


Scholars studying apostasy — defined as a disengagement from religious belief, identity and/or practice — are increasingly focused on its etiology. Generally, these studies focus on social and psychological causes for apostasy, such as disagreements with religious teachings (Babinski, Reference Babinski1995) and trauma from the death of loved ones (Zuckerman et al., Reference Zuckerman, Galen and Pasquale2016, pp. 97–98 and 100–101). However, there are several potential pathways that may link genetic factors with disengagement from religion, which would constitute a genetic etiology of apostasy.
First, in highly religious societies such as the USA, apostasy constitutes a form of risk-taking behavior. Apostates in highly religious societies risk losing access to important religion-based resources and/or experiencing trauma related to leaving religion (Zuckerman et al., Reference Zuckerman, Galen and Pasquale2016, p. 104). Therefore, genetic factors linked to risky behaviors could at least partially explain variance in the likelihood of becoming an apostate. This is particularly likely to be the case during the transition from adolescence to early adulthood when genes cause neurological changes that have been linked to increases in the levels of risk-taking behaviors such as delinquency and substance use (Chibbar et al., Reference Chibbar, Toma, Mitchell and Miller1990; Cicchetti & Dawson, Reference Cicchetti and Dawson2002; Gardner, Reference Gardner and Elster1999; Guo et al., Reference Guo, Roettger and Cai2008; Steinberg, Reference Steinberg2008).
Second, apostasy during the transition from adolescence to early adulthood could be characterized as a form of an externalizing behavior — defined as overt behaviors that have a negative effect on the external environment. This is because apostasy may constitute a rebellion against parental or societal authority. Because externalizing behaviors have been found to have a genetic etiology (Guo et al., Reference Guo, Roettger and Cai2008; Hendershot et al., Reference Hendershot, Bryan, Feldstein Ewing, Claus and Hutchison2011; Koenig et al., Reference Koenig, McGue and Iacono2008; Krueger et al., Reference Krueger, Hicks, Patrick, Carlson, Iacono and McGue2002; Young et al., 2000, Reference Young, Friedman, Miyake, Willcutt, Corley, Haberstick and Hewitt2009), it is possible that genetic factors that influence the likelihood of becoming an apostate during the transition from adolescence to early adulthood also influence apostasy during that same period.
Lastly, studies have shown that some factors correlated with apostasy have significant heritability. Therefore, genes that influence those factors may also explain variance in the levels of apostasy (Freese, Reference Freese2008).
The present study explores whether or not genetic factors at least partially explain variability in the likelihood of being an apostate during the transition from adolescence to early adulthood. I utilize univariate ACE models to decompose the components of the variance for three measures of apostasy: disengagement from religious institutions, cessation of prayer and religious disaffiliation. The hypothesis that I test in this study is that genetic factors at least partially explain the variance of all three measures of apostasy during the transition between adolescence and early adulthood.
Pathways Linking Genetic Factors and Apostasy
Apostasy as a Risk-taking Behavior
Various scholars have conceptualized disengagement from religious belief, identity and/or practice as a form of risk-taking behavior (Freese, Reference Freese2004; E. Y. Liu, Reference Liu2010; Miller, Reference Miller2000; Miller & Hoffmann, Reference Miller and Hoffmann1995; Miller & Stark, Reference Miller and Stark2002; Sinha et al., Reference Sinha, Cnaan and Gelles2007). Their main argument is that risk-averse religious adherents may avoid irreligiousness because they perceive the risks of not being religious when in fact religious beliefs are correct (e.g., eternal damnation) as far greater than the risks of being religious when in fact religious beliefs are incorrect — what has been described as ‘Pascal’s wager’, named for the French philosopher Blaise Pascal. Miller and Hoffmann (Reference Miller and Hoffmann1995) put forward this argument and found support for it in a sample of high school seniors. Within that sample they found risk preference to be inversely associated with levels of religiosity.
