



No CrossRef data available.
Published online by Cambridge University Press: 17 April 2023
Western Himalayan region is a rich repository of common bean genetic resources evolved over generations of farmer selection. The bean variability encompasses various use categories such as dry, snap and shelled beans. The present study aimed to characterize 40 small-seeded red common bean genotypes collected from different districts of Kashmir valley as well as Poonch, Rajouri and Bhaderwah areas of Jammu which are world famous for their beans. Based on 5-year evaluation, there was substantial variability for various qualitative traits as well as quantitative traits. Among variability parameters, higher PCV and GCV values were recorded for plant height, seed yield per plant and number of pods per plant and low for days to maturity, seed length and pod length. High heritability was recorded for all the traits ranging from 78.72 to 94.87%. There was substantial variability in root traits scored in laboratory and greenhouse as well as canopy temperature depression and relative water content. Various genotypes were found resistant to bean common mosaic virus (WB-N-1, WB-N-11, WB-1318, WB-401, WB-451, WB-1634); anthracnose (WB-1634 WB-83, WB-258, WB-1282, WB-1304); angular leaf spot (WB-6, WB-1634, WB-258, WB-451, N-1 and N-10); Fusarium wilt (WB-341, WB-451, WB-N-1 and WB-1587) and Ascochyta blight (WB-6). Similarly WB-92 was highly resistant to shattering. The genotypes WB-341, WB-1634, WB-N-1 performed better under water stress, WB-N-1 and WB-1634 were resistant to multiple diseases and WB-1634 had better cooking quality. The genotypes identified can be used in bean improvement programmes and have also been submitted to NBPGR for long-term storage.

Sofi et al. supplementary material 1

Sofi et al. supplementary material 2
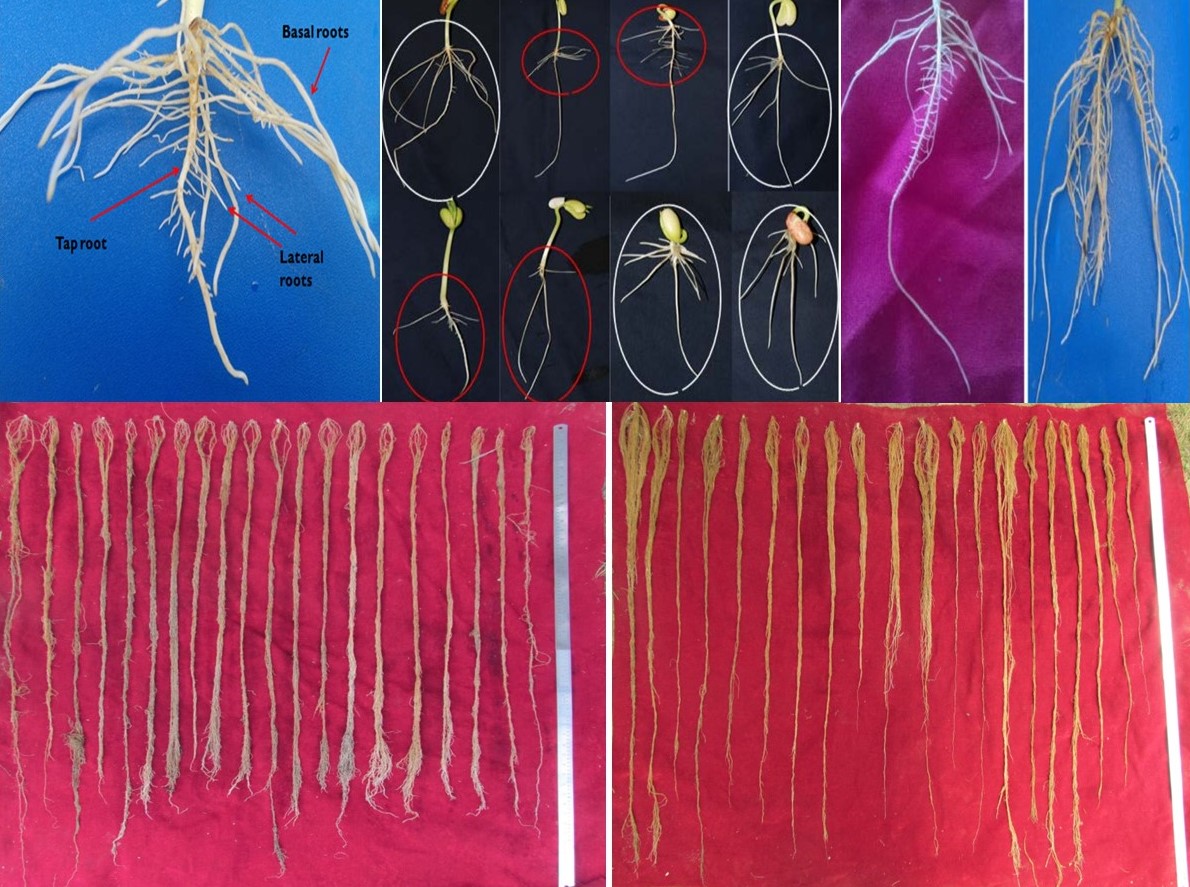

Sofi et al. supplementary material 3
