


Introduction
Seed dormancy affects both germination rate and percentage (Batlla and Benech-Arnold, Reference Batlla and Benech-Arnold2007; Bochenek et al., Reference Bochenek, Golaszewski and Gorecki2007; Batlla and Benech-Arnold, Reference Batlla and Benech-Arnold2010). Defined as the condition of a seed that inhibits germination under otherwise sufficient environmental conditions, dormancy can be classified as primary (i.e. present in seeds at maturity) or secondary (i.e. induced in previously non-dormant seeds or re-induced in seeds that have lost primary dormancy) (Benech-Arnold et al., Reference Benech-Arnold, Sánchez, Forcella, Kruk and Ghersa2000). Dormancy is not an ‘all or nothing’ seed characteristic, but varies over a continuum between some maximum and minimum level (Batlla et al., Reference Batlla, Kruk, Benech-Arnold, Benech-Arnold and Sánchez2004). Dormancy levels can vary among seeds within a population (Bradford, Reference Bradford2002; Batlla et al., Reference Batlla, Kruk, Benech-Arnold, Benech-Arnold and Sánchez2004), as well as seasonally throughout the year (Finch-Savage and Leubner-Metzger, Reference Finch-Savage and Leubner-Metzger2006).
The distinction between dormancy release and germination stimulation by environmental cues is a current topic of discussion in seed biology (e.g. Finch-Savage and Leubner-Metzger, Reference Finch-Savage and Leubner-Metzger2006; Finch-Savage and Footitt, Reference Finch-Savage and Footitt2012; Battla and Benech-Arnold, Reference Batlla and Benech-Arnold2015). Hydrothermal time models that deal with dormancy release and germination tend to support the idea that these two processes are not discrete but can be proceeding simultaneously in a seed population and possibly even in individual seeds. For seeds of winter annuals such as Bromus tectorum, which lose primary dormancy under dry conditions but accumulate progress towards germination only when at least partially imbibed, these two processes are largely uncoupled (Meyer and Allen, Reference Meyer and Allen2009). Secondary dormancy induction in a winter annual, on the other hand, is likely to take place in seeds that are at least partially imbibed, creating the possibility of accumulation of progress towards germination and secondary dormancy induction as opposing processes occurring simultaneously in individual seeds.
A hydrothermal time model is a population-based threshold model that describes time-normalized progress towards germination as a function of temperature and water potential. The use of hydrothermal time to describe seed germination was introduced by Gummerson (Reference Gummerson1986) and expanded by Bradford (Reference Bradford1990, Reference Bradford, Kigel and Galili1995). Key concepts of this modelling approach include a base or threshold temperature and water potential below which seeds do not germinate. Thus the time to germination for a specific fraction of a seed population can be calculated using the following equation:
 $${\rm \theta} _{{\rm HT}} = \left( {\Psi - \Psi _{\rm b} \left( g \right)} \right) \left( {T-T_{\rm b}} \right)t_g,$$
$${\rm \theta} _{{\rm HT}} = \left( {\Psi - \Psi _{\rm b} \left( g \right)} \right) \left( {T-T_{\rm b}} \right)t_g,$$
where θHT is the hydrothermal time requirement for the germination (hydrothermal time constant), Ψ and T are the actual water potential and temperature of the environment, Ψb is the base water potential for germination fraction g, T b is the base temperature for hydrothermal time to accumulate, and t is the actual time to germination for fraction g of the given population.
To extend this model to account for an entire population, Gummerson (Reference Gummerson1986) assumed that the distribution of base water potentials, the variable factor in a population, is approximately normal with mean base water potential (Ψb(50)) and standard deviation (σΨb). This assumption allowed him to use probit transformation, which linearizes the cumulative normal distribution associated with germination time courses, to characterize germination for the entire seed population using the following equation:
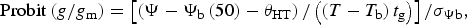 $${\rm Probit} \left( {g/g_{\rm m}} \right) = \left[ {\left( {\Psi - \Psi _{\rm b} \left( {50} \right) - \theta _{{\rm HT}}} \right)/\left( {\left( {T-T_{\rm b}} \right)t_{\rm g}} \right)} \right]/\sigma _{\Psi {\rm b}},$$
$${\rm Probit} \left( {g/g_{\rm m}} \right) = \left[ {\left( {\Psi - \Psi _{\rm b} \left( {50} \right) - \theta _{{\rm HT}}} \right)/\left( {\left( {T-T_{\rm b}} \right)t_{\rm g}} \right)} \right]/\sigma _{\Psi {\rm b}},$$
where g/g m is the fraction of viable seeds in the given population, Ψb(50) is the population mean base water potential, and σΨb is the standard deviation of base water potentials.
