


Folate status in women of childbearing age is important due to its role in the prevention of neural tube defects (NTD). Previous studies have observed a differential association between obesity and serum and red-blood-cell (RBC) folate concentrations in men, non-pregnant women of childbearing age and postmenopausal women( Reference Kimmons, Blanck and Tohill 1 – Reference Tinker, Hamner and Berry 5 ). Specifically, serum folate was found to be negatively associated, while RBC folate was positively associated, with BMI among non-pregnant adults( Reference Kimmons, Blanck and Tohill 1 – Reference Tinker, Hamner and Berry 5 ). The mechanisms for this differential association of obesity with serum and RBC folate concentrations are not clearly understood. Some investigators have suggested that the altered pharmacokinetics and distribution of folate in obese v. normal-weight persons may explain, at least in part, these differences( Reference Stern, Matok and Kapur 6 , Reference da Silva, Hausman and Kauwell 7 ). It was observed that the peak serum folate concentrations of obese and normal-weight women were statistically different after a single oral dose of folic acid (0·4 mg), but there was no difference in overall area under the curve, indicating that obesity does not impair the absorption of folic acid but may lead to redistribution of folate from circulation into tissue. Folate requirements are higher in pregnancy in order to maintain placental and fetal growth, and accelerated folate catabolism and haemodilution during pregnancy can also result in decreased circulating folate concentrations( Reference Higgins, Quinlivan and McPartlin 8 – Reference Açkurt, Wetherilt and Löker 10 ). Given the potential physiological and pharmacokinetic differences in pregnant v. non-pregnant populations, the association of obesity with serum and RBC folate concentrations may be altered. Owing to the observed differential associations of folate with BMI in previous studies, the standard RBC folate cut-off approach for assessing NTD risk in obese women may be limited. Lower serum folate in obese women may reflect a redistribution of folate in response to the higher mass of maternal tissue, potentially limiting the amount of folate available to the developing embryo which accesses folate through serum folate rather than RBC folate, thus increasing the risk of NTD. The identification of subgroups of women who may be susceptible to low folate status or respond differently to folic acid supplementation will ensure that public health interventions can be tailored to meet the needs of the entire population. We therefore examined the association of BMI with serum and RBC folate concentrations in pregnancy using data collected in the Ottawa and Kingston (OaK) Birth Cohort study( Reference Walker, Finkelstein and Rennicks White 11 ).
Method and materials
Data source
We used a subset of data from the OaK Birth Cohort, a prospective cohort study that recruited pregnant women at The Ottawa Hospital and Kingston General Hospital, Ontario, Canada, from September 2002 to April 2009. The subset included participants recruited from April 2008 to April 2009, from whom both serum and RBC folate were measured (Fig. 1). Participants were recruited between 12 and 20 gestational weeks and later delivered in the Ottawa–Carleton and Kingston regions. Participants were referred to the research team by clinic staff and asked to participate at their first antenatal visit. They were recruited into the study after information about the purpose of the study was provided and their written consent was obtained.

Fig. 1 Flowchart of participants, illustrating the procedure of study group identification (OaK, Ottawa and Kingston; GA, gestational age; RBC, red blood cells; MTHFR, methylenetetrahyrofolate reductase gene)
At the time of recruitment, participants’ self-reported demographic and lifestyle information including age, race, education, household income, folic acid supplementation and smoking. Nurses measured current height and weight at recruitment. Participants provided the brand name(s) of vitamin supplement(s) that were being consumed at the time of recruitment. Total daily supplemental folic acid intake was calculated based on the frequency of use and folic acid dose from all vitamin supplements. For women who were using more than one vitamin supplement, the total daily supplemental folic acid dose was calculated as the combined intake from all supplement sources.
Fasting blood samples were collected at the time of routine blood work, in order to avoid additional venepuncture. The blood samples were drawn from the antecubital vein or from the hand of the participating women at gestational age between 12 and 20 weeks, and measured in batches every month (i.e. the samples were stored for up to 1 month). Blood samples for serum folate testing were collected in serum separator tubes (Becton Dickinson, Franklin Lakes, NJ, USA) and allowed to clot. Samples were centrifuged for 10 min at 3000 g to separate serum. Serum was then removed and stored at −20°C until analysis. Folate concentrations were measured using the Beckman Coulter Access 2 and Unicel DxI 800 immunoassay analysers using the manufacturer’s reagents (Beckman Coulter, Brea, CA, USA).
