


The ecology, population dynamics, and conservation status of the two species of clouded leopard (Neofelis spp.) are poorly understood. These are medium-sized felids with allopatric distributions. The mainland clouded leopard N. nebulosa ranges across parts of Nepal, southern China, eastern India and most of mainland South-east Asia. The Sunda clouded leopard N. diardi, only recently distinguished from its mainland relative, is restricted to Sumatra and Borneo (Buckley-Beason et al., Reference Buckley-Beason, Johnson, Nash, Stanyon, Menninger and Driscoll2006).
Lack of knowledge of the most fundamental aspects of a species' ecology and abundance makes it difficult to assess threats and conservation status. Both species of clouded leopard are categorized as Vulnerable on the IUCN Red List (Hearn et al., 2011), although it is unclear how reliable these designations are considering the scarcity of basic population information for the species (Wilting et al., Reference Wilting, Mohamed, Ambu, Lagan, Mannan, Hofer and Sollmann2012). Estimating population density of cryptic, rare, and semi-arboreal animals such as clouded leopards is logistically challenging. Furthermore, many of the more commonly used analytical techniques for calculating density estimates are not rooted in statistical theory (Royle & Young, Reference Royle and Young2008). The use of recently developed spatially-explicit capture–recapture techniques may help circumvent some of these problems (Efford et al., Reference Efford, Borchers, Byrom, Thomson, Cooch and Conroy2009).
Here we present the results of a camera-trapping survey of Sunda clouded leopards in the Maliau Basin Conservation Area, Sabah, Malaysian Borneo, and spatially-explicit capture–recapture models to estimate population density. This 588 km2 Conservation Area encompasses a circular sedimentary basin and a surrounding buffer zone. Evidence of humans in the basin prior to the 1980s is limited (Hazebroek et al., Reference Hazebroek, Adlin and Sinum2004), suggesting that historical human use of the area might have been low. The Maliau Basin contains mixed lowland and hill dipterocarp rainforests and tropical heath forests surrounded by sedimentary mountains. Mean annual rainfall is c. 3,800 mm (Mykura, Reference Mykura and March1989). Many streams drain the basin and converge into the Maliau River. Primary forests in the basin comprise some of the last floristically and faunally intact rainforests (i.e. no logging and no known extinctions of Holocene vertebrates) in Borneo. Outside the basin but within the Conservation Area are stretches of dipterocarp forest that were selectively logged in the early–mid 1990s (J. Naimin, Yayasan Sabah, pers. comm.). Canopy cover in these logged forests is reduced and understorey and ground layers are thick and composed of grasses, forbs and ferns (J. Brodie, pers. obs.). Although nominated as a World Heritage Site in 2003, wildlife in the Conservation Area has been little researched, and to our knowledge this is the first attempt to estimate the population density of any animal species within the Conservation Area.
In January 2010 we established 26 camera-trap stations in the Maliau Basin Conservation Area. Our trapping area included logged forest outside the basin (11 stations), as well as tracts of adjacent, unlogged primary forest on the south-facing flank of the rim of the basin, near the basin's rim, and inside the basin (Fig. 1). Twelve stations were in mixed primary dipterocarp forest and three in heath forest. Stations were established 1–2 km apart, covering c. 75 km2 in total, and were situated on potential travel routes for clouded leopards, including wildlife paths (especially following ridge tops), human trails, old logging roads (Gordon & Stewart, Reference Gordon, Stewart and Meijaard2007) and stream beds. Each station comprised two Reconyx RM45 digital camera traps (Reconyx Inc., Holmen, USA) opposite each other along travel paths, affixed to tree trunks with steel cables and padlocks, ≥ 1 m above the forest floor. All units were active on high sensitivity 24 h day−1 and set to take three photographs in rapid succession after being triggered. No lures or baits were used. All cameras contained 1–2 GB memory cards capable of storing 7,500–15,000 images per card, which were retrieved in mid May 2010. Additional details regarding camera deployment and other species recorded can be found in Brodie & Giordano (Reference Brodie and Giordano2011).