The mean base water potential Ψb(50) has been shown to be a valuable index for characterizing seed populations with both ecological (Allen et al., Reference Allen, Benech-Arnold, Batlla, Bradford, Bradfod and Nonogaki2006; Köchy and Tielbörger, Reference Köchy and Tielbörger2007) and physiological (Bradford, Reference Bradford1986; Groot and Karssen, Reference Groot and Karssen1992) relevance. Changes in Ψb(50) have also been used to describe primary dormancy release through dry after-ripening (Christensen et al., Reference Christensen, Meyer and Allen1996; Bauer et al. Reference Bauer, Meyer and Allen1998; Bair et al., Reference Bair, Meyer and Allen2006). A combined hydrothermal time model that included summer dormancy loss through dry after-ripening and accumulation of hydrothermal time in imbibed seeds in the autumn successfully simulated these processes in the field for B. tectorum (Meyer and Allen, Reference Meyer and Allen2009). Here we aim to extend these models to account for the induction and release of secondary dormancy in B. tectorum seeds.
Seeds of many species experience secondary dormancy induction under conditions that are outside the range of environmental conditions that permit rapid germination. Young and Evans (Reference Young and Evans1975) demonstrated that seeds of B. tectorum were more likely to form a persistent seed bank in the field when dispersed into openings rather than under shrub canopies, and they surmised that harsher, drier conditions during winter were probably responsible for this difference. In a field retrieval experiment designed to indirectly test the effect of microclimate on secondary dormancy induction and seed bank persistence in this species, Allen et al. (Reference Allen, Meyer, Foote, Pendleton, Meyer and Schultz2010) found that seeds that were planted into mineral soil or placed beneath litter in September germinated during autumn storms, whereas seeds placed on the surface of the litter in September or planted in any position in early November were much more likely to enter secondary dormancy and persist until the following autumn. The study reported here was carried out with the objective of quantifying both the cues that induce secondary dormancy and the process of secondary dormancy induction and release. We hypothesized that: (1) secondary dormancy induction in B. tectorum would take place gradually at cool temperatures under moderate water stress, while subsequent dormancy release in summer would parallel the dry after-ripening process for primary dormancy release; (2) the propensity to enter secondary dormancy would vary among B. tectorum genotypes and would be related to habitat of origin; and (3) the process of secondary dormancy induction and release could be modelled using hydrothermal time, with changes in Ψb(50) explaining most of the observed patterns of change in dormancy status within a seed population.
Materials and methods
General experimental protocols
All florets (hereafter seeds) used in experiments were cleaned by hand after collection and allowed to after-ripen under ambient laboratory conditions (approximately 23°C and 20% relative humidity), rendering them non-dormant at the onset of each experiment.
Laboratory experimental datasets and the dataset from ‘Field experiment one’ were analysed as fully randomized factorial designs using generalized linear models for binomial response variables (PROC GLIMMIX; SAS 9.4, 2012; SAS Inc., Cary, NC, USA). Means separations were performed as appropriate using least squares means testing.
Laboratory experiments
Secondary dormancy induction at 5°C
Seeds from Bromus tectorum were collected in June 2011 from a wild population at the Brigham Young University research farm (Spanish Fork, UT, USA). Seeds were incubated (5°C, continual darkness) for 28 days at one of five water potentials (0, –0.5, –1.0, –1.5 or –2.0 MPa) obtained using solutions of polyethylene glycol 8000 (Michel and Kaufmann, Reference Michel and Kaufmann1972; Michel, Reference Michel1983). Solution water potentials were verified with a water activity meter (Dewpoint PotentiaMeter WP4, Decagon Devices, Pullman, WA, USA). For each treatment, four replications of 25 seeds each were placed in covered 100 mm Petri dishes on the surface of two blue germination blotters (Anchor Paper, St Paul, MN, USA) saturated with the appropriate solution. In order to minimize any changes in water potential due to drying, Petri dishes were stacked in sealed plastic sleeves on top of a water-saturated paper towel before being placed in an incubator. Germination (radicle emergence ≥ 1 mm) was recorded weekly. After 7,14 or 28 days, seeds in negative water potentials were transferred to water and all seeds in water were incubated at 20°C, (alternating 12 h fluorescent light/12 h dark) for an additional 28 days. Seeds were then scored for germination on days 1, 2, 4, 7, 11, 14, 21 and 28. Remaining seeds were considered viable and dormant on day 28 if firm when pressed, which we have found comparable to a cut test (Ooi et al., Reference Ooi, Aulk and Whelan2004) for B. tectorum seeds.
Secondary dormancy induction at multiple chilling temperatures
To determine if temperatures lower than 5°C would improve secondary dormancy induction, seeds from four genotypes were subjected to a factorial combination of low temperatures and water potentials. Seeds were collected from greenhouse-grown plants of four contrasting genotypes in summer 2010. These genotypes included Salt Desert 1 (Dugway Proving Grounds, UT), Salt Desert 2 (Hot Springs Mountain, NV), and two core (cold desert generalist) genotypes [Desert Experimental Range, UT; Cricket Mountains, UT; see Merrill et al. (Reference Merrill, Meyer and Coleman2012) and Meyer et al. (Reference Meyer, Leger, Eldon and Coleman2016) for details on genotype classification and collection location]. All seed lots were non-dormant and had germination >95% at the onset of the experiment.