Blood samples for measuring RBC folate concentration were collected in 3 ml purple-top EDTA tubes. Haematocrit was determined in fresh samples using an OSM3 hemoximeter (Radiometer, Copenhagen, Denmark). Samples were frozen at −20°C until analysis, at which point they were thawed and cells were lysed using Access RBC Folate Lysing Agent (Beckman Coulter). RBC folate concentrations were quantified using the Beckman Coulter UniCel DxI 800 Access Immunoassay System measured by competitive protein-binding assay. RBC folate concentrations were adjusted for haematocrit as described by the manufacturer.
Blood samples for genotyping the 677C→T polymorphism in the methylenetetrahyrofolate reductase gene (MTHFR) were collected in K2EDTA Vacutainer tubes. Samples were centrifuged at 1100 g and DNA was extracted using the BioRobot M48 and MagAttract DNA Blood Midi Kit (Qiagen, Hilden, Germany). PCR was used to amplify DNA regions of interest and genotyping was performed using the ABI 3130xl Genetic Analyzer and the ABI Prism SnaPshot Multiplex Kit (Applied Biosystems Inc., Foster City, CA, USA), which allows genotyping of several SNP in the same reaction( Reference Di Cristofaro, Silvy and Chiaroni 12 ). The laboratory procedures for genotyping have been described previously( Reference Walker, Finkelstein and Rennicks White 11 , Reference Ferraro, Barrowman and Prud’homme 13 ). For quality control, all assays were tested in hospital laboratories that are accredited by Ontario Laboratory Accreditation (OLA) and the Institute for Quality Management in Healthcare (IQMH), and are ISO certified for quality, competence and beyond.
Statistical analysis
Biomarker concentrations were non-normally distributed; therefore the geometric mean was used in our analysis. In regression analyses, logarithmic transformations were applied to variables with skewed distributions.
BMI was calculated and categorized into normal and underweight (<25·0 kg/m2), overweight (25·0–29·9 kg/m2) and obese (≥30·0 kg/m2). Underweight cases (n 41) were combined with normal-weight cases due to limited sample size. Use of folic acid supplements was categorized as: (i) no supplement; (ii) supplement with folic acid intake ≤1 mg/d; and (iii) supplement with folic acid intake >1 mg/d.
Characteristics of the participants were described and compared across BMI categories using χ 2 tests. ANCOVA was used to examine folate concentrations associated with participant characteristics, including BMI, daily folic acid supplement intake, MTHFR 677C→T genotype (CC, CT and TT), maternal age, gestational age at blood sample collection, race, maternal education, household annual income and smoking.
Multiple linear regression analysis was used to examine associations of folate concentrations with BMI, in all samples and in subgroups stratified by MTHFR 677C→T genotype or folic acid supplementation. Interaction between BMI and genotype was tested in the overall model. Supplementary analysis using the pre-gravid weight (self-reported) was also performed. The level of significance was set at 0·05. Statistical analyses were performed using the statistical software package SAS version 9.2.
Results
A total of 889 participants who had complete records for both serum and RBC folate concentrations were identified in the cohort. Seven participants reported the use of vitamin supplements but did not include information on dose, and thirteen women missed information on MTHFR 677C→T genotype, leaving 869 participants for the final analysis (Fig. 1).
The overall geometric mean concentration of serum folate was 45·7 (sd 18·7) nmol/l and the geometric mean concentration of RBC folate was 1549 (sd 378) nmol/l. Serum folate <7·0 nmol/l was non-existent, and only one participant’s serum folate was below 10 nmol/l. Demographic characteristics are shown in Table 1. Thirty-five participants (4 %) were not taking any vitamin supplements. The proportion of overweight and obesity were 26 % and 23 %, respectively. Twelve per cent of participants were homozygous for the MTHFR 677C→T variant (TT). Gestational age at recruitment, education, annual income, folic acid supplementation and MTHFR 677C→T genotype were statistically different among the BMI categories. Women with BMI ≥ 30·0 kg/m2 had a higher gestational age at recruitment, lower education, lower income, lower supplement use and were more likely to have the MTHFR CC variant.
Table 1 Demographic characteristics of the study participants by BMI categoryFootnote ‡: pregnant women (n 869) recruited between 12 and 20 weeks of gestation, Ottawa and Kingston (OaK) Birth Cohort study, April 2008–April 2009

GA, gestational age; MTHFR, methylenetetrahyrofolate reductase gene.
‡ Data are presented as n and %. BMI = [weight (kg)]/[height (m)]2. BMI was measured in pregnancy (between 12 and 20 weeks of gestation); this may lead to misclassification of BMI category.
§ P value of χ 2 test.
|| Folic acid from supplements, not including folic acid from fortified foods or naturally occurring food folate.
