


Introduction
Animal traditions are behavioural strategies that can allow species to adapt to changing conditions or better utilize their environment (Bonner, Reference Bonner1980; Fragaszy & Perry, Reference Fragaszy and Perry2003). Traditions begin as innovations by one or a few individuals and are then inherited through social learning mechanisms (Fragaszy & Perry, Reference Fragaszy and Perry2003). Traditions are not species-specific patterns of behaviour; rather, their occurrence is generally limited to a few, isolated subpopulations, making their distribution fragmented and their persistence highly vulnerable to human impact (van Schaik, Reference van Schaik2002).
Burmese long-tailed macaques Macaca fascicularis aurea maintain stone-hammering traditions along the coasts and mangroves of Thailand and Myanmar (Carpenter, Reference Carpenter1887; Malaivijitnond et al., Reference Malaivijitnond, Lekprayoon, Tandavanittj, Panha, Cheewatham and Hamada2007). Their tool use is primarily a coastal foraging adaptation, as tool use is not known to occur away from coastal habitats. Consequently, conserved coastal areas are a prerequisite for maintaining these rare macaque tool traditions. Macaca fascicularis fascicularis, which is widely distributed in Indonesia and Malaysia, is categorized as Least Concern on the IUCN Red List but M. fascicularis aurea is categorized as Data Deficient (Ong & Richardson, Reference Ong and Richardson2008).
Stone-using macaques are rare. They have a limited distribution in Thailand, in only seven locations (Malaivijitnond et al., Reference Malaivijitnond, Lekprayoon, Tandavanittj, Panha, Cheewatham and Hamada2007; YouTube, 2008; Gumert et al., Reference Gumert, Kluck and Malaivijitnond2009, Reference Gumert, Low, Tan and Malaivijitnond2010). The most observable tool use occurs at Piak Nam Yai Island, in Laem Son National Park, Ranong, and Koram Island in Khao Sam Roi Yot National Park, Prachuab Khiri Khan. Koram is highly disturbed by tourist activity but Piak Nam Yai is more pristine and less disturbed. In Myanmar stone use was reported > 120 years ago (Carpenter, Reference Carpenter1887) but a nationwide survey in 2004–2009 did not report any stone-using populations (San & Hamada, Reference San, Hamada, Gumert, Fuentes and Jones-Engel2011). However, during this survey there was no specific intent to look for tool use. In 2011 a local guide in Kawthaung claimed the behaviour still occurred in the southern parts of the Mergui Archipelago (pers. comm. to MDG), suggesting further investigation is needed.
The stone-using habits of Burmese long-tailed macaques make them part of a rare class of extant primates that exhibit customary stone-hammering traditions, which also includes western chimpanzees Pan troglodytes verus in West Africa and bearded capuchins Cebus libidinosus in Brazil (Haslam et al., Reference Haslam, Hernandez-Aguilar, Ling, Carvalho, de la Torre and Destefano2009). Other than these few primates only sea otters Enhydra lutris regularly use stone tools, although various other species of primates and birds use stone tools intermittently (Shumaker et al., Reference Shumaker, Walkup and Beck2011). Animal populations that exhibit stone tool-use are rare, and consequently these traditions are vulnerable to being extinguished by human activity.
Much of the wildlife in South-east Asia is experiencing pressure caused by rapid development and high rates of deforestation (Achard et al., Reference Achard, Eva, Stibig, Mayaux, Gallego, Richards and Malingreau2002). Forest loss has led to a risk of extinction for a large proportion of the region's biodiversity (Sodhi et al., Reference Sodhi, Koh, Brook and Ng2004) and is also a major driver of increasing human–wildlife conflict (Woodroffe et al., Reference Woodroffe, Thirgood and Rabinowitz2005). Conflict with wildlife in South-east Asia now involves numerous species, including elephants Elephas maximus, tigers Panthera tigris (Nyhus & Tilson, Reference Nyhus and Tilson2004), pigs Sus scrofa (Linkie et al., Reference Linkie, Dinata, Nofrianto and Leader-Williams2007), sun bears Helarctos malayanus (Fredriksson, Reference Fredriksson2005) and several primate species (particularly Macaca spp.; Linkie et al., Reference Linkie, Dinata, Nofrianto and Leader-Williams2007; Riley, Reference Riley2007; Gumert, Reference Gumert, Gumert, Fuentes and Jones-Engel2011).