Seeds of the four genotypes were incubated in each of five water potentials (0, –0.5, –1.0, –1.5, –2.0) at one of three low temperatures (0, 2, 5°C). For each treatment, four replications of 25 seeds were placed in covered 100 mm Petri dishes and incubated for 28 days. After 28 days, seeds in negative water potentials were transferred to water and all seeds in water were incubated at 20°C (alternating 12 h fluorescent light/12 h dark) for an additional 28 days. Germination was recorded as previously described.
Field experiment one
Bromus tectorum seeds belonging to ten inbreeding lines were produced by greenhouse cultivation in 2007 and allowed to after-ripen under laboratory conditions. These included representatives of habitat specialist genotypes Warm Desert 1 (FEDD), Warm Desert 2 (EBBF), Salt Desert 1 (IEBB), Montane 1 (GCCB) and Montane 2 (DABB), as well as five representative core generalist genotypes [see Merrill et al. (Reference Merrill, Meyer and Coleman2012) and Meyer et al. (Reference Meyer, Leger, Eldon and Coleman2016) for details on B. tectorum genotype classification and collection location].
Seeds of each line were randomly divided into lots of approximately 50 and inserted into nylon mesh bags. These bags were placed in the field on 16 September 2008, on the surface of the litter at the Whiterocks study site (40° 19.680′ N, 112° 46.680′ W, elevation 1446 m, average annual precipitation 200 mm). This placement was based on results of an earlier experiment showing that seeds placed on the surface of the litter did not accumulate sufficient hydrothermal time to germinate (Allen et al. Reference Allen, Meyer, Foote, Pendleton, Meyer and Schultz2010). On 20 October, 17 November, 15 December, 23 February and 8 May, eight bags of each line were retrieved at random. No seeds germinated or developed Pyrenophora semeniperda disease signs in the field. Retrieved seeds were placed in incubation at 10/20°C or 20/30°C with a 12-h photoperiod (cool white fluorescent light). Each incubation temperature treatment included four replicate Petri dishes of approximately 50 seeds per line. Germination was scored at 1, 2, 4, 7, 11, 14, 21 and 28 days. Remaining ungerminated seeds were scored as viable (firm, as described above), non-viable, or killed by P. semeniperda (with black stromata protruding from ungerminated seeds; for details see Finch-Boekweg et al., Reference Finch-Boekweg, Gardner, Allen and Geary2016). There were no pathogen-killed seeds in the first three retrievals, while the February retrieval showed an average of 11% mortality and the May retrieval showed an average of 16%. This pathogen attacks dormant seeds (Beckstead et al. Reference Beckstead, Meyer, Molder and Smith2007). We assumed that pathogen-killed seeds were killed after entering secondary dormancy and therefore used initially viable seed number as the basis for expressing the germinable fraction.
Field experiment two
Seeds from two populations of B. tectorum were collected in June 2011: one from the Brigham Young University research farm (Spanish Fork, UT) and the other from Tooele County (Whiterocks, UT). Seeds from each population were divided and placed into 40 nylon mesh bags (ca 300 seeds/bag). On 9 November 2012 at the Whiterocks study site, bags were placed on the soil surface after removal of the top 2 cm of soil and covered with 2 cm of autoclaved B. tectorum litter. This procedure was followed in an effort to limit P. semeniperda infection. Bags and litter were held in place by coarse nylon mesh secured with nails.
A digital data logger (Em50, Decagon Devices) was installed at the site immediately adjacent to the field plot. Hourly values for soil temperature and water content in the seed zone were recorded using a 5TM Soil Moisture and Temperature Sensor (Decagon Devices). Soil water content readings were converted to water potential values through a calibration curve as described by Meyer and Allen (Reference Meyer and Allen2009).
Subsamples (two random bags from each population) were retrieved and returned to the laboratory at monthly intervals beginning on 10 December 2012 and ending on 8 August 2013. Retrieved seeds were randomly assigned to one of three laboratory incubation water potentials (0, –0.5 or –1.5 MPa) and one of two incubation temperatures (15 or 25°C), thus providing data for the calculation of hydrothermal time equations after each retrieval. Seeds that had germinated or had developed P. semeniperda stromata were counted and excluded from further analysis. For each treatment, four replications (an equal number of the remaining seeds, generally at least 20) of each seed population were placed in covered Petri dishes on the surface of two blue germination blotters (Anchor Paper) saturated with water or the appropriate polyethylene glycol (PEG) solution. After 28 days, seeds at low water potential were transferred to water and incubated at the same temperatures (15 or 25°C) for an additional 28 days. Germination was recorded on days 1, 2, 4, 7, 11, 14, 21 and 28, and remaining ungerminated seeds were scored as dormant, non-viable, or killed by P. semeniperda. For this experiment, germination fraction was expressed in terms of currently viable seeds rather than initially viable seeds, i.e. field-germinated and pathogen-killed seeds were not included. Overall, approximately 11% of seeds germinated in the field and 8% suffered pathogen mortality that was evident either in the field or after incubation.