Table 2 shows the determinants of serum and RBC folate concentrations. BMI showed an inverse association with serum folate but a positive association with RBC folate whether adjusted for or not (Table 2). Serum folate was lower with each additional variant allele such that CC > CT > TT; however, the trend was not statistically significant. Conversely, unadjusted and adjusted RBC folate was higher in participants homozygous for the T allele such that TT > CC and TT > CT. Folic acid supplementation was associated with higher serum and RBC folate concentrations. Gestational age at recruitment was a determinant of serum folate, and maternal age, gestational age at recruitment, education and smoking were determinants of RBC folate.
Table 2 Geometric mean concentrations of serum and RBC folate by characteristics of participants: pregnant women (n 869) recruited between 12 and 20 weeks of gestation, Ottawa and Kingston (OaK) Birth Cohort study, April 2008–April 2009

RBC, red blood cell; MTHFR, methylenetetrahyrofolate reductase gene; GA, gestational age.
* Significantly different from the first category of the listed variables (P < 0·05).
† Significantly different from the second category of the listed variables (P < 0·05).
‡ Adjusted for all other variables listed.
Table 3 shows the association between BMI and serum or RBC folate, for all participants and in subgroups stratified by MTHFR 677C→T genotype or folic acid supplementation, adjusted for demographic variables. Overall, serum folate was inversely associated with BMI and RBC folate was positively associated with BMI. Interaction between BMI and genotype was not significant (P>0·05) in the overall model (data not shown). When stratified by MTHFR genotype, serum folate was inversely associated with BMI in women with CC and CT variants, while RBC folate was positively associated with BMI in women with the CC variant. In women with the TT variant, both associations were insignificant, but the direction remained consistent with that observed for the CC and CT variants.
Table 3 Association of BMI with serum and RBC folate, stratified by MTHFR 677C→T genotype and folic acid supplementationFootnote ‡; pregnant women (n 869) recruited between 12 and 20 weeks of gestation, Ottawa and Kingston (OaK) Birth Cohort study, April 2008–April 2009
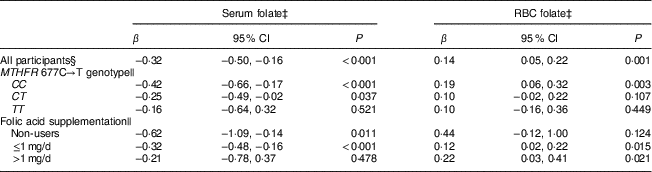
RBC, red blood cell; MTHFR, methylenetetrahyrofolate reductase gene; β, the partial regression coefficient of log-transformed BMI.
‡ BMI, serum folate and RBC folate concentrations were log-transformed.
§ Regression coefficient of BMI in all participants, adjusted for MTHFR 677C→T genotype, folic acid supplementation, age, gestational age at recruitment, race, maternal education, household annual income and smoking.
|| Regression coefficient of BMI in subgroups stratified by MTHFR 677C→T genotype or folic acid supplementation, adjusted for age, gestational age at recruitment, race, maternal education, household annual income and smoking.
Supplementary analysis using self-reported pre-gravid weight yielded similar results (data not shown).
Discussion
Our study confirmed a differential association of BMI with serum and RBC folate concentrations in pregnant women, such that BMI and serum folate concentration had an inverse relationship, and BMI and RBC folate concentration had a positive relationship. The differential associations were not significant in women with CT or TT genotype, although the direction of these associations was consistent in women with MTHFR CC genotype. Our data indicate that the differential association between serum and RBC folate and BMI observed in non-pregnant obese women likely persists into pregnancy. Serum folate was also found to be inversely associated with BMI at mid and late pregnancy in Korean women, although no RBC folate was measured simultaneously in that study( Reference Kim, Hwang and Kim 14 ). In both Canadian and American populations, non-pregnant obese women, including those of childbearing age, have been shown to have lower plasma folate and higher RBC folate in comparison to normal-weight women( Reference Bird, Ronnenberg and Choi 2 , Reference Tinker, Hamner and Berry 5 , Reference da Silva, Hausman and Kauwell 7 ). These associations remained even after adjustment for dietary and supplemental folate intake, an important consideration given the observed pattern of lower folate intake among obese women( Reference Bird, Ronnenberg and Choi 2 , Reference Tinker, Hamner and Berry 5 ).
