



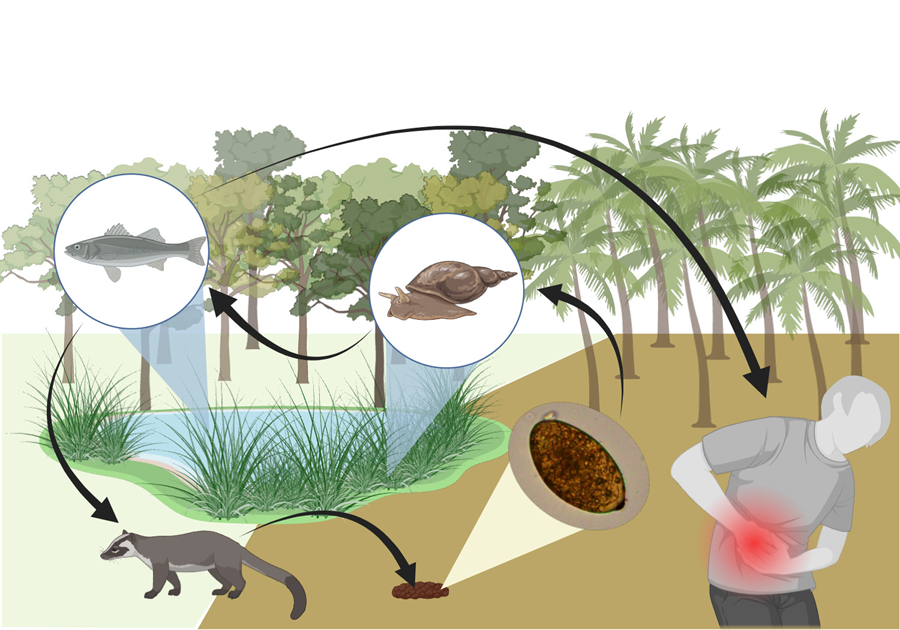
Introduction
Trematodiases are an important group of neglected tropical diseases caused by digenetic trematodes, which include the intestinal, liver, lung and blood flukes (Saijuntha et al., Reference Saijuntha, Andrews, Sithithaworn, Petney, Petney, Saijuntha and Mehlhorn2021). Intestinal trematodes and their hosts are highly diverse in Southeast Asia, where rapid and extensive changes in land use and land conversion can intensify the interactions among wildlife, domestic animals and humans, thereby creating new opportunities for parasite transmission (Jones et al., Reference Jones, Grace, Kock, Alonso, Rushton, Said, McKeever, Mutua, Young, McDermott and Pfeiffer2013). Over the last 7 decades, the Kinabatangan floodplain, an important biodiversity hotspot in Southeast Asia, has faced significant forest loss due to extensive commercial logging and industrial oil palm (Abram et al., Reference Abram, Xofis, Tzanopoulos, MacMillan, Ancrenaz, Chung, Peter, Ong, Lackman, Goossens and Ambu2014). Intensive human activity in the area (e.g. oil palm agriculture, ecotourism, low-scale fisheries), and high occurrence of wild and of domestic animals, can increase the likelihood of zoonotic pathogen transmission, particularly in areas frequented by infected livestock or where open defecation occurs (Fried et al., Reference Fried, Graczyk and Tamang2004). With large-scale conversion of forests to oil palm plantations, the dynamics of zoonotic diseases, including lesser-known parasites like echinostomatids, are inevitably being modified. Consequently, this study was undertaken to evaluate the presence of echinostomatid eggs in the feces of wildlife inhabiting the forest–oil palm plantation interface. The implications for potential public health concern in these human-dominated landscapes are also examined and discussed.
