We use cookies to distinguish you from other users and to provide you with a better experience on our websites. Close this message to accept cookies or find out how to manage your cookie settings.
Online ordering will be unavailable from 17:00 GMT on Friday, April 25 until 17:00 GMT on Sunday, April 27 due to maintenance. We apologise for the inconvenience.
This journal utilises an Online Peer Review Service (OPRS) for submissions. By clicking "Continue" you will be taken to our partner site
https://mc.manuscriptcentral.com/qrb-discovery.
Please be aware that your Cambridge account is not valid for this OPRS and registration is required. We strongly advise you to read all "Author instructions" in the "Journal information" area prior to submitting.
To save this undefined to your undefined account, please select one or more formats and confirm that you agree to abide by our usage policies. If this is the first time you used this feature, you will be asked to authorise Cambridge Core to connect with your undefined account.
Find out more about saving content to .
To send this article to your Kindle, first ensure [email protected] is added to your Approved Personal Document E-mail List under your Personal Document Settings on the Manage Your Content and Devices page of your Amazon account. Then enter the ‘name’ part of your Kindle email address below. Find out more about sending to your Kindle.
Find out more about saving to your Kindle.
Note you can select to save to either the @free.kindle.com or @kindle.com variations. ‘@free.kindle.com’ emails are free but can only be saved to your device when it is connected to wi-fi. ‘@kindle.com’ emails can be delivered even when you are not connected to wi-fi, but note that service fees apply.
Single-stranded nucleic acid (ssNA) binding proteins must both stably protect ssNA transiently exposed during replication and other NA transactions, and also rapidly reorganize and dissociate to allow further NA processing. How these seemingly opposing functions can coexist has been recently elucidated by optical tweezers (OT) experiments that isolate and manipulate single long ssNA molecules to measure conformation in real time. The effective length of an ssNA substrate held at fixed tension is altered upon protein binding, enabling quantification of both the structure and kinetics of protein–NA interactions. When proteins exhibit multiple binding states, however, OT measurements may produce difficult to analyze signals including non-monotonic response to free protein concentration and convolution of multiple fundamental rates. In this review we compare single-molecule experiments with three proteins of vastly different structure and origin that exhibit similar ssNA interactions. These results are consistent with a general model in which protein oligomers containing multiple binding interfaces switch conformations to adjust protein:NA stoichiometry. These characteristics allow a finite number of proteins to protect long ssNA regions by maximizing protein–ssNA contacts while also providing a pathway with reduced energetic barriers to reorganization and eventual protein displacement when these ssNA regions are diminished.
DNA unzipping by nanopore translocation has implications in diverse contexts, from polymer physics to single-molecule manipulation to DNA–enzyme interactions in biological systems. Here we use molecular dynamics simulations and a coarse-grained model of DNA to address the nanopore unzipping of DNA filaments that are knotted. This previously unaddressed problem is motivated by the fact that DNA knots inevitably occur in isolated equilibrated filaments and in vivo. We study how different types of tight knots in the DNA segment just outside the pore impact unzipping at different driving forces. We establish three main results. First, knots do not significantly affect the unzipping process at low forces. However, knotted DNAs unzip more slowly and heterogeneously than unknotted ones at high forces. Finally, we observe that the microscopic origin of the hindrance typically involves two concurrent causes: the topological friction of the DNA chain sliding along its knotted contour and the additional friction originating from the entanglement with the newly unzipped DNA. The results reveal a previously unsuspected complexity of the interplay of DNA topology and unzipping, which should be relevant for interpreting nanopore-based single-molecule unzipping experiments and improving the modeling of DNA transactions in vivo.