The associations were significant only for women with CC genotype and not significant for those with TT owing to limited sample size in the TT group, as well as increased measurement error attributed to the use of a competitive protein-binding assay to measure RBC folate, which is an important limitation of the present study. We observed a CC>CT > TT pattern in serum folate concentration, but a reversed TT>CC and TT>CT pattern for RBC folate concentration. In a meta-analysis, Tsang et al. reported a CC>CT>TT pattern in serum and RBC folate concentrations measured by microbiological assay, which was reversed for RBC folate concentrations measured by protein-binding assay( Reference Tsang, Devine and Cordero 15 ). The differences between the assays are due to differences in affinity of the assays for various folate forms. Plasma folates are exclusively monoglutamates and the majority are in the form of 5-methyltetrahydrofolate, whereas the majority of RBC folates are variable-length, long-chain polyglutamate 5-methyltetrahydrofolate. RBC folates must be hydrolysed into the monoglutamate form. The differences in affinities among these folate forms, as well as inconsistent proportion of folate forms among those with MTHFR 677C→T homozygous genotype (TT), impair the interpretation of RBC folate concentration measured by protein-binding assay( Reference Bagley and Selhub 16 ).
There are several other limitations to our study. First, the study design is cross-sectional, and serum and RBC folate were measured in early pregnancy; therefore, it is not possible to determine whether these associations hold through late pregnancy when haemodilution could have a significant effect. Second, the type and dose of supplements were self-reported and may be subject to measurement error due to inaccurate patient report and irregular pattern of use. We did not conduct dietary surveys on participants. Previous studies reported that obese women tended to under-report their food intake( Reference Scagliusi, Polacow and Artioli 17 , Reference Scagliusi, Ferriolli and Pfrimer 18 ), and folate from naturally occurring food sources exhibits variable and incomplete bioavailability, which can be affected by physiological conditions, pharmaceuticals and genetic polymorphisms( Reference Prinz-Langenohl, Brönstrup and Thorand 19 – Reference Obeid, Koletzko and Pietrzik 21 ). Third, BMI was measured in early pregnancy (95 % were measured before 15 weeks) and this may lead to misclassification of obesity, although the weight gain in early second trimester is generally small( 22 ). Finally, samples were measured in batches and the storage time varied, but was no longer than 1 month. Serum folate stored in a frost-free freezer at −20°C for even a short period may be relatively unstable and sensitive to minor temperature fluctuations( Reference Bailey 23 ). This may contribute to the variation in folate concentrations.
There are several strengths of the present study. First, to our knowledge, it is the first study to examine the association of obesity with serum and RBC folate concentrations in pregnant women. Because of the difference in folate metabolism between the non-pregnant and pregnant populations, it is important to examine if the observed differential association between obesity and folate concentrations measured from serum v. RBC would persist in a pregnant population( Reference Kimmons, Blanck and Tohill 1 – Reference Tinker, Hamner and Berry 5 ). Second, our study was based on data collected in a prospective cohort, which mitigates recall bias. Third, serum folate and RBC folate concentrations in our study were derived from the same fasted participants, thereby mitigating bias/confounding when examining the association between serum and RBC folate with BMI.
It is not clearly understood why obesity was inversely associated with serum folate concentration but positively associated with RBC folate concentration. Obese women may demonstrate lower serum folate concentrations due to volumetric dilution of the blood. Alternatively, higher RBC folate may be due to the redistribution of folate from circulation into tissues. Cell culture and animal studies have shown that the expression of tissue folate receptors and transporters is responsive to folate concentrations: folate deficient conditions induce the expression of reduced folate carrier in rat intestine and cultured human Caco2 cells( Reference Said, Chatterjee and Haq 24 , Reference Subramanian, Chatterjee and Said 25 ) whereas supplementation reduces its expression( Reference Ashokkumar, Mohammed and Vaziri 26 ). It could be hypothesized that the delay in absorption of folate observed in obese women may alter the expression of cellular folate transporters/receptors such that it promotes cellular uptake.
Folate status in women of childbearing age is important due to the role of folate in the prevention of NTD. The placenta takes up folate from the maternal plasma by the reduced folate carrier, the proton-coupled folate transporter and the folate receptor α( Reference Yasuda, Hasui and Yamamoto 27 , Reference Solanky, Requena Jimenez and D’Souza 28 ). Placental uptake of circulating folate ensures an adequate folate supply to the developing fetus. Obese women are at increased risk for NTD-affected pregnancy( Reference McMahon, Liu and Zhang 29 – Reference Waller, Shaw and Rasmussen 32 ). A proportion of the NTD risk associated with maternal obesity may be due to lower circulating folate in obese women, which can be corrected to some degree with higher folate intake( Reference McMahon, Liu and Zhang 29 ). On the other hand, it must be noted that a proportion of the NTD risk associated with obesity appears to be independent of folate status and may be due to other metabolic disturbances( Reference Parker, Yazdy and Tinker 33 ). Obese women of childbearing age may therefore require supplemental folic acid to increase or maintain a plasma folate status that is associated with maximal protection from NTD. However, folate status associated with NTD protection is currently expressed in terms of RBC folate( 34 ). Because of the observed differential association of RBC folate with BMI, the standard RBC folate cut-off approach for assessing NTD risk in obese women may be limited. The relationship between RBC folate and serum folate, and how they relate to NTD risk reduction, may need to be determined for women of all BMI categories to determine appropriate ‘cut-offs’ for NTD protection.