Hydrothermal time equation development
Laboratory data were used to estimate θHT, σΨb and Ψb(50) for fully after-ripened seeds of each seed population. These models assumed that T base was 0°C as demonstrated earlier (Christensen et al. Reference Christensen, Meyer and Allen1996). A probit regression model was created for each seed population by collapsing germination curves obtained from the ten factorial incubation treatments (two temperatures × five water potentials) into a single regression. This resulted in estimates of hydrothermal time parameters for each population (Bradford, Reference Bradford, Kigel and Galili1995; Table 1 herein, initial values).
Table 1. Changes in hydrothermal time parameters for two Bromus tectorum seed collections when all model parameters were allowed to vary during secondary dormancy induction and release in the field (modelling approach 2)

Retrievals from missing months (January–June) had insufficient germination for parameter estimation because of very high dormancy levels. Units for θHT are MPa degree days, and for σΨb and Ψb(50) are MPa.
Modelling technique one
We used two different approaches to characterize acquisition and loss of secondary dormancy in the field using hydrothermal concepts. The first model assumed that θHT and σΨb remained constant across field retrievals, while Ψb(50) was allowed to fluctuate as an indication of dormancy status. These assumptions were previously validated for primary dormancy loss under dry conditions in B. tectorum (Christensen et al., Reference Christensen, Meyer and Allen1996; Bair et al., Reference Bair, Meyer and Allen2006). When germination was >50% for field-retrieved seeds imbibed in water, time to 50% germination was determined directly from germination time courses. In order to calculate Ψb(50) the following relationship was used:
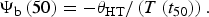 $$\Psi _{\rm b} \left( {50} \right) = -{\rm \theta}_{{\rm HT}} /\left( {T\left( {t_{50}} \right)} \right).$$
$$\Psi _{\rm b} \left( {50} \right) = -{\rm \theta}_{{\rm HT}} /\left( {T\left( {t_{50}} \right)} \right).$$
Values for t 50 were determined using linear interpolation between the two points surrounding the 50% fraction. When final germination did not reach 50% of viable seeds, but was between 5 and 50%, we used Eqn (3) to calculate Ψb(g) from the t g for a known germination fraction g, then used σΨb to estimate Ψb(50) with the following equation (Bauer et al. Reference Bauer, Meyer and Allen1998):
 $$\Psi _{\rm b} \left( {50} \right) = \Psi _{\rm b} \left( g \right) - \left( {{\rm \sigma}_{\Psi {\rm b}}} \right)\,{\rm probit}\,\left( g \right).$$
$$\Psi _{\rm b} \left( {50} \right) = \Psi _{\rm b} \left( g \right) - \left( {{\rm \sigma}_{\Psi {\rm b}}} \right)\,{\rm probit}\,\left( g \right).$$
Modelling technique two
As an alternative to allowing only Ψb(50) to vary across retrievals, we used the same dataset but recalculated all parameters [θHT, σΨb and Ψb(50)] following laboratory incubation at multiple temperatures and water potentials after each seed retrieval. We included this method because the competing processes of hydrothermal time accumulation and secondary dormancy induction can potentially occur simultaneously in even partially imbibed seeds. Thus in our second modelling approach we hypothesized that θHT would decrease over time as seeds progressed towards germination, while Ψb(50) would increase as secondary dormancy was induced.
Germination curve predictions
To evaluate the success of each model, we compared predictions of germination time courses from each of the hydrothermal models with observed germination time courses for laboratory-germinated seeds in water (0 MPa) at two incubation temperatures following retrieval from the field. We used the probit equation [Eqn (2)] to create predicted time courses using the hydrothermal time parameters from each model and then transformed the probit values to the corresponding germination fractions.
Results
Laboratory experiments
Secondary dormancy induction at 5°C
Seeds incubated at 20°C across all water potentials showed no dormancy following transfer to water (Fig. 1A). Similarly, seeds incubated for 7 or 14 days at 5°C at these same water potentials failed to become dormant (Fig. 1B, C). However, seeds incubated at 5°C and sub-zero water potentials for 28 days had decreased total germination. These seeds remained firm and free from fungal contamination, which indicated that they were viable but had been induced into secondary dormancy (Fig. 1D). Dormancy levels after the 28-day chilling treatment varied by Ψ; maximum dormancy induction occurred at –1.0 MPa. Seeds incubated in water at 0 MPa and 5°C failed to enter secondary dormancy, as indicated by complete germination (Fig. 1D). Thus it was clearly the interaction between Ψ and chilling duration that resulted in secondary dormancy induction (chilling duration × water potential interaction: d.f. = 19, 60; F = 8.50, P < 0.0001).

Figure 1. Results of a secondary dormancy induction experiment with fully after-ripened Bromus tectorum seeds (Spanish Fork Farm, UT) incubated at 20°C in water for 28 days (A) or at 5°C for either 7 (B), 14 (C) or 28 days (D) at several water potentials (0, –0.5, –1.0, –1.5 and –2.0 MPa). Seeds at all negative water potentials were transferred to water and incubated for an additional 28 days at 20°C (transfer represented by the vertical dashed line on each graph). Final germination means separations for the chilling duration by water potential interaction are from generalized mixed model analysis of variance for a binomial variable (PROC GLIMMIX, SAS 9.4). Final germination values followed by the same letter do not differ significantly at the P < 0.05 level.