Human and long-tailed macaque populations overlap substantially and this may be the most widespread human–wildlife conflict occurring in South-east Asia (Eudey, Reference Eudey2008; Gumert, Reference Gumert, Gumert, Fuentes and Jones-Engel2011). Human–macaque conflict is causing a reduction in long-tailed macaque populations through an increase in programmes to reduce their numbers (e.g. sterilization and culling) and systematic wild capture to collect live animals for trade (Eudey, Reference Eudey2008; Hasan & Feeroz, Reference Hasan and Feeroz2010; Gumert, Reference Gumert, Gumert, Fuentes and Jones-Engel2011). Control programmes are the result of increasing public complaints about macaques as pests, which are ultimately the result of an increase in degree of space shared between humans and macaques (Sha et al., Reference Sha, Gumert, Lee, Fuentes, Rajathurai, Chan and Jones-Engel2009). The activities of humans (Gumert et al., Reference Gumert, Fuentes, Engel, Jones-Engel, Gumert, Fuentes and Jones-Engel2011) and their domesticated animals, such as dogs Canis familiaris (Anderson, Reference Anderson1986), put pressure on long-tailed macaques, altering their survivorship and behaviour; this is particularly problematic for small populations (Afendi et al., Reference Afendi, Rachmawan, Gumert, Gumert, Fuentes and Jones-Engel2011). Furthermore, anthropogenic impacts alter the species’ ecology, increasing reliance on anthropogenic food resources, and some individuals have become completely separated from natural habitats (Fuentes et al., Reference Fuentes, Southern, Suaryana and Patterson2005; Malaivijitnond et al., Reference Malaivijitnond, Vasquez, Hamada, Gumert, Fuentes and Jones-Engel2011). Such adjustments to anthropogenic influence can change the wild foraging strategies of macaques, and this could potentially extinguish naturally occurring traditions such as tool use.
The tool-using long-tailed macaques in Laem Son National Park are facing the potential effects of increasing human activities. We need to understand the degree of overlap of these macaques with humans and their domestic animals, how this affects their survival, and whether it will impact their rare stone-using traditions. We have observed long-tailed macaques on Piak Nam Yai since 2005 and conducted a detailed census in 2011. We identified their grouping patterns, distribution, and population size, and observed anthropogenic impacts on the island and assessed how these factors are affecting the macaques.
Study area
Laem Son National Park is a marine national park on the western seaboard of Thailand. It protects parts of the Andaman Sea and surrounding bays, islands, and coastal areas (Fig. 1). The park was established in 1983 and covers 315 km2, of which 267 km2 is open sea (Faculty of Forestry, 1987). The Park protects 60 km of coastline and 15 islands, with 14 km2 of its area being mangrove forest comprising predominantly Rhizophora apiculata (DNP, 2006). These mangroves are of high conservation concern and part of a protection programme under the Ramsar Convention (Royal Forest Department, 2002). Other habitats include sandy beaches, mud flats, rocky shores, tropical rainforest and coastal Casuarina equisetifolia forests (DNP, 2006).

Fig. 1 Laem Son National Park (LSNP) is a marine national park along the western coast of Thailand. The study site at Piak Nam Yai Island is in black. The arrow on the inset indicates the Park's location in Thailand.
Of the 15 islands in the Park, long-tailed macaques inhabit only Piak Nam Yai and Thao Islands. The macaques on both of these islands use stone tools, are completely wild, and flee from approaching humans (Malaivijitnond et al., Reference Malaivijitnond, Lekprayoon, Tandavanittj, Panha, Cheewatham and Hamada2007). We chose Piak Nam Yai for our study because the macaques and their tool use are more easily observable there.
Piak Nam Yai is a small island with an area of 1.7 km2 and 5.4 km of coast (Fig. 2). The island contains mountainous tropical forest interlaced with freshwater streams. The coast is predominantly rocky shore but there is a large mangrove area and a small stretch of sandy beach. Between the mangrove and beach is a lowland grass area intermixed with sparse Casuarina groves.