A second consideration is whether obese women respond to folic acid supplementation similar to normal-weight women. Folic acid supplement use did not necessarily mitigate the observed differential association of BMI with serum and RBC folate in these pregnant women. A pharmacokinetic study found that obese women demonstrated a lower peak serum folate concentration after a 400 µg bolus folic acid dose and a lower area under the curve in the absorptive phase (0–3 h post-bolus), but no difference in the overall area under the curve( Reference da Silva, Hausman and Kauwell 7 ). The data indicate that obesity may delay the absorption of folic acid but that overall absorption is not impaired. It should be noted that the women in our study had a folate status associated with maximal protection from NTD risk (≥906 nmol/l)( Reference Daly, Kirke and Molloy 35 ). In our study, the overall mean RBC folate was 1549 (sd 378) nmol/l and 1275 (95 % CI 1155, 1409) nmol/l in supplement non-users. This is comparable to findings from the Canadian Health Measures Survey (2007–2009) where mean RBC folate (also measured by immunoassay) was 1142 (95 % CI 1153, 1239) nmol/l in women of childbearing age who did not consume a folic acid supplement( Reference Shi, De Groh and MacFarlane 36 ). These findings are reflective of the mandatory folic acid fortification of white wheat flour in Canada. The relationship of obesity with folate status may be more of a concern in countries where fortification is not widespread and supplement use is lower.
The differential association of obesity with serum folate concentration v. RBC folate concentration observed in the present study deserves attention in research and perinatal health practice alike. The identification of subgroups of women who may be susceptible to low folate status or respond differently to folic acid supplementation will ensure that public health interventions can be tailored to meet the needs of the entire population. Obesity is a known risk factor for a number of adverse maternal and infant outcomes including pre-eclampsia( Reference Lisonkova and Joseph 37 , Reference Roberts, Bodnar and Patrick 38 ), gestational diabetes( Reference Baci, Üstüner and Keskin 39 , Reference Chu, Callaghan and Kim 40 ) and NTD( Reference Rasmussen, Chu and Kim 41 ). Folic acid deficiency is a known risk factor for NTD( Reference Czeizel and Dudas 42 , 43 ) and may be associated with other pregnancy complications such as pre-eclampsia( Reference Wen, Champagne and Rennicks White 44 , Reference Wen, Chen and Rodger 45 ) and other congenital anomalies( Reference Pei, Zhu and Zhu 46 , Reference Feng, Wang and Chen 47 ). As a result, in the study of the association between folate status and adverse maternal and infant outcomes, it is critical to use the appropriate biological specimens for the measurement of folate. Since BMI is positively associated with RBC folate but inversely associated with serum folate, the observed associations between folate status and pregnancy complications may be overestimated if serum folate is used while underestimated if RBC folate is used. Folate status varies substantially during pregnancy( Reference Higgins, Quinlivan and McPartlin 8 ), which can further complicate the accurate assessment of folate status.
In summary, we confirmed the differential associations of folate with BMI in pregnant women, indicating that the RBC folate cut-off approach for assessing NTD risk may be limited. Because absorption and metabolism of folate are modified by adiposity, RBC folate may not reflect freely available folate at cellular level in pregnant women.
Acknowledgements
Acknowledgements: The authors thank the staff and pregnant women at The Ottawa Hospital and Kingston General Hospital for their support and participation in this study. Financial support: This research was supported in part by the Canadian Institutes for Health Research (CIHR; grant numbers MOP53188, MOP 82802 and MCT-98030). The CIHR had no role in the design, analysis or writing of this article. Conflict of interest: None. Authorship: S.W.W. designed and initiated the study; S.W.W., M.R. and M.C.W. secured funding and resources for the study; M.S. performed statistical analysis of the data and drafted the manuscript; R.R.W. managed the implementation of the study; S.H.C., A.J.M., L.G., G.N.S., R.R.W. and S.W.W. made critical revisions to the manuscript; all authors reviewed and approved the manuscript. Ethics of human subject participation: This study was conducted according to the guidelines laid down in the Declaration of Helsinki and all procedures involving patients were approved by the research ethics boards of The Ottawa Hospital and Kingston General Hospital. Written informed consent was obtained from all patients.




