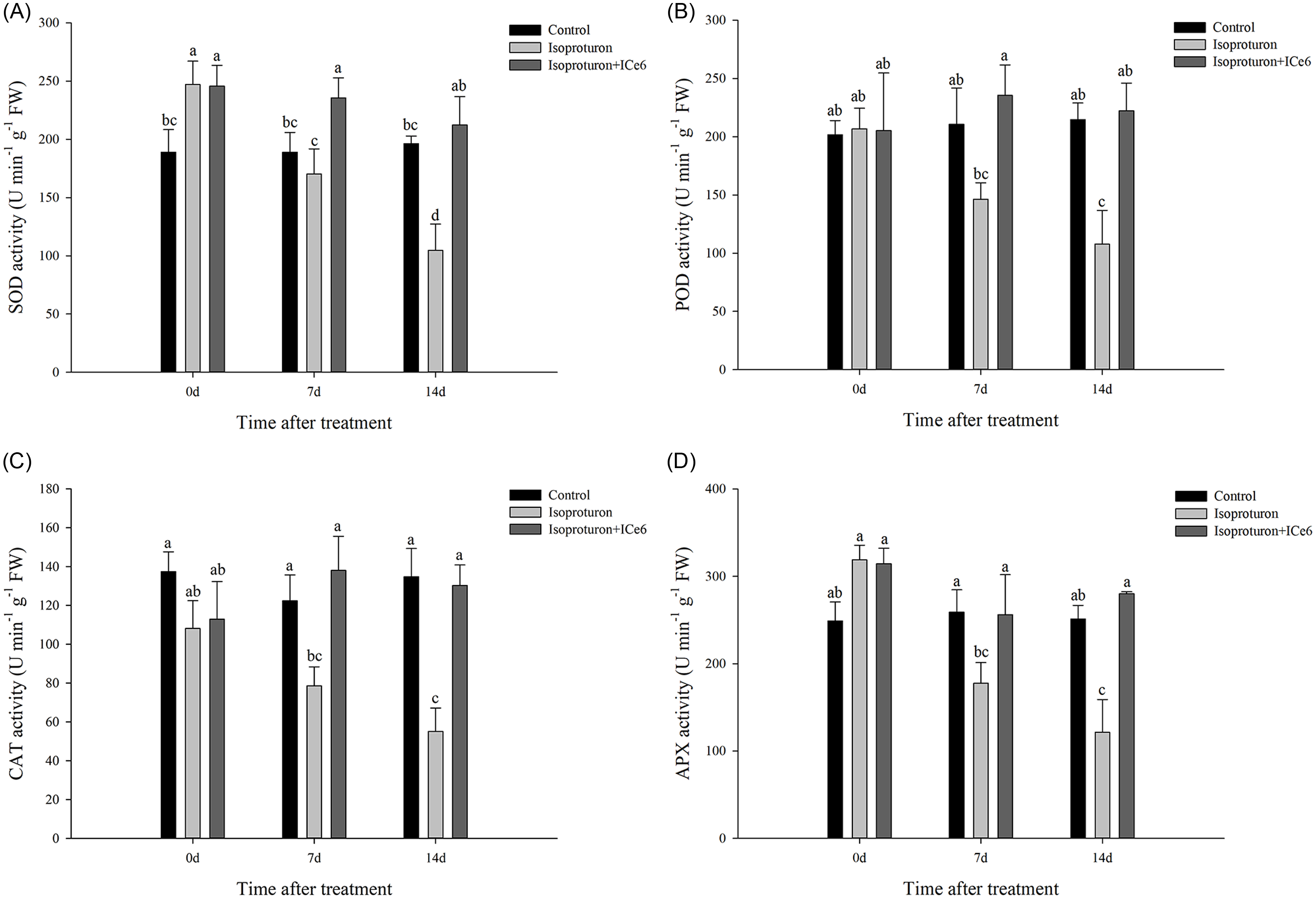
Figure 4. Effect of iron chlorin e6 (ICe6) treatment (0.018 g ai ha−1) on (A) superoxide dismutase (SOD), (B) catalase (CAT), (C) peroxidase (POD), and (D) ascorbate peroxidase (APX) activity of wheat damaged by isoproturon (2,250 g ai ha−1) during the subsequent 14 d. Points with the same letter are not significantly different at P < 0.05.