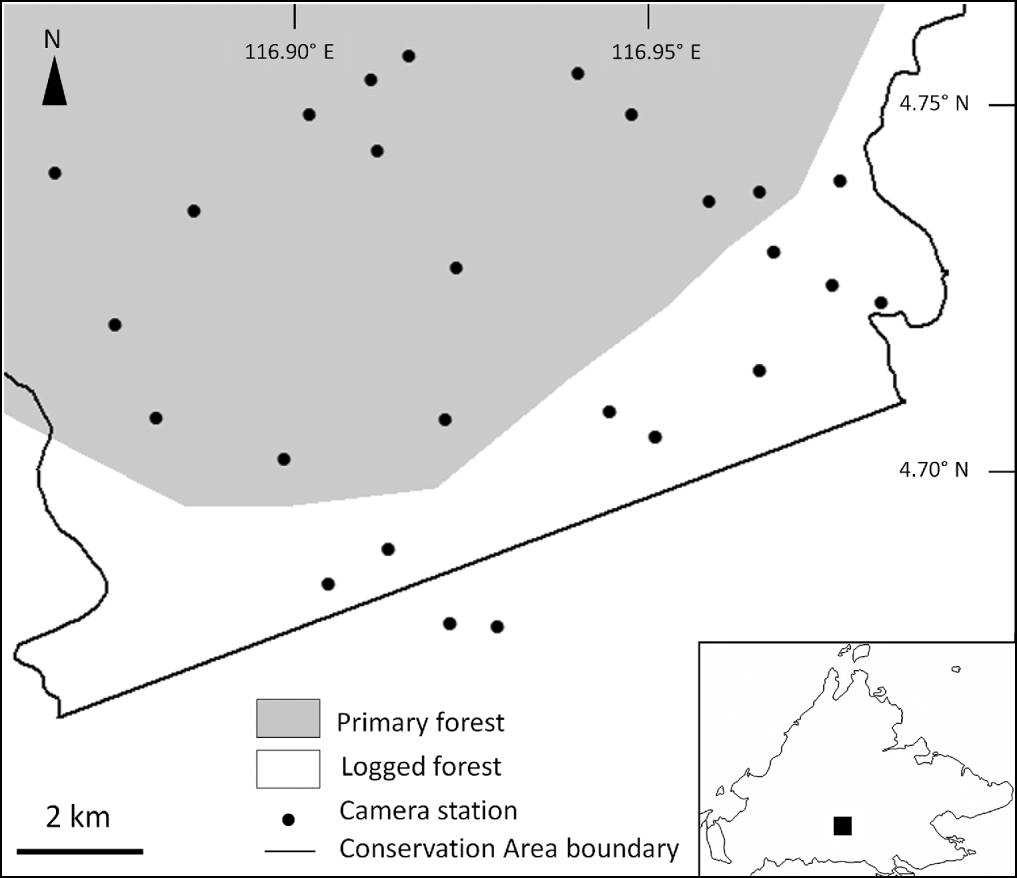
Fig. 1 The location of the camera-trap stations for the Sunda clouded leopard Neofelis diardi in the southern portion of the Maliau Basin Conservation Area. The rectangle on the inset shows the location of the Conservation Area in Sabah, Malaysian Borneo.
Our spatially-explicit capture–recapture approach to estimate clouded leopard population density was based on maximum likelihood (using Density; Efford, Reference Efford2009). To conduct our analysis we chose the 81-day capture window containing the most clouded leopard detections (1 Feb–17 April). Because of the low detection rate we used a longer time window than is generally used for other felids (Karanth & Nichols, Reference Karanth and Nichols2002), to achieve an adequate minimum sample size for analysis. Each sampling occasion consisted of 3-day groupings (days 1–3, 4–6, etc.) so as to increase per-occasion detection rates and potentially obtain parameter estimates with less bias (cf. Otis et al., Reference Otis, Burnham, White and Anderson1978). We employed a multiple live trap layout design based on the Poisson distribution model with a full likelihood setting and half normal detection function (default settings in Density; Efford, Reference Efford2009). Spatially-explicit capture–recapture models estimate three parameters: density (D), g0, and σ; the latter two together define the detection function. Because of the low sample size we tested only simple models in which both detectability (g) and σ were constant.