Fig. 2 Piak Nam Yai, Piak Nam Noi, and Thao Islands lie within 1.5 km of the mainland. The thick land border indicates shores above sea level and the thin intertidal border indicates intertidal mangroves and sandy shores. The home ranges of the nine macaque groups observed (see 2-letter codes in Table 1) are indicated, with the developing farms and infrastructure, and boat locations when dogs were sighted on the coast.
In December 2004 the Indian Ocean tsunami that struck Banda Aceh, Phuket, Phang Nga and other regions of Asia, India and Africa, also struck Laem Son National Park and the surrounding villages (USAID, 2005; Kendall et al., Reference Kendall, Aryuthaka, Jittannon and Pateson2010). Based on local reports no one in the nearest village, Bang Ben, was killed, but most structures were demolished and numerous livestock were lost. Reconstruction occurred quickly, supported by international and Thai charities (USAID, 2005). During reconstruction in 2006 a bridge was built that leads to a pier on Piak Nam Noi Island, a small island 100 m from Piak Nam Yai (Fig. 2). Laem Son National Park does not restrict access to this new bridge and pier, labelling it a free access area, and thus it has become a point of free entry into the Park for the local community and visitors. This has increased access to and use of the Park.
Methods
Surveys were conducted by boat around Piak Nam Yai between 15 January and 9 June 2011. We conducted surveys on 89 days, for an average of 5 hours per day. Each survey was tracked using a global positioning system (GPS), which collected positional data every 30 seconds. Circuits of the island were always completed so that every part of the coast was surveyed.
We circumnavigated the island until macaques were sighted, and then stopped and noted the position of the boat. If visible, the macaques were observed from a distance of 10–60 m, and counted. In addition, we identified individuals using observation and photography. A macaque was considered identified after unambiguous recognition of an individual by the researcher over several days. We also recorded whether each individual was ever observed using a tool.
Identifications were used to determine groups and to count individuals. The total population was determined using the counts of identified individuals, plus estimated counts of the few unidentified individuals, which were based on head counts. The coastal range of each group was determined from the GPS data. We calculated the proportion of the population that used tools, and tested if group size was related to the length of coastal range used by groups and number of times sighted, using the Spearman rank correlation coefficient (ρ). Statistics were calculated with SPSS v. 19 (SPSS Inc., Chicago, USA).
Our census method assumed that all macaques living on Piak Nam Yai must regularly come to the coast. This is a reasonable assumption. Firstly, long-tailed macaques are an edge species and reliably come to forest edges (Gumert, Reference Gumert, Gumert, Fuentes and Jones-Engel2011). Secondly, macaque home ranges can be equivalent to the entire area of Piak Nam Yai (Fooden, Reference Fooden1995), and thus it is unlikely a group would remain only in the interior forest. Lastly, on a small island the coast probably plays a highly important role in the macaques’ diet (Gumert et al., Reference Gumert, Kluck and Malaivijitnond2009).
We recorded any types of anthropogenic impact observed on Piak Nam Yai during this study and during shorter studies conducted since 2005 (29 January–2 February, 23–25 March 2005; 1–5 December 2007; 27 February–1 March, 3–23 June, 10–20 December 2008; 5–21 December 2009; 7–12 December 2011). We quantified aspects of this impact. Firstly, all groups were categorized as either having high or low overlap with human activity; high overlap was defined as > 20% of the group's range overlapping with farm plots and forest clearing. Secondly, we recorded the location of dogs, with a GPS, and recorded any dog–macaque interactions. We measured the degree to which a macaque group overlapped with dog activity by calculating a dog intensity score, using: DI gr = (D gr/R gr) × 100, where D is number of dog sightings in the range of a group, R is length of coastal range of group, gr is group, and DI is intensity of dog activity. This score provided a measure of dog activity that accounted for the effect of range size on the likelihood of dog–macaque encounters.