Secondary dormancy induction at multiple chilling temperatures
When seeds of four B. tectorum genotypes were chilled at 0°C across a range of water potentials, few or none became dormant (Fig. 2, white bars), and the only substantial dormancy induction observed was during chilling at 5°C at sub-zero water potentials. Genotypes of B. tectorum varied significantly in response to the dormancy induction treatments (genotype × chilling temperature × water potential interaction: d. f. = 59,180; F = 8.76; P < 0.0001). Seeds of the core Cricket Mountain line were not rendered secondarily dormant under any condition in the laboratory, while seeds of the core Desert Experimental Range line experienced slight but significant dormancy induction only in the 5°C chilling treatment at –1.5 MPa. Seeds of the Salt Desert 1 line were somewhat induced into secondary dormancy at –1.5 MPa in the 2°C chilling treatment, but the induction effect was significantly more pronounced in the 5°C treatment. Seeds of the Salt Desert 2 line were induced into dormancy only at 5°C, but significant dormancy induction was observed at water potentials from –0.5 to –1.5 MPa. For this genotype, –1.0 MPa was the optimum water potential for dormancy induction. Overall, this experiment showed that chilling temperatures <5°C were much less conducive to dormancy induction than 5°C, and also that chilling in free water (0 MPa) at any temperature did not induce dormancy. Strong differences between genotypes in induction response were also evident.

Figure 2. Results of a secondary dormancy induction experiment with fully after-ripened greenhouse-grown seeds of four Bromus tectorum genotypes showing final germination after seeds were incubated at 0, 2 and 5°C for 28 days at water potentials from 0 to –2.0 MPa, then transferred to water at 20°C for an additional 28 days. Error bars represent standard error of the mean. See text for statistics.
Field experiment one
When B. tectorum seeds of contrasting genotypes were placed in a field retrieval experiment on the surface of litter in mid-September, seeds of all ten lines remained non-dormant at both post-retrieval incubation temperatures through mid-December, and there were no significant differences among lines during this period (Fig. 3). Secondary dormancy was induced in most lines between mid-December and late February, and germination percentage also decreased significantly later in the winter (as measured in the May retrieval) for six of the ten lines. Only the Jensen, Greensprings, Dry Wash and Diamond Fork lines reached maximum dormancy by late February.

Figure 3. Results of a field retrieval experiment with greenhouse-grown seeds of ten Bromus tectorum genotypes from contrasting habitats showing changes in post-retrieval seed germination fraction after 28 days in water at two incubation temperatures following 0–8 months on the litter surface at the Whiterocks UT study site. Error bars represent standard error of the mean. See text for statistics.
Seeds of most lines showed significantly higher dormancy with the higher incubation temperature treatment as measured over the last two retrievals; the only exceptions were the core lines from Wallsburg, House Mountains and Tenmile Creek (Fig. 3). All ten lines exhibited some level of dormancy induction in the field, with germination percentages in May averaged across incubation temperatures ranging from 22 to 71%. Germination percentages after the February retrieval ranged from 26% (Greensprings) to 97% (Dutch John) at 10/20°C and from 0% (Greensprings) to 87% (Dutch John) at 20/30°C. In May, germination percentages ranged from 27% (Dry Wash) to 84% (Jensen) at 10/20°C and from 1% (Greensprings) to 59% (House Mountains) at 20/30°C. The habitat specialist genotypes tended to show high dormancy induction at the higher incubation temperature whether from desert or montane environments, but Montane 2 (Dutch John) was a clear exception to this trend, and some core genotypes (Dry Wash, Diamond Fork) also tended to display this pattern. Overall there was no clear relationship between genotype or habitat of origin and dormancy induction response.
Field experiment two
Field seed bed environmental conditions
The field experiment was installed into relatively dry soil shortly before a cold storm and the onset of early winter conditions (Fig. 4). After this storm, estimated soil water potentials ranged between –1.0 and –2.0 MPa during nearly all of the first 6 weeks in the field. Within 3 days following installation, soil temperatures dropped to 5°C and fluctuated diurnally above and below this temperature for the first 5 weeks. Thus the variable field soil conditions during the first 6 weeks approximated those that induced seeds into secondary dormancy in the laboratory. The soil surface froze in mid-December and by early January we observed snow cover to a depth of >30 cm. These conditions kept the soil frozen at an unsaturated soil moisture condition until mid-March, when the snow melted and saturated the soil for approximately 10 days. Soil gradually dried to <–0.5 MPa by mid-May, and remained relatively dry and warm (20–60°C, which included large diurnal fluctuations) through the summer months.