We obtained a total of 2,003 trap-nights in both primary and previously-logged forest across the 81-day capture window. In primary forest we obtained 59 photographic capture events (i.e. photos that, if taken at the same site, occurred >1 hour apart) of clouded leopards in 1,134 trap-nights, with one station being lost because of flooding. Captures were distributed heterogeneously among four individuals, with 48, 6, 4, and 1 captures. In logged forest we obtained 869 trap-nights in the 81 days, although camera batteries at several stations failed before 17 April. Despite this extensive sampling effort we did not record any photos of clouded leopards in this period; however, we did record one clouded leopard photo outside this interval. We therefore restricted our density analysis to primary forest by masking the logged forest out of the spatial analysis. Using Google Earth (Google Inc., 2009) we partitioned the study area into primary versus logged forest based on the presence of road networks in the latter.
The root pooled spatial variance (RPSV) in the data was 3,102.3 m, so we set the buffer width for initial analyses to 4 RPSV (c. 12,000 m; cf. Efford, Reference Efford2009). The model incorporating a mask of logged forest restricted analysis to primary forest and gave a population density estimate, D, of 0.019 km−2 (95% confidence limits: 0.007–0.054), g0 = 0.098 (0.050–0.182) and σ = 3,347.981 (2,161.841–5,184.922), or c. 1.9 individuals per 100 km2. The model for the entire study area, irrespective of forest type (i.e. without using the habitat mask), gave D = 0.008 km−2 (0.002–0.026), g0 = 0.106 (0.053–0.200), and σ = 3,236.020 (1,910.317.841–5,481.721), or c. 0.8 individuals per 100 km2.
Our study provides one of the first published population density estimates of either species of clouded leopard. Several other studies have estimated clouded leopard density in Sabah based on tracks (Davies & Payne, Reference Davies and Payne1982; Wilting et al., Reference Wilting, Fischer, Abu Bakar and Linsenmair2006) but these techniques are problematic because of the uncertainty in reliably differentiating unique individuals (Karanth, Reference Karanth, Tilson and Seal1987; Karanth et al., Reference Karanth, Nichols, Seidensticker, Dinerstein, Smith and McDougal2003; Gordon et al., Reference Gordon, Stewart and Meijaard2007).
However, our results must be interpreted with appropriate caution. The low precision of our estimates, although not unexpected considering the cryptic nature of the species and its semi-arboreal habits (which together contribute to low detection probabilities with ground-based camera-trapping methods), and the small sampling area and resulting small sample size, will make it difficult to detect population trends in the Maliau Basin or differences in density between Maliau and other sites. Comparison with the clouded leopard density estimates obtained by Wilting et al. (Reference Wilting, Mohamed, Ambu, Lagan, Mannan, Hofer and Sollmann2012) in two commercial forests in Sabah are also difficult because of the different modelling approaches used. Nevertheless, because the Sunda clouded leopard is categorized as Vulnerable, with a decreasing population trend (Hearn et al., Reference Hearn, Sanderson, Ross, Wilting and Sunarto2011), it warrants future surveys at this and other sites.
Acknowledgements
We thank the Malaysian Economic Planning Unit, Yayasan Sabah, the staff of Maliau Basin Conservation Area, particularly H. Tangki and J. Naiman, and the Sabah Wildlife Department, particularly L. Ambu. Crucial assistance was provided by S.T. Wong, E.A. Razlie and O. Helmy. Financial and in-kind support for the project was provided by Panthera, the Denver Zoological Foundation, the Columbus Zoo, the Clouded Leopard Project, Ideawild, and SPECIES. The article has also been much improved by comments from several anonymous reviewers.
Biographical sketches
Jedediah Brodie and Anthony J. Giordano are interested in the ecology and conservation of vertebrates, particularly medium- to large-sized mammals, and tropical rainforest communities. Jedediah focuses on how anthropogenic stressors such as bushmeat hunting, logging and climate change affect mammal populations and also how they potentially disrupt animal–plant interactions such as seed dispersal. He mainly works in South-east Asia but has also conducted research in Africa, Central America, South Asia and North America. Anthony focuses on non-invasive estimation of population parameters, the effects of habitat conversion and fragmentation on competition and predator–prey dynamics, and trophic interactions in carnivore assemblages. He is involved in conservation and research projects in tropical Latin America, Asia and Africa. Both are on the board of directors for regional sections of the Society for Conservation Biology and are currently Fulbright Fellows based in Malaysian Borneo and Paraguay, respectively.