Materials and methods
Non-invasive surveys were carried out along 2 km linear transects running along the boundaries of each one of the 3 oil palm estates and the natural forest. Each transect was visited 3 consecutive days per month between April and July 2019, where fresh feces from wild and free-roaming domestic species were collected. A modified formalin-ethyl acetate sedimentation protocol was used to inspect samples for the presence of trematode eggs (Frias et al., Reference Frias, Hasegawa, Chua, Sipangkui, Stark, Salgado-Lynn, Goossens, Keuk, Okamoto and MacIntosh2021). Parasite prevalence, or the proportion of the host population infected with at least one trematode egg, was measured and the eggs per gram of feces (EPG) was calculated by taking the average of 5 replicate counts of all observed trematode eggs under a McMaster chamber's grid. Host species identification was conducted by amplifying a small fragment of the cytochrome b (cytb) gene using the L14724/H15915 primer pair (Irwin et al., Reference Irwin, Kocher and Wilson1991). Samples positive for trematodes through microscopy underwent DNA sequencing of both the nuclear ribosomal 18S gene and the mitochondrial cytochrome oxidase I (COI) gene. Amplification and sequencing of these genes were performed using 18S gene specific primers (18S9modF, 18S637modR) and COI specific primers (Dice1F, Dice11R and Dice14R) following established PCR protocols (Moszczynska et al., Reference Moszczynska, Locke, McLaughlin, Marcogliese and Crease2009; Van Steenkiste et al., Reference Van Steenkiste, Locke, Castelin, Marcogliese and Abbott2015).
Results
A total of 49 feces from 3 different forest–oil palm interface areas were collected. Trematode eggs were detected through microscopy in 28 samples (57.14%) from Asian palm civets (Paradoxurus hermaphroditus), leopard cats (Prionailurus bengalensis), domestic dogs (Canis lupus familiaris), domestic cats (Felis catus), long-tailed macaques (Macaca fascicularis) and humans (Table 1). Echinostomatid eggs detected in feces were ovoid to ellipsoid with a pronounced operculum, and 135 (129–145) μm × 75 (70–82) μm (n = 6) in size (Fig. S1), compatible with size and shape descriptions of human-infecting echinostomes (Esteban et al., Reference Esteban, Muñoz-Antoli, Toledo and Ash2019). Parasite DNA was only successfully amplified for COI in 6 samples from civets (GenBank accession numbers LC773275-LC773280), displaying a close genetic similarity of 97.05%–97.64% with parasites of the genus Artyfechinostomum spp. (Echinostomatidae) in the NCBI database, and not previously reported in Sabah (Fig. 1).
Table 1. Prevalence and abundance of trematode eggs found in feces of wildlife, domestic animals and humans, as detected through microscopy


Figure 1. Phylogenetic tree depicting the relationship between our samples (508 bp) and other related echinostomes inferred from 1000 replicates of partial COI sequences based on the neighbour-joining method and by using Fasciolopsis buski as an outgroup. Evolutionary distances were computed by using the Kimura 2-parameter methods and are presented as number of base substitutions per site. Terminal nodes in colour denote specimens collected in the current study and the other sequences were retrieved from GenBank. Tree reconstruction was conducted in Geneious Prime v. 2023.2.
Discussion
Despite being one of the fastest-growing economies in Southeast Asia, parasitic infections are still widespread in Malaysia. Rural and agricultural areas in Sabah are often remote, with limited access to healthcare, drinking water and poor environmental sanitation, and where the presence of intermediate and reservoir hosts can perpetuate parasite transmission (Zin et al., Reference Zin, SabaiAung, Saupin, Myint, KhinSN, Aung and Shamsu2015). In this study, leopard cats and Asian palm civets exhibited the highest parasite prevalence. Leopard cats and civets are not only native to most of Asia but are also more likely to frequent human settings, providing an insight into their potential as reservoir hosts for various pathogens (Guerrero-Sánchez et al., Reference Guerrero-Sánchez, Wilson, González-Abarzúa, Kunde, Goossens, Sipangkui and Frias2022). Free-ranging cats are not frequently found in the studied interface areas; however, plantation workers do own domestic cats and dogs that are often allowed to roam freely, as observed during our surveys. Despite their smaller population, dogs are a constant presence in these areas, as they play an important role in accompanying plantation workers. Dogs had the highest parasite egg output in this study, suggesting that infected individuals can end up contributing significantly to parasite environmental pollution, and highlighting the importance of including domestic animals in public health strategies aimed at controlling food-borne trematodiases in humans. Additionally, 9 samples originally presumed to be from carnivores were found to be from humans, with a reported prevalence (77.7%) that is notably higher than previous studies on echinostomes in Southeast Asia (7%–17% and 0.7%–1.8% for E. ilocanum [Sohn et al., Reference Sohn, Kim, Yong, Eom, Jeong, Kim, Kang, Kim, Park, Ji and Sinuon2011a; Chai and Jung, Reference Chai, Jung, Toledo and Fried2019], 7.5%–22.4% for E. revolutum [Sohn et al., Reference Sohn, Chai, Yong, Eom, Yoon, Sinuon, Socheat and Lee2011b] and an average of 43% for E. lindoense [Carney et al., Reference Carney, Sudomo and Purnomo1980], but see [Chai, Reference Chai, Fried and Toledo2009]). Our findings highlight the importance of implementing epidemiological surveys in these human–wildlife interfaces using a more holistic approach. In Sabah, the intermediate hosts for E. lindoense and E. revolutum have not yet been found, whereas the intermediate freshwater snail hosts for A. malayanum have been reported from at least three districts, including the study area (Table S1), suggesting that all the necessary components for the completion of this parasite's life cycle are present.