Figure 4. Seed bed microclimate conditions during and after the period of secondary dormancy induction in Field experiment 2 at the Whiterocks Study site in 2012–2013. The dashed horizontal line represents 5°C; the continuous line represents 0°C. Arrow indicates date of retrieval installation. Hourly seedbed temperature data were recorded in situ, while daily precipitation data for the Whiterocks study site were downloaded from the PRISM climate interpolator: http://www.prism.oregonstate.edu/explorer/
Dormancy induction in the field
Seeds were 100% germinable when placed in the field in November. They became increasingly dormant through early winter and remained dormant through the winter and spring months (observed germination time courses are shown in Fig. 5). By mid-January, germination percentages in water had dropped to <15%, and by mid-February > 95% of seeds that had not germinated in the field or been killed by P. semeniperda were dormant. A large majority of these seeds remained dormant through June. At this point retrieved seeds increased in total germination percentage under summer conditions until the final retrieval in mid-August, when >85% of remaining viable seeds retrieved from the field germinated when incubated in water.

Figure 5. Predicted (lines) and observed (symbols) time course curves for germination of Spanish Fork (SFF) and Whiterocks (WRK) Bromus tectorum seeds in post-retrieval incubation in water (Ψ = 0) after 1–8 months in the field. Curves were obtained by fitting each of the two hydrothermal time models to the observed laboratory data. Dashed lines represent Modelling approach 1, which allowed only mean base water potential [Ψb(50)] to change (Fig. 4). Continuous lines represent Modelling approach 2, which allowed all hydrothermal parameters [θHT, Ψb(50), σΨb] to vary across retrievals (Table 1). From January to June insufficient germination occurred to estimate Model 1 hydrothermal parameters [θHT, Ψb(50), σΨb], and from February to June germination percentages were too low (0–5% in water) even for estimation of Ψb(50) from the initial hydrothermal time equation.
Comparing modelling approaches
Our first modelling approach required that only Ψb(50) of seeds in free water seeds be re-calculated after each seed retrieval to account for seeds entering and exiting secondary dormancy. Because germination of at least 5% in water was required to calculate hydrothermal time parameters, we were unable to calculate Ψb(50) during winter and spring months. Ψb(50) values for both seed populations were initially low, increased dramatically as seeds were induced into dormancy through the winter, and decreased as seeds lost this dormancy through secondary after-ripening during the following summer (Fig. 6). Whiterocks seeds had an initially lower Ψb(50) value than Spanish Fork and a smaller σΨb. Germination is prohibited when Ψb rises above 0 in hydrothermal models, and a majority of seeds from both populations entered dormancy after 1 month in the field. The threshold value of Ψb(50) above which no seeds will germinate is approximately twice σΨb, shown as a horizontal line for each seed population in Fig. 6. Values of Ψb(50) were generally higher when measured at 25°C compared with 15°C. Both seed populations lost substantial dormancy by July as Ψb(50) decreased to below 0 MPa, and both germination percentage and rate were further increased as Ψb(50) continued to drop through August. Germination time courses predicted using the initial hydrothermal time equations with only changes in Ψb(50) generally fit observed time courses reasonably well except for the December retrieval, when wide divergence was observed in three of four cases (Fig. 5). The seed populations were still somewhat dormant at the time of the August retrieval, i.e. Ψb(50) had not decreased to values similar to those for fully after-ripened seeds, and there was still a Ψb(50) differential between incubation temperatures, at least for the Whiterocks population (Table 1). We assume that dormancy loss would have continued in the field through late summer.

Figure 6. Mean base water potentials calculated from post-retrieval germination time course curves in water at two incubation temperatures, using initial hydrothermal time equations (Table 1) for two Bromus tectorum collections following retrieval from the field over a 9-month period. The dashed line represents the lowest mean base water potential at which any germination in water would be predicted, based on the initial standard deviation of base water potentials for each collection [i.e. the threshold value of Ψb(50) is twice σΨb; see Table 1]. Months with missing values had mean base water potentials above this threshold; i.e. seeds were essentially completely dormant in water.
Our second modelling approach required that all hydrothermal parameters be re-calculated after each seed retrieval. Whiterocks and Spanish Fork seeds displayed similar trends as in Modelling technique 1; specifically, Ψb(50) increased during winter and later decreased through the summer months (Table 1). Values for σΨb remained relatively constant through time for the Whiterocks seed population but showed unexplained wide variation for Spanish Fork. The most notable change through time during dormancy induction and release was in θHT, the hydrothermal time requirement for germination. This parameter reached very low values by the December retrieval, presumably because the process of hydrothermal time accumulation was proceeding simultaneously with the process of dormancy induction. Unfortunately, our modelling approaches did not allow us to directly distinguish between these two processes. There was some field germination in response to the first storm during the first month of the retrieval experiment, so it is likely that even seed fractions that did not germinate experienced gains in hydrothermal time, i.e. progress towards germination. Models that allowed for this downward shift in θHT showed good fit with the experimental data across water potentials (R 2 > 0.92; Table 1). Time course curves in water generated using these December hydrothermal time equations were generally much closer to the observed time courses than the curves fit with the Ψb(50)-change-only model (Fig. 5).