We studied the effects of anthropogenic impact on group composition and coastal usage by comparing groups that had low and high overlap with human and dog activity. We also tested the relationship between group composition and differing levels of dog activity. We used Mann–Whitney U tests for the comparisons and Spearman's rank correlation coefficient to test the relationship.
Results
Population census
We made 248 circumnavigations of Piak Nam Yai over 89 days, averaging 2.79 (range 1–5) circuits per day. We sighted macaques on the coast 462 times (Table 1), averaging 1.86 sightings per circuit (range 0–6 groups per circuit). We found that macaque sightings were almost continuously distributed along the coastal region of Piak Nam Yai (Fig. 2) except for the sandy beach and northern mangrove. Both of these zones are open areas on opposite coasts (i.e. east and west), adjacent to farming plots where people frequently work (Fig. 2). Macaques had been observed in these zones in 2007 and 2008 (MDG pers. obs.), when human activity was less frequent.
Table 1 Population data for each of the nine groups of Burmese long-tailed macaques Macaca fascicularis aurea at the end of the census in June 2011, with the number of sightings of each group, total count of individuals, number of individuals identified, group composition, percentage composition of young macaques (juveniles + infants), length of coastal range, number of sightings of dogs within each group's range, and the dog intensity score within a group's range (see text for further details). Shaded rows show groups with > 20% of their range overlapping farmland.
All adult and adolescent animals that came to the coastal areas were identified and logged. In June 2011 there were 143 identified individuals (52 adult females, 25 adult males, 12 adolescent females, 18 adolescent males, and 36 juveniles), plus a head count of 24 juveniles and 25 infants that were not individually recognizable, bringing the total population count to 192 individuals (Table 1). Observations of tool use indicated that at least 88% of the adults and adolescents, 58% of the population, used tools. The latter figure could be an underestimate because we could not reliably count tool use in unidentified juveniles.
We identified nine groups (Fig. 2). They were highly variable in size (8–35 individuals), in the length of coastline they utilized (500–1,350 m), and in the number of times we sighted them on the coast (14–116; Table 1). The range of coastline a group used was highly correlated with group size (n = 9, ρ = 0.803, P = 0.009). There was a potentially positive but non-signficant relationship between group size and the number of times a group was sighted on the coast (n = 9, ρ = 0.580, P = 0.102).
Anthropogenic impacts
After the 2004 tsunami, development occurred in many areas of Laem Son National Park, including Piak Nam Yai. Although no people yet live permanently on Piak Nam Yai, we observed an expansion of anthropogenic impact. The most significant impact has been land conversion to rubber tree Hevea brasiliensis, oil palm Elaeis guineensis, and coconut Cocos nucifera plantations on the northern part of the island (Fig. 2). The rubber and oil palm farms are particularly damaging because forest and hillsides are clear-cut to plant saplings. A small day camp for plantation workers was built (Fig. 2), which consists of small houses, bungalows, a water well, and fencing. In addition, a variety of vegetable crops have been planted around the settlement.
There are several other human impacts on Piak Nam Yai. Domestic dogs were observed on the coasts, mainly near the plantations (Fig. 2). Several hundred metres of water piping were left along the eastern shores and mangroves after a failed attempt to extract fresh water from the streams during post-tsunami reconstruction. We observed collection of oysters, clams and cockles along the shores and mudflats, and high levels of net fishing around the shores. In the forests we observed opportunistic logging, metal binding of fruit trees (e.g. bitter bean Parkia speciosa) to prevent wild animals ascending, and the redirection of streams using canals. On three occasions we also found dead or injured macaques near the plantations.
To assess if human activity was affecting macaque mortality we compared the composition of juveniles and infants across groups with respect to high (n = 4 groups) and low (n = 5) overlap with the farms (Table 1). High-overlap groups had significantly fewer juveniles and infants (M = 0.33, SD = 0.14, range 0.13–0.43) than groups with low overlap (M = 0.51, SD = 0.02, range 0.50–0.54; Mann–Whitney U Test: U = 0.00, Z = −2.460, P = 0.012; Fig. 3). High-overlap groups were potentially sighted less often on the coasts (M = 31.75 times, SD = 18.25, range 14–57) than low-overlap groups (M = 67.00 times, SD = 35.71, range 25–116) but the difference was not significant (Mann–Whitney U Test: U = 3.5, Z = −1.599, P = 0.110; Fig. 3).