In addition to the perceived risks of negative consequences of apostasy, risk-averse religious adherents may choose to remain religious due to the fear of losing access to important forms of material and social capital. Studies show that religious bodies provide a variety of material and social capital benefits for religious adherents, such as childcare services, counseling and employment opportunities (Coleman, Reference Coleman1988; Emerson & Smith, Reference Emerson and Smith2000, pp. 161–162). Apostasy can lead to ostracism from religious communities, which may prevent access to these valuable resources. Evidence for this comes from Park and Sharma (Reference Park and Sharma2016). They found that individuals who are not affiliated with religion have lower levels of parental network closure, a form of social capital that indicates closeness in an individual’s social network, than individuals with a religious affiliation.
Lastly, religious adherents may avoid apostasy because they fear the possibility of experiencing trauma when attempting to leave religion. This experience of trauma is particularly likely in societies characterized by high levels of religiosity such as the USA. According to Zuckerman et al. (Reference Zuckerman, Galen and Pasquale2016), ‘Americans described their apostasy as being quite intense. It entailed a real personal struggle. It was a dramatic, transformative, life-changing experience that they recalled as being very significant and even painful’ (p. 104). Individuals who live in societies characterized by high levels of religiosity and choose to leave a religion face being rejected by loved ones, physical violence or even death (Theodorou, Reference Theodorou2016).
Genetic etiology of apostasy as a risk-taking behavior
If the conceptualization of apostasy as a risk-taking behavior is correct, genetic factors may influence the etiology of apostasy through the initiation of neurological changes that increase levels of impulsivity. According to Steinberg (Reference Steinberg2008), during puberty the density of receptors in the nucleus accumbens region of the prefrontal cortex are reduced, while the density of myelinated axons (white matter) leading into the prefrontal cortex increases. These changes lead to increased levels of circulating dopamine in the prefrontal cortex and increased rewards from risky behaviors, which can lead to an increased prevalence of risk-taking behaviors such as reckless driving, risky sexual behaviors, delinquency and substance use (Steinberg, Reference Steinberg2008). These neurological changes have been found to have a genetic etiology (Chibbar et al., Reference Chibbar, Toma, Mitchell and Miller1990; Cicchetti & Dawson, Reference Cicchetti and Dawson2002; Gardner, Reference Gardner and Elster1999). Therefore, any risk-taking behaviors predicted by these neurological changes, which may include apostasy, would also have a genetic etiology.
Religiosity as an Externalizing Behavior
Externalizing behaviors have been described as overt behaviors that have a negative effect on the external environment. They include disruptive, hyperactive and aggressive behaviors, including delinquency (J. Liu, Reference Liu2004; White & Renk, Reference White and Renk2012). Individuals exhibiting externalizing behaviors may be more likely to rebel against various types of authority, which may manifest itself in increased levels of delinquency or parent–child discord.
In the USA, apostasy can be characterized as a form of externalizing behavior because it often involves individuals rebelling against the authority of their parents, who often encourage their children to be involved in their religious tradition; and/or the authority of the church. In communities characterized by high levels of religiosity (e.g., Mormon communities in Utah), religion may constitute rebellion against the social and political leadership of the community.
Genetic etiology of apostasy as an externalizing behavior
If apostasy is in fact an externalizing behavior, then genetic factors associated with other externalizing behaviors may also influence an individual’s likelihood of being an apostate. Evidence for a genetic etiology of externalizing behaviors, and possibly apostasy, comes from the literature on a phenomenon known as ‘behavioral disinhibition’. Behavioral disinhibition is an umbrella term used to refer to a genetic liability for substance use behaviors and externalizing behaviors (Koenig et al., Reference Koenig, McGue and Iacono2008; Krueger et al., Reference Krueger, Hicks, Patrick, Carlson, Iacono and McGue2002; Young et al., 2000, Reference Young, Friedman, Miyake, Willcutt, Corley, Haberstick and Hewitt2009). Research on behavioral disinhibition began after researchers observed the comorbidity of a variety of externalizing behaviors such as oppositional defiant disorder, conduct disorder, attention-deficit disorder, as well as substance use behaviors. Researchers estimate the heritability of behavioral disinhibition to be approximately 80% (Krueger et al., Reference Krueger, Hicks, Patrick, Carlson, Iacono and McGue2002; Young et al., Reference Young, Stallings, Corley, Krauter and Hewitt2000) — meaning that 80% of the variance of behavioral disinhibition is due to genetic factors.