Plant antioxidant enzymes (SOD, POD, CAT, and APX) play a very important role in clearing ROS. SOD can catalyze O2 .− to produce H2O2, followed by POD, CAT, and APX catalyzing H2O2 to produce nontoxic and harmless H2O (Foyer and Noctor Reference Foyer and Noctor2009). In the present study, we found that the activities of SOD, POD, and APX in wheat were not lower than those of the control after 7 d of treatment with isoproturon. This is similar to the results of Ma et al. (Reference Ma, Zhang, Zhang, Zhang, Li, Wang, Yu and Yang2018) and Yin et al. (Reference Yin, Jiang, Song and Yang2008), but different from the consistent decrease in antioxidant enzyme activity of Nemat Alla and Hassan (Reference Nemat Alla and Hassan2014). We speculate that the differences may be caused by different leaf ages of the wheat tested. The ROS content was positively related to the H2O2 and MDA content, and we found that the H2O2 and MDA content (isoproturon-treated wheat) continued to increase over time, while their antioxidant enzyme activities continued to decline. Therefore, we speculate that this may be due to the long-term and excessive generation of ROS caused by isoproturon (2,250 g ai ha−1), ultimately leading to the destruction of the antioxidant enzyme itself by the accumulated ROS and thus its inability to continue to function (Nemat Alla and Hassan Reference Nemat Alla and Hassan2014; Yin et al. Reference Yin, Jiang, Song and Yang2008). CAT may be more sensitive to these injuries, so its decline in activity was advanced (Alscher et al. Reference Alscher, Erturk and Heath2002; Chagas et al. Reference Chagas, Silveira, Ribeiro, Vitorello and Carrer2008; Yin et al. Reference Yin, Jiang, Song and Yang2008). However, the activity of antioxidant enzymes (SOD, POD, CAT, and APX) in wheat sprayed with ICe6 after 7 d of treatment with isoproturon did not significantly decrease. This is similar to the results of Cao et al.’s (Reference Cao, Yu, Zhao, Wei, Song, Chen and Wang2016, Reference Cao, Li, Yu, Shi, Chen and Yu2018) research on the effect of ICe6 on salt-stressed soybeans and canola. ICe6 is a chlorophyll derivative that has been proven to have good antioxidant properties when applied to crops, specifically in canola and soybean (Cao et al. Reference Cao, Yu, Zhao, Wei, Song, Chen and Wang2016, Reference Cao, Li, Yu, Shi, Chen and Yu2018). Therefore, we speculate that ICe6 can effectively eliminate ROS by increasing the antioxidant enzyme activity of wheat under isoproturon stress. A widely studied chlorophyll derivative, chlorophyllin, the sodium-copper salt and the water-soluble analogue of the ubiquitous green pigment chlorophyll, as a highly effective antioxidant, has been shown to possesses higher antioxidant ability than Ascorbic Acid (AsA), Glutathione (GSH), mannitol, and tert-butanol at equimolar concentrations for protecting mitochondria, inhibiting the mutagenicity of various chemicals in bacteria or mice (Cao et al. Reference Cao, Li, Yu, Shi, Chen and Yu2018; Kamat et al. Reference Kamat, Boloor and Devasagayam2000). Therefore, we speculate that ICe6 may also act as an antioxidant, similar to AsA or GSH, directly participating in the clearance of excessive ROS in wheat under isoproturon stress (Cao et al. Reference Cao, Yu, Zhao, Wei, Song, Chen and Wang2016; Mittler Reference Mittler2002), thereby alleviating membrane lipid peroxidation damage and greatly maintaining cell membrane integrity. In summary, ICe6 increased the antioxidant capacity of wheat damaged by isoproturon.
Effects of ICe6 on Photosynthesis
Chlorophyll Content and Carotenoid Content
As shown in Figure 5, after 7 to 21 d of treatment with isoproturon, the chlorophyll and carotenoid content of the isoproturon-treated wheat gradually decreased, while the chlorophyll and carotenoid content of the isoproturon+ICe6–treated (0.018 g ai ha−1) wheat were lower than in the control but did not continue to decrease. After 7 and 14 d of treatment with ICe6, the chlorophyll content of the isoproturon+ICe6–treated (0.018 g ai ha−1) wheat was significantly higher than that of the isoproturon-treated wheat. After 14 d of treatment with ICe6, the carotenoid content in the isoproturon+ICe6–treated (0.018 g ai ha−1) wheat was significantly higher than that in the isoproturon-treated wheat.