After the December retrieval, germination at sub-zero water potentials was essentially zero, making it impossible to fit specific equations during winter and spring. For specific equations fit to data taken during summer dormancy loss, the best models included an offset in Ψb(50) to account for higher Ψb(50) at the higher incubation temperature; this is similar to the pattern for primary dormancy loss (Christensen et al., Reference Christensen, Meyer and Allen1996; Table 1). θHT increased to values similar to the previous values for fully after-ripened seeds during the summer; apparently, any effect of hydrothermal time accumulation prior to winter was no longer evident. The two alternative modelling approaches predicted germination time course curves equally well in August and for the Whiterocks seed population in July (Fig. 5). For Spanish Fork Farm seeds, the Ψb(50)-change-only model under-predicted at 15°C, while the unique hydrothermal time model under-predicted at 25°C.
In general, the Ψb(50)-change-only models fit the observed data and supported the conclusion that secondary dormancy induction and loss in B. tectorum can be explained largely through changes in this hydrothermal time parameter. In fact, both modelling approaches support this conclusion. Secondary dormancy induction and loss therefore operate in a manner parallel to primary dormancy loss in this species, i.e. primarily through changes in Ψb(50).
Discussion
Under constant laboratory conditions, maximum secondary dormancy induction occurred at water potentials from –0.5 to –1.5 MPa and a temperature of 5°C, confirming our hypothesis that dormancy induction would take place at cool temperature and moderate water stress (Figs 1 and 2). Also as hypothesized, the process was gradual, as induction treatments of less than 4 weeks were not effective. In the second field experiment, temperatures and water potentials in the weeks following a cool late autumn storm fluctuated around 5°C and –1 to –2 MPa, conditions that mimicked the most effective dormancy induction treatments in the laboratory. These conditions rendered a large fraction of the seeds dormant after 1 month, and almost all remaining viable seeds became dormant after 2 months (Fig. 5). Field results therefore confirmed laboratory results: secondary dormancy is induced via the combination of low temperatures and low water potentials under fluctuating conditions as well as constant conditions. In fact, dormancy induction was more complete in the field, suggesting that conditions in the laboratory were not optimal for induction, and that more complex laboratory regimes could induce higher levels of secondary dormancy.
A more surprising result was the fact that 5°C was clearly the optimum temperature for dormancy induction; lower chilling temperatures were not effective (Fig. 2). The relationship between chilling temperature and primary dormancy release has been studied in several species. For example, for Nothofagus species, there was no difference in efficacy for temperatures from 0.5 to 4°C (Arana et al., Reference Arana, Gonzalez-Polo, Martinez-Meier, Gallo, Benech-Arnold, Sánchez and Batlla2016), whereas for the summer annual rice weed Echinochloa oryzicola in the Mediterranean climate of California, 7.5°C was somewhat more effective for releasing primary dormancy than either 2.5 or 5°C (Boddy et al., Reference Boddy, Bradford and Fischer2013). For recalcitrant seeds of Aesculus hippocastaneum, Pritchard et al. (Reference Pritchard, Tompsett and Manger1996) found that stratification efficacy increased linearly below a ceiling temperature of approximately 12°C, i.e. colder stratification temperatures were more effective. The cues that regulate secondary dormancy induction are less well studied, however, especially for fall-germinating species (Hilhorst, Reference Hilhorst1998). For oilseed rape (Brassica napus), secondary dormancy induction takes place under water stress, but the relationship with temperature is not strong (Momoh et al. Reference Momoh, Zhou and Kristiansson2002).
While the physiological mechanism that accounts for an increase in secondary dormancy induction with increasing chilling temperature in B. tectorum is not known, this pattern has a clear ecological interpretation. In the semi-arid winter-rainfall regions of western North America where this weed has become a major invader, autumn storms are generally sporadic and unreliable. A cool autumn storm can trigger germination of seeds in favourable microsites but be likely to induce those in drier, less favourable microsites into dormancy, thereby ensuring both the establishment of a current-year stand and the maintenance of a seed bank for the following year (Allen et al., Reference Allen, Meyer, Foote, Pendleton, Meyer and Schultz2010). If the autumn is very dry, so that the first significant precipitation arrives under cold winter conditions that do not permit germination, it would be advantageous for most of the seeds to remain non-dormant, so that spring emergence remains an option for establishing a current-year stand. Conditions in late winter or early spring could still induce secondary dormancy for seeds in drier microsites, making both spring germination and seed bank carryover possible. Thus it makes sense that the near-freezing temperatures associated with winter storms would not induce secondary dormancy to the same degree as conditions during late autumn or early spring storms.