In addition to the heritability studies, a few specific genetic variants have been found to be associated with substance use and externalizing behaviors. For example, Hendershot and colleagues (Reference Hendershot, Bryan, Feldstein Ewing, Claus and Hutchison2011) found that the cholinergic muscarinic receptor 2 gene (CHRM2), which codes for a muscarinic acetylcholine receptor subtype, is associated with latent measures of substance use and overall behavioral disinhibition (Threlfell et al., Reference Threlfell, Clements, Khodai, Pienaar, Exley, Wess and Cragg2010; Volpicelli & Levey, Reference Volpicelli and Levey2004; Woolf & Butcher, Reference Woolf and Butcher2011). If in fact apostasy is an externalizing behavior, then genetic factors such as CHRM2 may predict an individual’s likelihood of being an apostate, which would be revealed as a significant heritability for one or more measures of apostasy.
Correlates of Apostasy
Studies have identified several factors that are correlated with apostasy. These factors include being male (Zuckerman et al., Reference Zuckerman, Galen and Pasquale2016, pp. 95 and 97), having high levels of educational attainment (Ecklund & Scheitle, Reference Ecklund and Scheitle2007; Mayrl & Oeur, Reference Mayrl and Oeur2009; Zuckerman et al., Reference Zuckerman, Galen and Pasquale2016, pp. 95 and 97), using alcohol and/or marijuana (Hadaway, Reference Hadaway1989; Uecker et al., Reference Uecker, Regnerus and Vaaler2007), being politically liberal (Hadaway, Reference Hadaway1989; Zuckerman et al., Reference Zuckerman, Galen and Pasquale2016, pp. 95 and 97), being single (Hadaway, Reference Hadaway1989), having lower levels of happiness (Zuckerman et al., Reference Zuckerman, Galen and Pasquale2016, pp. 95 and 97), and having parents who are divorced (Lawton & Bures, Reference Lawton and Bures2001).
Genetic etiology of apostasy via the ‘phenotypic bottleneck’
Of those factors correlated with apostasy, several have a genetic etiology. These include using alcohol and/or marijuana (Han et al., Reference Han, McGue and Iacono1999; McGue & Iacono, Reference McGue and Iacono2008; Treutlein & Rietschel, Reference Treutlein and Rietschel2011), having high levels of educational attainment (Rietveld et al., Reference Rietveld, Medland, Derringer, Yang, Esko, Martin and Koellinger2013), being politically liberal (Hatemi et al., Reference Hatemi, Medland, Klemmensen, Oskarsson, Littvay, Dawes and Martin2014), being single (Jerskey et al., Reference Jerskey, Panizzon, Jacobson, Neale, Grant, Schultz and Lyons2010), and having lower levels of happiness (Nes & Røysamb, Reference Nes and Røysamb2017). Genetic factors associated with these traits may influence apostasy through what Freese (Reference Freese2008) describes as ‘the phenotypic bottleneck’. According to Freese, genes influence the social world indirectly through their effects on individual-level characteristics (phenotypes). Therefore, as genetic factors influence individual-level traits such as substance use, they may also indirectly influence a correlated social factor such as levels of apostasy in society.
Materials and Methods
Sample
The data for my analyses come from Wave I and Wave III of the National Longitudinal Study of Adolescent to Adult Health (Add Health), a nationally representative longitudinal survey of adolescents and young adults obtained from an initial in-school survey of middle and high school students conducted from September 1994 to April 1995. In total, 90,118 adolescents who attended 80 high schools and 54 feeder schools (both public and private) took part in the initial interview. During the months of April through December 1995, a sample of the in-school respondents (stratified by gender and grade) were selected to participate in an in-home, face-to-face interview (Wave I). These respondents have been followed up three times over the past 15 years for a total of four waves of in-home data collection.