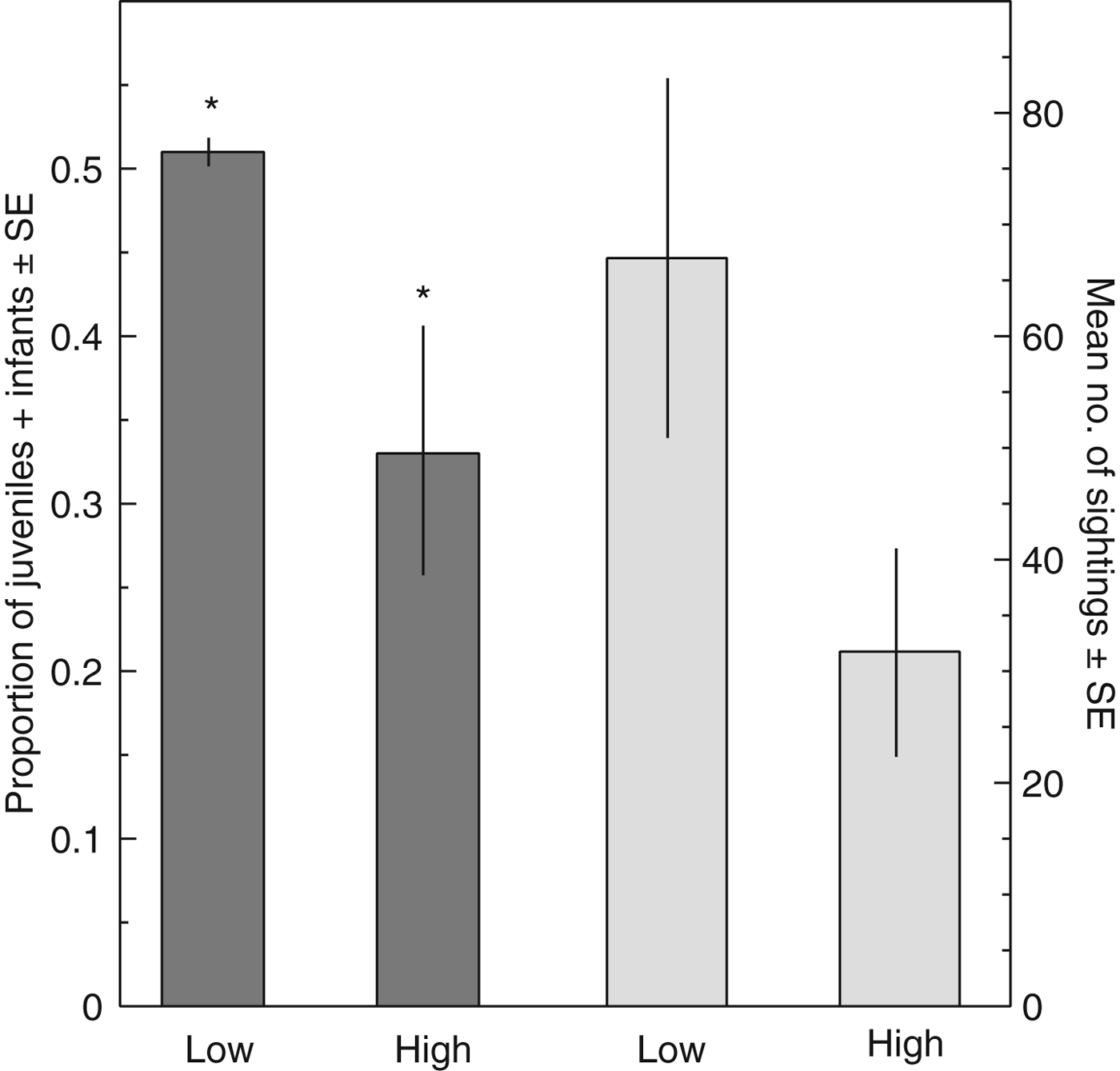
Fig. 3 Differences between groups with low and high overlap with farms in the proportion of juveniles + infants and number of sightings on the coast. Bars marked with an asterisk (*) are significantly different from each other.