The predisposition of seeds to enter secondary dormancy was shown to differ among genotypes for B. tectorum (Figs 2 and 3). These differences are likely to be genetic, as seeds used in these experiments were produced in a common environment, reducing the probability of maternal effects. Weber et al. (Reference Weber, Frick, Bruber and Calupen2010) reported large genetically based differences in the propensity of oilseed rape to enter secondary dormancy under osmotic stress, a result similar to ours. The ecological significance of this variation in B. tectorum is not clear, however. There was a trend for genotypes from more xeric habitats to have a greater propensity to enter secondary dormancy under both laboratory and field conditions than genotypes from core cold desert and sagebrush steppe habitats (Figs 2 and 3). However, montane specialists from the most mesic sites were as likely to be induced into secondary dormancy under field experimental conditions as specialists from more arid warm and salt desert sites (Fig. 3). Results from multiple studies of in situ seed banks (Smith et al., Reference Smith, Meyer and Anderson2008; Allen et al. Reference Allen, Meyer and Beckstead2013) suggest that seed populations at drier sites are more likely to enter secondary dormancy and form a persistent seed bank than those at mesic sites, but these differences could be due to the higher probability of dormancy-inducing conditions associated with inadequate autumn precipitation at drier sites rather than to genetic differences in propensity for secondary dormancy induction.
The successful use of Ψb(50) as an indicator of secondary dormancy status adds further support to the value of using this parameter as an index of germination status in several contexts (Christensen et al., Reference Christensen, Meyer and Allen1996; Bauer et al., Reference Bauer, Meyer and Allen1998; Meyer et al., Reference Meyer, Debaene-Gill and Allen2000; Alvarado and Bradford, Reference Alvarado and Bradford2005; Bair et al., Reference Bair, Meyer and Allen2006; Meyer and Allen, Reference Meyer and Allen2009). The second and more complex modelling approach, involving recalculation of all hydrothermal parameters to describe seed dormancy status has rarely been used (Gianinetti and Cohn, Reference Gianinetti and Cohn2007; Bazin et al., Reference Bazin, Batlla, Dussert, El-Maarouf-Bouteau and Bailly2011). This approach has the advantage that it is possible to detect a decrease in the remaining hydrothermal time requirement as seeds accumulate hydrothermal time (evidenced by a reduction in θHT) even as they are simultaneously induced into secondary dormancy [evidenced by the increase in Ψb(50)]. As ‘fast’ seeds (with initially low Ψb values) are germinating, ‘slow’ seeds (those with higher Ψb values) may still accumulate hydrothermal time but also slowly enter dormancy. As with other threshold-type models, a normal distribution of Ψb values within the population means that some seeds would germinate more quickly and escape secondary dormancy induction, while the slowest seeds would fail to accumulate enough hydrothermal time before secondary dormancy was induced. This clearly results in a truncation of the original normal distribution of Ψb values via germination, an effect not taken into account in our modelling procedure. The effect of hydrothermal time accumulation on estimated θHT for the December retrieval was nonetheless strongly evident, even when only ungerminated seeds were used in model development (Table 1). Furthermore, using this estimate for θHT rather than the estimate from the initial hydrothermal time equations gave much better fit to the observed December retrieval data (Fig. 5). For retrieval data collected during dormancy release that occurred the following summer, fitted hydrothermal time equations again had θHT values similar to initial values. Over the winter and into the summer months it is likely that the previously gained hydrothermal progress was lost or ‘forgotten’, as also occurs when imbibed seeds that have nearly completed germination are subjected to rapid drying (Debaene-Gill et al., Reference Debaene-Gill, Allen and White1994; Meyer and Allen, Reference Meyer and Allen2009).
There was no clear pattern for Ψb(50) to vary as a function of incubation temperature during dormancy induction in the field in December, suggesting that secondary dormancy induction in B. tectorum is not a simple reversal of the process of dormancy loss (Table 1; Fig. 6). However, the pattern for dormancy loss in the summer for secondarily dormant seeds of both seed populations clearly followed the same pattern as for primary dormancy loss, i.e. Ψb(50) was always higher at the higher incubation temperature during this process. There would be little ecological advantage to increased dormancy at high temperature during dormancy induction, which takes place at low temperature, whereas the advantages for maintaining dormancy longer at high temperature in the summer are clear (Christensen et al., Reference Christensen, Meyer and Allen1996; Meyer and Allen, Reference Meyer and Allen2009). We propose referring to the process of secondary dormancy loss under dry summer conditions as secondary after-ripening.
To our knowledge, this is the first time hydrothermal modelling has been used explicitly to explain secondary dormancy induction and loss in seeds. We found that changes in Ψb(50) are useful for modelling these processes, as has previously been reported for modelling primary dormancy (Christensen et al., Reference Christensen, Meyer and Allen1996; Bauer, Reference Bauer, Meyer and Allen1998; Meyer et al., Reference Meyer, Debaene-Gill and Allen2000; Alvarado and Bradford, Reference Alvarado and Bradford2005; Bair et al., Reference Bair, Meyer and Allen2006; Gianinetti and Cohn, Reference Gianinetti and Cohn2007; Meyer and Allen, Reference Meyer and Allen2009). The conceptual simplicity of the hydrothermal time approach makes it a logical tool for examination of secondary seed dormancy acquisition and loss in other species.
Acknowledgements
We thank Kedra Foote for carrying out the 2008–2009 retrieval study.
Financial support
This research was funded in part by grants to P.S.A. and S.E.M. from the Joint Fire Sciences Program (JFSP-2007-1-3-10; JFSP-2011-S-2-6).
Conflicts of interest
None.