All the data for these analyses come from the sibling subsample of Add Health. The Add Health study oversampled twin-pairs identified in the in-school survey, and this sample design enables these analyses. Respondents who reported during Wave I that they had a full sibling or a twin were included in the pairs roster; and of the 3139 pairs who were asked, 83% (n = 2612) agreed to take part in the study. Any respondents who were singletons (i.e., had no sibling) were dropped from my analyses (n = 19,263). To avoid the potential of biased estimates of heritability due to racial confounding, non-White cases were dropped (n = 687). I chose to focus on White respondents alone because religion, and by extension apostasy, may function very differently among different racial/ethnic groups. For example, studies show that religion plays a much more central role in the lives of African Americans in the USA than it does for Whites (Ellison & Sherkat, Reference Ellison and Sherkat1999; Hunt & Hunt, Reference Hunt and Hunt1999; Pattillo-McCoy, Reference Pattillo-McCoy1998). Therefore, the factors that lead to apostasy among African Americans may be very different than the factors that lead to apostasy among Whites.
In order to compare individuals who were adolescents during Wave I and early adults who had transitioned out of their parental homes by Wave III, cases were also dropped if the respondent was older than 18 years during Wave I (n = 29) and had not transitioned out of their family home by Wave III (n = 165). Lastly, cases were also dropped if they had missing values for apostasy during either Wave I or Wave III (n = 100). As Table 1 indicates, this leaves a total analytic sample that includes 337 twin-pairs. This sample includes 127 monozygotic (identical) twin-pairs and 210 dizygotic (fraternal) twin-pairs. Table 1 reveals that the full analytic sample is 67% male and has ages ranging from 12 to 18 during Wave I and 18 to 25 during Wave III.
Table 1. Descriptive statistics for age, sex and apostasy with intrapair correlations

Note: N = number of nonmissing cases in full sample, MZ = monozygotic (identical) twin-pairs, DZ = dizygotic (fraternal) twin-pairs. *Intrapair correlations are tetrachoric correlations.
Measures
Apostasy
Because religion is a multidimensional concept, three distinct measures of apostasy are used in the analyses: disengagement from religious institutions, cessation of prayer and religious disaffiliation. Each measure of apostasy captures disengagement from a different aspect of religion. While some scholars may only consider ‘true apostates’ to be individuals who are not engaged in any aspect of religiosity after first being engaged in religion, most scholars of apostasy recognize that apostates are a diverse group and vary in the specific way in which they disengage from religion (Zuckerman et al., Reference Zuckerman, Galen and Pasquale2016, pp. 93 and 94). Within my sample, only 20 individuals (≈3% of the sample) would be considered ‘true apostates’, meaning that they disengaged from religious institutions, ceased to pray and disaffiliated from religion altogether. This group only included four complete twin-pairs, meaning that an analysis of only this group would not be feasible with these data.
Disengagement from religious institutions
Disengagement from religious institutions measures disengagement from religious behaviors — specifically, disengagement from attending meetings of religious individuals. Disengagement from religious institutions is a measure based on two indicators: attendance at religious services and participation in religious activities. Respondents were asked how often they attended religious services in the past 12 months and how often they attended ‘special activities outside of regular worship services — such as classes, retreats, small groups, or choir’ in the past 12 months. Responses for both indicators range from never to more than once a week. Individuals were considered to be disengaged from religious institutions if they attended religious services and/or activities during Wave I and did not attend religious services or religious activities during Wave III. According to Table 1, 14% of respondents have disengaged from religious institutions between Wave I and Wave III.
