No CrossRef data available.
Published online by Cambridge University Press: 20 May 2024
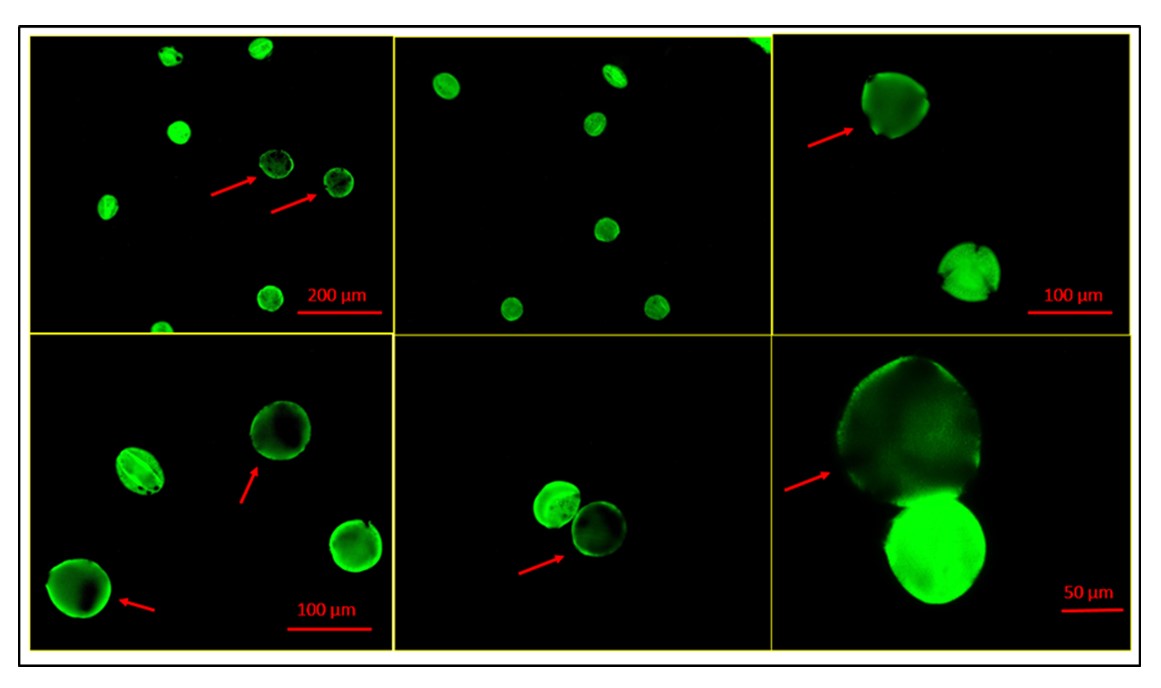
Bitter gourd is a highly nutritious vegetable and important medicinal plant of economic importance. The present study is focused on cytogenetical characterization of 12 accessions of bitter gourd from different parts of India, aiming to differentiate their karyotypes and outline diagnostic features of the chromosomes within each accession's haploid complement. All the accessions possess 2n = 22 numbers of chromosomes. The chromosomes mainly were metacentric (16‒22 chromosomes), and the presence or absence of sub-metacentric (0‒6 chromosomes) chromosomes. The length of the chromosomes varied from 0.83 to 1.93 μm among the accessions studied. Significant differences were obtained for the seven intra-chromosomal indices and four inter-chromosomal indices among the accessions. Principal component analysis and unweighted pair group method with arithmetic mean study revealed relatively distant positioning of individuals that advocated intraspecific phylogenetic relationships and higher karyoevolutionary affinity in bitter gourd accessions. In the meiotic study, regular meiotic behaviour indicates genetic stability and a stable sexual cycle in different accessions. The percentage of pollen viability of all the studied accessions was very high (89.41–94.11%), and these accessions can be considered to be good pollinators. The results obtained will guide characterizing the elite genotypes, genotypes management and designing effective breeding programmes and crop improvement programmes.