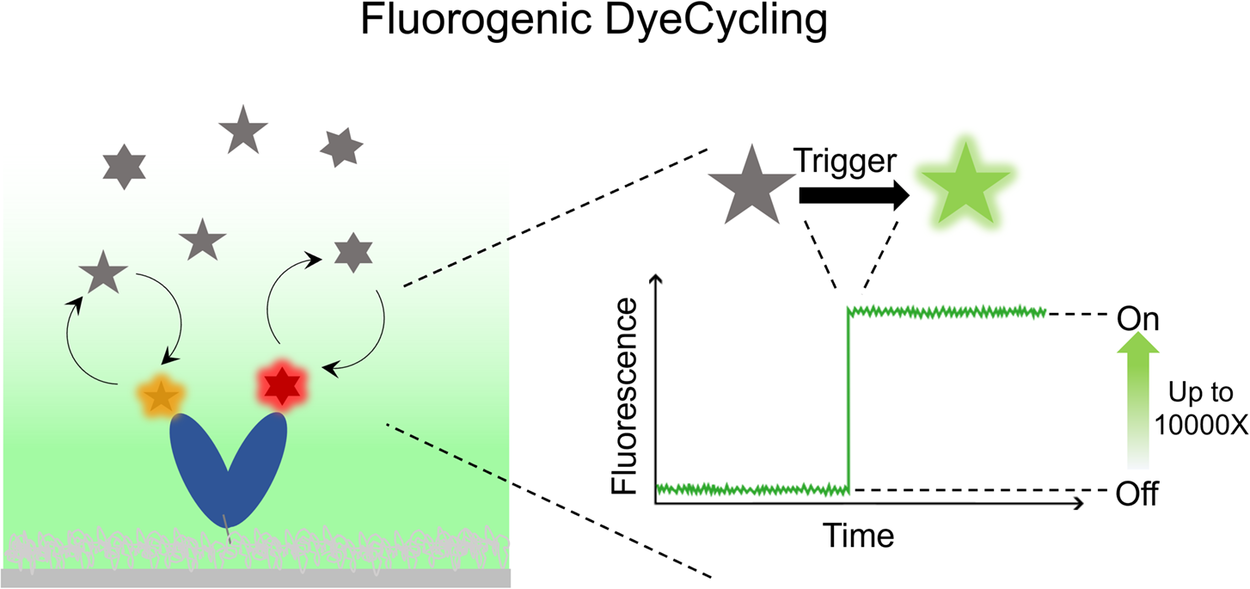
Single Molecule Förster Resonance Energy Transfer (smFRET) is a popular technique to directly observe biomolecular dynamics in real time, offering unique mechanistic insight into proteins, ribozymes, and so forth. However, inevitable photobleaching of the fluorophores puts a stringent limit on the total time a surface-tethered molecule can be monitored, fundamentally limiting the information gain through conventional smFRET measurements. DyeCycling addresses this problem by using reversibly – instead of covalently – coupled FRET fluorophores, through which it can break the photobleaching limit and theoretically provide unlimited observation time. In this perspective paper, we discuss the potential of various fluorogenic strategies to suppress the background fluorescence caused by unbound, freely diffusing fluorophores inherent to the DyeCycling approach. In comparison to nanophotonic background suppression using zero-mode waveguides, the fluorogenic approach would enable DyeCycling experiments on regular glass slides with fluorogenic FRET probes that are quenched in solution and only fluoresce upon target binding. We review a number of fluorogenic approaches and conclude, among other things, that short-range quenching appears promising for realising fluorogenic DyeCycling on regular glass slides. We anticipate that our discussion will be relevant for all single-molecule fluorescence techniques that use reversible fluorophore binding.
Integrins are critical transmembrane receptors that connect the extracellular matrix (ECM) to the intracellular cytoskeleton, playing a central role in mechanotransduction – the process by which cells convert mechanical stimuli into biochemical signals. The dynamic assembly and disassembly of integrin-mediated adhesions enable cells to adapt continuously to changing mechanical cues, regulating essential processes such as adhesion, migration, and proliferation. In this review, we explore the molecular clutch model as a framework for understanding the dynamics of integrin – ECM interactions, emphasizing the critical importance of force loading rate. We discuss how force loading rate bridges internal actomyosin-generated forces and ECM mechanical properties like stiffness and ligand density, determining whether sufficient force is transmitted to mechanosensitive proteins such as talin. This force transmission leads to talin unfolding and activation of downstream signalling pathways, ultimately influencing cellular responses. We also examine recent advances in single-molecule DNA tension sensors that have enabled direct measurements of integrin loading rates, refining the range to approximately 0.5–4 pN/s. These findings deepen our understanding of force-mediated mechanotransduction and underscore the need for improved sensor designs to overcome current limitations.