Religious disaffiliation
Religious disaffiliation measures disengagement from a religious identity. Individuals who disaffiliate no longer see themselves as being part of a religious denomination (e.g., Baptist, Catholic). This is fundamentally different to disengagement from religious institutions because an individual can still consider him or herself part of a religious denomination and not attend services. Alternatively, someone can consider him or herself no longer part of a religious denomination while continuing to attend religious services. Evidence for this comes from Ecklund and Lee (Reference Ecklund and Lee2011), who found that atheist parents attend religious services at a higher rate compared to atheist nonparents. Religious disaffiliation is a measure based on whether or not a respondent indicated being part of a religious denomination or not. A respondent was considered disaffiliated if he or she indicated that they were part of a religious denomination in Wave I and indicated that they were not part of any religious denomination in Wave III. According to Table 1, 12% of respondents have disaffiliated from religion between Wave I and Wave III.
Cessation of prayer
Because there is no variable within Add Health that specifically asks whether or not a respondent believes in God or a higher power, cessation of prayer measures disengagement from religious beliefs. The logic behind this decision is that prayer assumes a belief in a higher power, and therefore disengagement from prayer measures a change in religious beliefs. Each measure of apostasy is dichotomous with a score of 1, indicating that a respondent is an apostate for that particular aspect of religion, and 0, indicating whether the respondent is not an apostate in that particular aspect of religion. Cessation of prayer is a measure based on an indicator of frequency of prayer. Respondents were asked how often they prayed, and responses ranged from never to daily. Individuals were considered to have ceased from praying if they prayed at least once a month during Wave I and never prayed during Wave III. According to Table 1, 12% of respondents have ceased praying between Wave I and Wave III.
Model Fitting
For each measure of apostasy, four structural models are fit: a full ACE model that decomposes observed individual-level variance into latent genetic (A), shared environmental (C) and unshared environment/error components (E); a reduced model that drops the shared environmental component; a reduced model that drops the genetic component; and a reduced model that drops both the genetic and shared environmental components.
Each model is estimated by maximum likelihood methods using OpenMx. OpenMx provides estimates for each model parameter by numerical search for the parameter values that minimize a function that is twice the difference between the likelihood of the data under the model to be tested and the likelihood for the perfectly fitting model. Each nested submodel (i.e., AE, CE and E) is compared to the full ACE model utilizing the −2 log likelihood (−2LL), chi-square p value and the Akaike Information Criterion (AIC). For each of the measures of apostasy the model is considered to have the best fit if (1) the difference in the −2LL between two models is nonsignificant as indicated by the chi-square p value; and (2) it has the smallest AIC. The AIC is calculated using the following equation: AIC = 2k − 2ln (L), where k is the number of parameters in the model and L is the maximum likelihood estimation (MLE) for the model. While the AIC has several benefits, including its ability to compare nonnested models, it has several drawbacks that include not having a distribution. This means that difference tests cannot be used to compare AICs. Also, standard AICs are not applicable for a small sample, so an adjustment for small samples (see Burnham & Anderson, Reference Burnham and Anderson2002 and Hurvich & Tsai, Reference Hurvich and Tsai1989, for an explanation of the AICs for small samples) is necessary.
Results
Table 1 displays the descriptive statistics for each of the apostasy measures for the full sample and by twin type. While the means of the ages and percentages for sex and apostasy appear to be fairly consistent across groups, differences appear between the intrapair correlations for all three measures of apostasy. Specifically, the intraclass correlations appear to be larger for MZ twin-pairs compared to DZ twin-pairs. In the case of cessation of prayer (rMZ = .41; rDZ = .08) and religious affiliation (rMZ = .79; rDZ = .31), the intrapair correlations are much larger for MZ twin-pairs compared to DZ twin-pairs. These findings would indicate that genetic factors likely influence apostasy due to the fact that differences in the correlations on a given trait between MZ twin-pairs and DZ twin-pairs, assuming that environments are equal, are likely due to the increased number of shared genes between MZ twin-pairs compared to DZ twin-pairs.