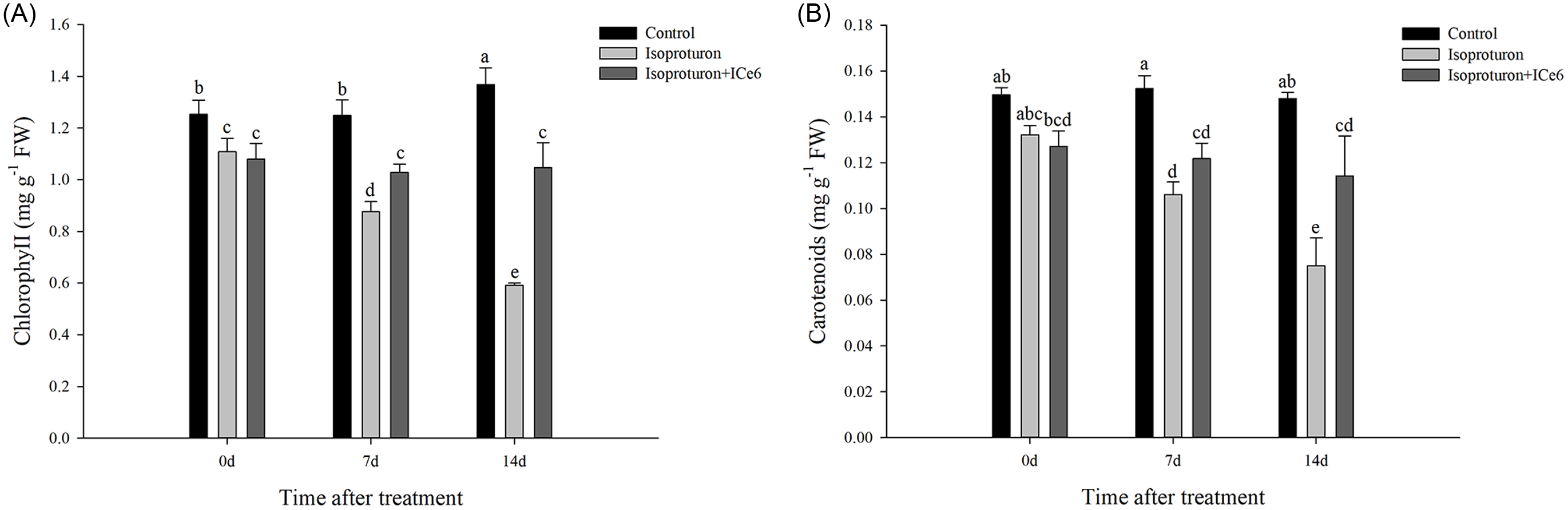
Figure 5. Effect of iron chlorin e6 (ICe6) treatment (0.018 g ai ha−1) on (A) chlorophyll and (B) carotenoid content of wheat damaged by isoproturon (2,250 g ai ha−1) during the subsequent 14 d. Points with the same letter are not significantly different at P < 0.05.
Carotenoids protect chlorophylls and chloroplasts from photooxidation, and chlorophyll is a necessary catalyst for plant photosynthesis. It is closely involved in various aspects of photosynthesis, especially playing a core role in light absorption during photosynthesis (Gutierrez-Gamboa et al. Reference Gutierrez-Gamboa, Marin-San Roman, Jofre, Rubio-Breton, Perez-Alvarez and Garde-Cerdan2018; Katz et al. Reference Katz, Norris, Shipman, Thurnauer and Wasielewski1978; Nemat Alla et al. Reference Nemat Alla, Hassan and El-Bastawisy2008). Previous studies have shown that isoproturon destroys carotenoids in plants and that chlorophyll, deprived of carotenoid protection, is quite sensitive to isoproturon (Nemat Alla and Hassan Reference Nemat Alla and Hassan2014; Yin et al. Reference Yin, Jiang, Song and Yang2008). A similar result was obtained in our study, in which the isoproturon phytotoxicity significantly reduced the chlorophyll and carotenoid content in wheat. However, ICe6 limited the decrease in chlorophyll content and carotenoid content. Excessive ROS cause damage to organelles and intracellular macromolecules, leading to a decrease in photosynthetic pigment content (Nemat Alla and Hassan Reference Nemat Alla and Hassan2014). It can be seen that ROS production was inhibited (Figure 3), while at the same time the decrease in photosynthetic pigment content was inhibited (Figure 5). Therefore, we hypothesize that the reason for this result may be the removal of excess ROS. In summary, ICe6 limited the decrease in chlorophyll and carotenoid content of wheat damaged by isoproturon.