Despite having its limitations, fecal samples are often the only non-invasive method available to obtain information on gastrointestinal parasites in wildlife (Bechert, Reference Bechert, Miller and Fowler2012). Only 21.4% of the samples detected as positive by microscopy were amplified by PCR. The low amplification rate could be attributed to the presence of DNA polymerase inhibitors in feces (Stensvold et al., Reference Stensvold, Saijuntha, Sithithaworn, Wongratanacheewin, Strandgaard, Ørnbjerg and Johansen2006; Lamaningao et al., Reference Lamaningao, Kanda, Laimanivong, Shimono, Darcy, Phyaluanglath, Mishima and Nishiyama2017), but it is also possible that more than one trematode species was present in the samples, and that primer affinity was not sufficient to characterize all of them.
The Kinabatangan floodplain's fragmented landscape creates a network of intricate channels and drains interconnected with the river and lakes (Harun et al., Reference Harun, Dambul, Abdullah and Mohamed2014), teeming with abundant fish, snails and crustaceans. This ecosystem serves as an abundant food source for generalist species like civets, whose activity ranges coincide with those of humans and domestic animals (Wells et al., Reference Wells, Gibson, Clark, Ribas, Morand and McCallum2018). The overlapping presence of definitive and intermediate hosts, combined with the unique characteristics of the floodplain, plays a significant role in facilitating the persistence of multi-host parasites (Anh et al., Reference Anh, Phuong, Murrell, Johansen, Dalsgaard, Thu, Chi and Thamsborg2009). Despite the significant public health importance of A. malayanum and A. sufrartyfex, there is limited information available in the literature. Moreover, since the symptoms of echinostomiasis due to Artyfechinostomum spp. tend to appear long after these parasites have established themselves in the host and are similar to those of other diarrhoeal diseases, medical diagnosis is particularly challenging (Ghatani et al., Reference Ghatani, Arya, Tandon, Yadav, Tandon and Hoti2018).
Although there are obvious links between extensive/industrial oil palm agriculture, riverine/estuarine ecology, and food safety, little attention has been given to the potential threat that oil palm agriculture poses to public health through changes in parasite ecology and food safety. Swamp aquatic ecosystems and flooded areas, prevalent in oil palm-dominated landscapes, can sustain habitats and reservoirs ideal for the development of invertebrate populations that can act as intermediate hosts for trematodes. Therefore, the negative impact of extensive agriculture on the health of aquatic ecosystems and food safety needs to be considered and addressed, as it may have significant implications for the development of effective control strategies in the region.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S0031182023001257.
Data availability statement
Data available within the article or its supplementary materials.
Acknowledgements
Permits to access natural resources were reviewed and granted by Sabah Wildlife Department and the Sabah Biodiversity Centre (JKM/MBS.1000-2/2 JLD.8 [110]; JKM/MBS.1000-2/2 JLD.10 [57,59]). We thank plantation managers for granting us access to plantation estates. Graphic abstract was created using BioRender.
Author contributions
LF and SGS conceived and designed the study. LF and SGS conducted data gathering. LF, BOM and THC conducted the analyses. SS provided support throughout the application process for permits. LF and SGS wrote the original draft. BOM, THC and SS reviewed and edited the original draft.
Financial support
This research was supported by a Nanyang Technological University Presidential Postdoctoral Fellowship (#020617-00001) to LF.
Competing interests
None.
Ethical standards
Not applicable.





 Open access
Open access