Table 2 presents the goodness-of-fit criterion along with the estimated parameters of the ACE model and submodels for each of the measures of apostasy. Table 2 reveals that for disengagement from religious institutions there is a small difference (Δ = 0.62) between the AIC of the AE model and the CE model. Therefore, there is not enough power to differentiate between A and C, and thus the best explanation of the results is familial aggregation (CE). This finding is more in line with the general perception of apostasy, and religiosity as a whole, which is that it is solely the consequence of social and psychological factors. However, Table 2 also reveals that the reduced model that drops shared environment (AE) is the best fitting model for religious disaffiliation and cessation of prayer. In accordance with the intrapair correlations from Table 1, this finding reveals that genetic factors likely influence religious disaffiliation and cessation of prayer. Specifically, genetic factors account for approximately 75% of the variance in religious disaffiliation and approximately 34% of the variance in cessation of prayer. These results support the idea that genetic factors associated with risky behaviors, educational attainment or political ideology may also influence whether or not someone disengages from religion.
Table 2. Model fit statistics for ACE models and variance components for measures of apostasy
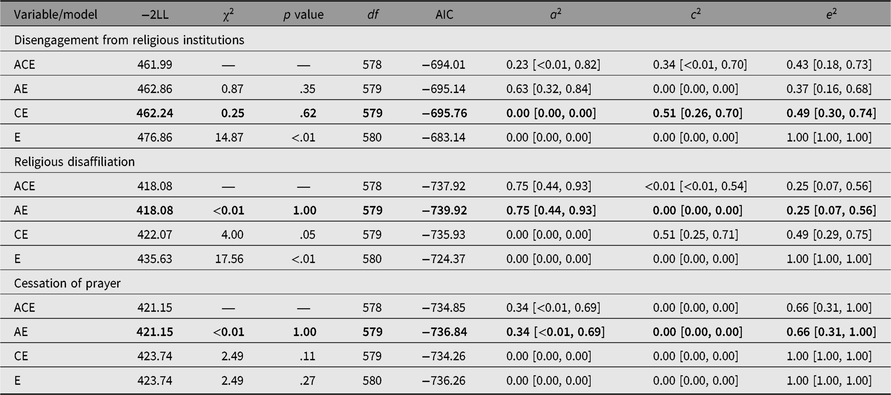
Note: Insignificant p values (p > .05) and low AIC (relative to other models) indicate acceptable model fit for nested models. Cell entries are: full model (containing genetic, shared environmental and nonshared environmental factors)/reduced model (shared environmental factor dropped)/reduced model (genetic factor dropped)/reduced model (genetic factor and shared environmental factor dropped). 95% confidence intervals are shown in square brackets. The best fitting models are shown in bold.
Lastly, because of the potential of bias in the submodels of the ACE model (Edwin et al., Reference Edwin, Oord, Simonoff, Eaves, Pickles, Silberg and Maes2000), 95% confidence intervals are included for each model parameter estimate. While the confidence intervals show a wide range for each parameter estimate, they also reveal that each of the model parameters is significantly different to zero. This further supports the idea that genetic factors influence whether or not individuals disengage from religion between adolescence and early adulthood.
Discussion
The present study explores whether genetic factors explain variation in the levels of apostasy — defined as a disengagement from religious belief, identity and/or practice — in a US-based sample during the transition from adolescence to early adulthood. My main hypothesis is that genetic factors at least partially explain the variance of all three measures of apostasy during the transition between adolescence and early adulthood. Results lend support to my hypothesis for two measures of apostasy in my study: disaffiliation from religious institutions and cessation of prayer. Genetic factors account for approximately 75% of the variance in religious disaffiliation and 34% of the variance in cession of prayer, though the 95% confidence intervals suggest a wide range of potential estimates for the contribution of genetic factors to the variance of these measures of apostasy.