The most obvious daily impact on the macaques was from six identified domestic dogs that we observed a total of 47 times on the shores (Fig. 2). We observed 15 dog–macaque interactions. The dogs chased macaques 13 times, passively displaced them once, and on one occasion the macaques did not react as the dogs sat resting. The dogs thus displaced macaques in 14 out of 15 (93.3%) interactions, causing them to flee and alarm call, indicating that dogs are a major deterrent for macaques using the coast.
The largest number (n = 27) of dog sightings occurred in the range of the northernmost group, BC (Fig. 2, Table 1). We first began to regularly observe dogs in the BC range in 2009 and since then we have not observed juvenile (age 1–3 years) macaques in the group. We observed at least six juveniles in the group in December 2007 and seven juveniles and three infants (age 0–1 year) in December 2008. In December 2009 we found no infants or juveniles. In November 2010, one infant was observed but, in January 2011, it had not survived, and no juveniles or infants were observed. During our census, three infants were born into the group and one disappeared at 8 weeks of age. The other two were still surviving at the end of the census in June 2011 but were absent in December 2011.
To assess the possible impact of hunting dogs on juvenile mortality we analysed how the degree of overlap with dogs related to group composition. We calculated the intensity of dog activity in each group's range (Table 1), and then correlated each group's dog-intensity measure with juvenile–infant composition. We found a significant negative correlation between intensity of dog activity and juvenile–infant composition (n = 9, ρ = −0.698, P < 0.037; Fig. 4). Although we never witnessed a dog killing a macaque, the data on group composition shows that where dog–macaque overlap was highest the proportion of young macaques was much lower, suggesting predation.
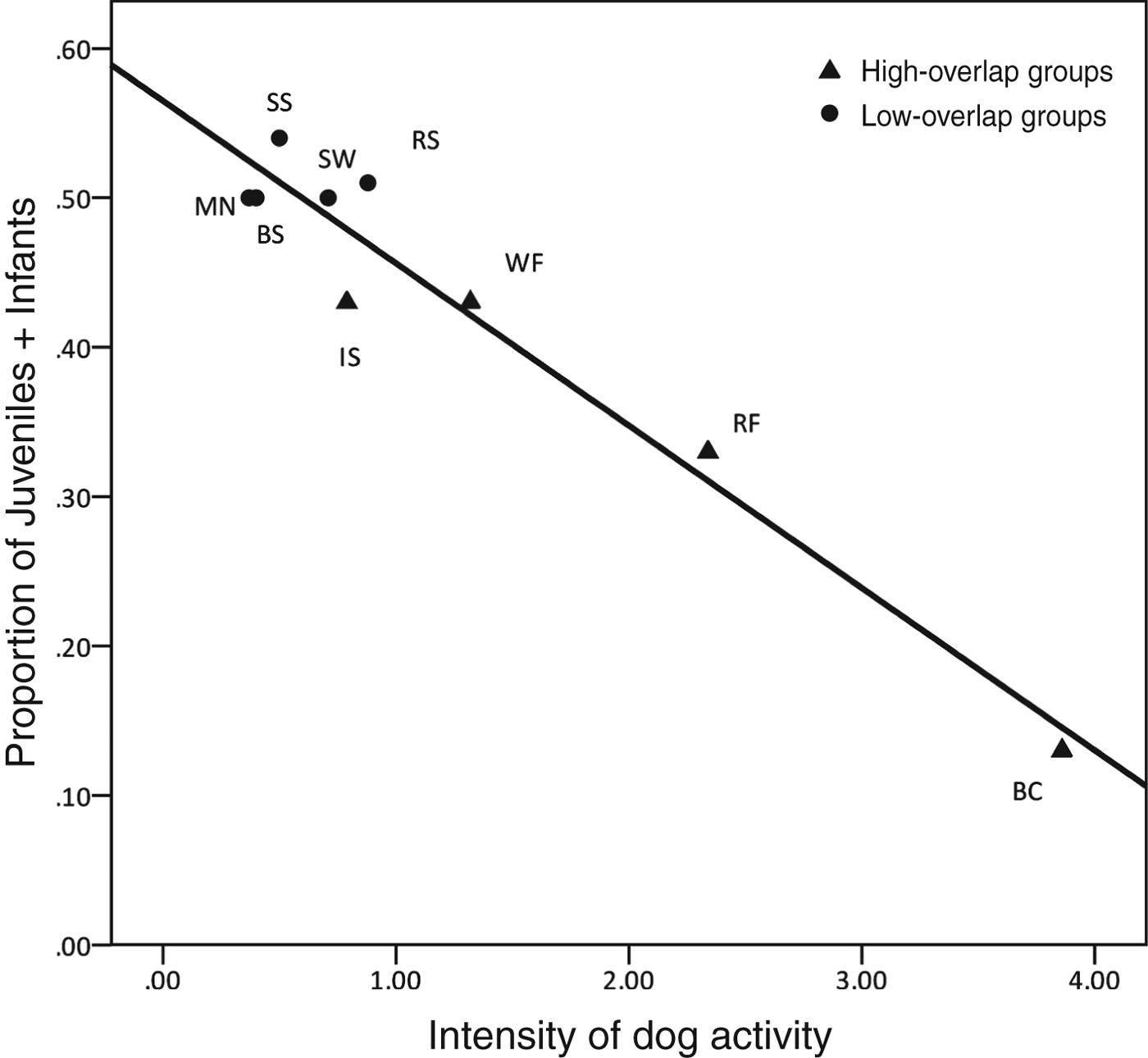