Photosynthetic Parameters
Fourteen days after treatment with ICe6 (21 d after treatment with isoproturon), the P n, G s, T r, and ΦPSII of the isoproturon-treated wheat and the isoproturon+ICe6–treated (0.018 g ai ha−1) wheat were lower than in the control, but the P n, G s, T r, and ΦPSII of the isoproturon+ICe6–treated wheat were significantly higher than those of the isoproturon-treated wheat (Figure 6). The F v/F m of the isoproturon-treated wheat was significantly lower than that of control and the isoproturon+ICe6–treated (0.018 g ai ha−1) wheat. The C i of the isoproturon-treated wheat was significantly higher than that of the isoproturon+ICe6–treated (0.018 g ai ha−1) wheat and control.

Figure 6. Effect of iron chlorin e6 (ICe6) treatment (0.018 g ai ha−1) on photosynthesis of wheat damaged by isoproturon (2,250 g ai ha−1). The measured photosynthetic parameters include (A) net photosynthetic rate (P n), (B) stomatal conductance (G s), (C) transpiration rate (T r), (D) intercellular CO2 concentration (C i), (E) maximum photochemical efficiency (F v/F m), and actual photochemical efficiency (ΦPSII). The measurement was conducted after 14 d of ICe6 treatment. Points with the same letter are not significantly different at P < 0.05.
Photosynthetic decline is a physiological injury common to plants under abiotic or biotic stresses (Cao et al. Reference Cao, Yu, Zhao, Wei, Song, Chen and Wang2016). Isoproturon is a photosynthetic inhibitor that inevitably has a negative impact on crop photosynthesis in weed control. Previous studies have reported that ICe6 can restore photosynthesis in soybean and canola under salt stress (Cao et al. Reference Cao, Yu, Zhao, Wei, Song, Chen and Wang2016, Reference Cao, Li, Yu, Shi, Chen and Yu2018).Thus, we measured the photosynthetic parameters. We found that P n and G s were significantly lower, while C i was significantly higher in isoproturon-treated wheat, suggesting that the reduction in P n was mainly caused by nonstomatal limitation. And isoproturon+ICe6–treated (0.018 g ai ha−1) wheat showed significantly higher P n and G s and lower C i, which may be due to an increase in the rate of CO2 assimilation at a lower CO2 concentration, suggesting that ICe6 regulates nonstomatal limitation. Closing stomata to reduce water transpiration is one of the common strategies used by plants in response to external stresses (Yan et al. Reference Yan, Zhang, Li, Ding, Zhong, Xu, Wei and Li2021b). Isoproturon+ICe6–treated (0.018 g ai ha−1) wheat maintained a higher G s, which was consistent with higher T r, suggesting that ICe6 enhanced the water uptake capacity of isoproturon-treated wheat roots. Our results showed that ICe6 mitigated the decrease of F v/F m and ΦPSII in isoproturon-treated wheat, suggesting that ICe6 can mitigate the effect of isoproturon on PSII in wheat, which may be due to the limitation of the decrease of photosynthetic pigment content by ICe6. In summary, ICe6 restored the photosynthesis of wheat damaged by isoproturon.
In conclusion, the present study found that the severe phytotoxicity of isoproturon to wheat manifested as inhibition of growth, wilting of plant tissue, and leaf chlorosis was alleviated after spraying ICe6 (0.018 g ai ha−1) in the early stage. The alleviative effect is mainly achieved through enhancement of photosynthesis and antioxidant capacity. The present study provides a remedial measure for wheat damaged by isoproturon and expands the application scope of ICe6.
Funding statement
This work was supported by the National Key Research and Development Program of China (2023YFD1400501).
Competing interests
The author(s) declare no competing interests.