There are at least three potential pathways that may link genetic factors with disaffiliation from religion and cessation of prayer during the transition from adolescence to early adulthood. First, during puberty, genes may cause neurological changes that increase an individual’s likelihood for disaffiliating from religion or ceasing to pray. Studies have shown that during puberty genetic factors lead to neurological changes that increase impulsivity in individual. Steinberg (Reference Steinberg2008) postulates that impulsive behaviors begin to increase during adolescence because the dopaminergic system is ‘reconfigured’ during this period. In particular, the density of receptors in the nucleus accumbens region of the prefrontal cortex are reduced, while the density of myelinated axons (white matter) leading into the prefrontal cortex increases, which leads to increased levels of circulating dopamine in the prefrontal cortex. According to Steinberg, these factors lead to increased rewards from risky behaviors, which lead to an increased prevalence of these behaviors. Declaring oneself free of a denomination or deciding to cease from daily prayers may be thrilling for a risk-seeking person, while they would be daunting to a risk-averse person.
Second, genetic factors related to externalizing behaviors may also lead individuals to become apostates. Ceasing to pray or disaffiliating from religion may be seen by religious adolescents as a form of resistance to parents or other religious authorities. Therefore, religious adolescents with a predisposition toward externalizing behaviors may be more likely to cease prayer or disaffiliate from religion.
Lastly, genetic factors may influence apostasy indirectly via the ‘phenotypic bottleneck’. Genetic factors may directly (or indirectly) influence an individual’s level of educational attainment or substance use, and/or whether or not the respondent is politically liberal. In turn, the traits may also influence an individual’s likelihood of disaffiliating from religion or ceasing to pray. Therefore, genetic factors associated with substance use, political ideology or education attainment would also partially account for variance in cessation of prayer and religious disaffiliation.
I suspect that the lack of a significant genetic component of the variance of disengagement from religious institutions may be explained by the fact that disengaging from religious institutions is a fundamentally different process than disaffiliating from religion or ceasing to pray. Failure to attend church may be the consequence of factors that do not impact the strength of one’s religiosity, such as a changing work schedule or moving to a new area. In other words, an individual may not attend religious services, but not be less religious than they were when they did attend religious services regularly. In addition, individuals may replace attending religious services with virtual churches such as televangelists or religious services available online (Emerson et al., Reference Emerson, Mirola and Monahan2011, pp. 41–45). Ceasing to pray and/or disaffiliating from religion seems to be a more definitive break from religion, which reflects either a lessening of religious belief (cessation of prayer) or a fundamental rejection of religious bodies (religious disaffiliation).
Overall, I feel this study significantly pushes forward our knowledge regarding the etiology of apostasy in the USA, though there are some limitations. First, these findings are limited to individuals in the 18–25-year-old range. While apostasy can occur throughout the life course, the potential causes of genetic influence on apostasy described in this article may not be relevant to apostasy that occurs earlier or later in the life course. Second, these findings are limited to White respondents in the USA. While this was done for reasons laid out previously in the paper, it does limit the scope of this particular paper. Lastly, underlying assumptions of twin models often call their validity into question. The violation of one assumption in particular could significantly bias these results. The equal environments assumption, which is the assumption that the environments of MZ twins are no more or less similar than the environments of DZ twins, allows us to identify our models by holding the correlation of the shared environments of MZ twins and DZ twins to 1.0. A violation of this assumption would not only make it impossible for us to identify our models, it would also call into question the validity of the twin method itself. However, studies have tested this assumption using misidentified twins (e.g., DZ twins identified as MZ twins), and from these studies there is little evidence that the assumption is violated in most analyses or that any violations biased results (Evans & Martin, Reference Evans and Martin2000).
Future research should explore which specific genetic variants (e.g., single nucleotide polymorphisms) influence apostasy, as well as the specific biological mechanisms linking genetic variants, and whether or not an individual chooses to disengage from religion. Future research should also explore how specific aspects of the shared family environment (e.g., parental religiosity) influence religiosity. However, this is beyond the scope of this particular article, which is specifically deciphering whether or not apostasy is heritable.
Acknowledgments
I would like to acknowledge the help of Dr. J. Micah Roos who provided advice to improve this paper prior to submission; and my wife, Alison Hart Freeman, who helped me revise this paper for resubmission.
Conflict of interest
None.