Fig. 4 The relationship between a group's juvenile–infant composition and the intensity of dog activity (for details of dog intensity score, see text) within their range. The two-lettered codes for each group are as in Table 1.
Discussion
Piak Nam Yai is a protected island in Thailand's marine national park system. It is unique because it contains the most conspicuous and regularly observable stone-tool use of any currently known wild long-tailed macaque population. There are many areas on the mainland coast near Piak Nam Yai with a similar ecology but we have not observed extensive coastal use or regular tool-based foraging by macaques there. These mainland coastal areas are surrounded by farms and roads and have been affected by regular activities of people and their domesticated animals (i.e. dogs and buffalo Bubalus bubalis). These anthropogenic influences may inhibit usage of these areas by macaques. In our study we found that human activity negatively affects tool-using macaques by limiting their usage of the coast and the proportion of young in their groups. If these impacts continue the macaque population on Piak Nam Yai could decrease, be displaced from the shores, and have their foraging strategies altered, all of which could disrupt the persistence of their stone-using tradition.
Significant anthropogenic changes in an animal's environment can alter its dietary habits, foraging strategies, and movement (Sutherland, Reference Sutherland1998; Ditchkoff et al., Reference Ditchkoff, Saalfeld and Gibson2006), and the introduction of dogs can affect habitat use through harassment and predation (Anderson, Reference Anderson1986; de Oliveira et al., Reference de Oliveira, Linares, Corrêa and Chiarello2008). On Piak Nam Yai human impact is altering the island's ecosystems and the foods available to macaques. Development for farmland is clearing large portions of forest, harvesting of bivalves is reducing the amount available to macaques, and dogs are repelling macaques from the shores and may prey on vulnerable young. If these changes continue the macaques could alter their foraging strategies, possibly limiting further development of their coastal stone-using traditions.
Cercopithecine primates, such as baboons Papio spp. (Maples et al., Reference Maples, Maples, Greenhood and Walek1976; Altmann & Muruthi, Reference Altmann and Muruthi1988; Hill, Reference Hill2000), vervets Chlorocebus spp. (Else, Reference Else and Box1991) and macaques (Southwick & Siddiqi, Reference Southwick and Siddiqi1994; Fuentes et al., Reference Fuentes, Southern, Suaryana and Patterson2005, Reference Fuentes, O'Neill, Shaw and Cortes2007; Riley, Reference Riley2007) are particularly affected by anthropogenic impacts and will significantly change their behaviour where they are affected. Their ability to adjust appears adaptive because after shifting from wild foraging patterns to synanthropy (i.e. tolerance and dependence on humans), populations of savannah baboons Papio cynocephalus (Altmann & Muruthi, Reference Altmann and Muruthi1988), rhesus macaques Macaca mulatta (Southwick & Siddiqi, Reference Southwick, Siddiqi, Prince Ranier and Bourne1977) and long-tailed macaques (Brotcorne et al., Reference Brotcorne, Wandia, Rompis, Soma, Suartha, Hunyen, Gumert, Fuentes and Jones-Engel2011) regularly rebound with population increases. This flexibility allows them to avoid local extinction and thrive in human-altered environments but at the cost of losing their wild behaviour.
Traditions are particularly vulnerable to environmental disturbances. The ‘fragility of traditions’ hypothesis (van Schaik, Reference van Schaik2002) predicts that sustained disruptions from human activity can extinguish animal traditions. The threats we observed at Piak Nam Yai are continuing and have the potential to inhibit the development and cross-generational transmission of stone-use, if not controlled. We found that young macaques have a higher mortality rate around humans and domesticated animals, and this leaves fewer developing individuals to learn traditions. Additionally, harassment by domestic dogs lessens the time macaques spend on the shores, increases their need to be vigilant, and reduces attention to learning complex tool-using patterns.
The human activities affecting macaques in Laem Son National Park are illegal according to Thailand's 1961 National Parks Act. However, ongoing legal arbitration in Thailand has confused the issue of how to regulate use of national park land (ICEM, 2003; USAID, 2011). As a result, use of protected land by local communities is common across Thailand (FAO, 2009). This issue appears to affect how Laem Son National Park is being managed because Park officials have allowed local authorities to make decisions on Park usage, which has resulted in the present conversion of land inside the Park. New management at Laem Son National Park attempted to re-establish protection in June 2011, by posting eviction notices at all plantations on Piak Nam Yai. Shortly thereafter a guard post was erected before the bridge to the free-access pier. However, these efforts were not successful and the development of the island continues.
The macaques of Laem Son National Park are an important representative population of stone-using long-tailed macaques in Thailand. We recommend developing a programme to monitor and sustain the rare stone-using traditions found in some of Thailand's long-tailed macaque populations. A nationwide census needs to be conducted to examine the extent and distribution of stone tool-using macaques. Protected habitats in the coastal regions of Thailand's marine national park system should be managed so that coastal macaques can forage undisturbed. Actions to limit human impacts in protected coastal habitats would not only benefit stone-using macaques but also other species that depend on the threatened coastal ecosystems of Thailand.
Acknowledgements
The Chulalongkorn University Centenary Academic Development Project funded this study, with additional support from the Academic Research Fund of the School of Humanities and Social Sciences at Nanyang Technological University and the Ministry of Education in Singapore (Grant RG07/95). The National Research Council of Thailand provided a Foreign Research Permit to MDG, and the National Park, Wildlife, and Plant Conservation Department of Thailand provided an entry permit for Laem Son National Park. The field methods used in this study were reviewed and approved by the NTU-Institutional Animal Care & Use Committee in Singapore (ref: ARF SBS/NIE-A 0138 AZ). We thank the park management at Laem Son National Park for their assistance, and Haromai Raksa for providing a boat service. We also thank W.C. McGrew,Martin Fisher and an anonymous reviewer for useful help and comments.
Biographical sketches
Michael D. Gumert studies the behaviour and ecology of Macaca fascicularis in Thailand, Singapore and Indonesia. He is interested in their social and cultural behaviour, as well as their relationship with humans and anthropogenic habitats. Yuzuru Hamada studies primate morphology and macaque population ecology across South-east Asia. He recently completed nationwide surveys in Thailand, Laos and Myanmar. Suchinda Malaivijitnond has done surveys of macaques throughout Thailand and discovered the tool-using long-tailed macaque population at Laem Son National Park. She is currently leading development of Thailand's Primate Research Institute. Her research programme involves collaboration with international scientists and the Institute is a national centre for primate field studies in Thailand.





