



Introduction
Skeletal remains of Late Pleistocene mammals can be found across the Netherlands as they were preserved by fluvial sediments which now surface across the country (Doppert et al., Reference Doppert, Ruegg, Van Staalduinen, Zagwijn and Zandstra1975; Van Kolfschoten & Laban, Reference Van Kolfschoten and Laban1995; De Mulder et al., Reference De Mulder, Geluk, Ritsema, Westerhoff and Wong2003; Gouw & Erkens, Reference Gouw and Erkens2007; Laban & Van der Meer, Reference Laban and Van der Meer2011; DINOloket, 2020). At dredging sites, large numbers of such fossils have been collected and subsequently published (Mol et al., Reference Mol, Van den Bergh and De Vos1999; Van Kolfschoten, Reference Van Kolfschoten2001; Van der Jagt, Reference Van der Jagt2005; Van Kolfschoten et al., Reference Van Kolfschoten, Van der Jagt, Beeren, Argiti, Van der Leije, Van Essen, Busschers, Stoel and Van der Plicht2011).
Thousands of specimens have been collected during dredging operations at Lent near Nijmegen (51°52'38.6"N, 5°51'57.5"E; Fig. 1A). In total, 909 bones in this collection were considered suitable for identification. The faunal remains comprise 732 Holocene and 177 Late Pleistocene mammals. The Late Pleistocene fauna is analysed, with particular attention paid to fly puparia found embedded in skull fragments of the woolly mammoth, Mammuthus primigenius (Blumenbach, Reference Blumenbach1799), contained in the collections of the Oertijdmuseum at Boxtel (MAB10673).
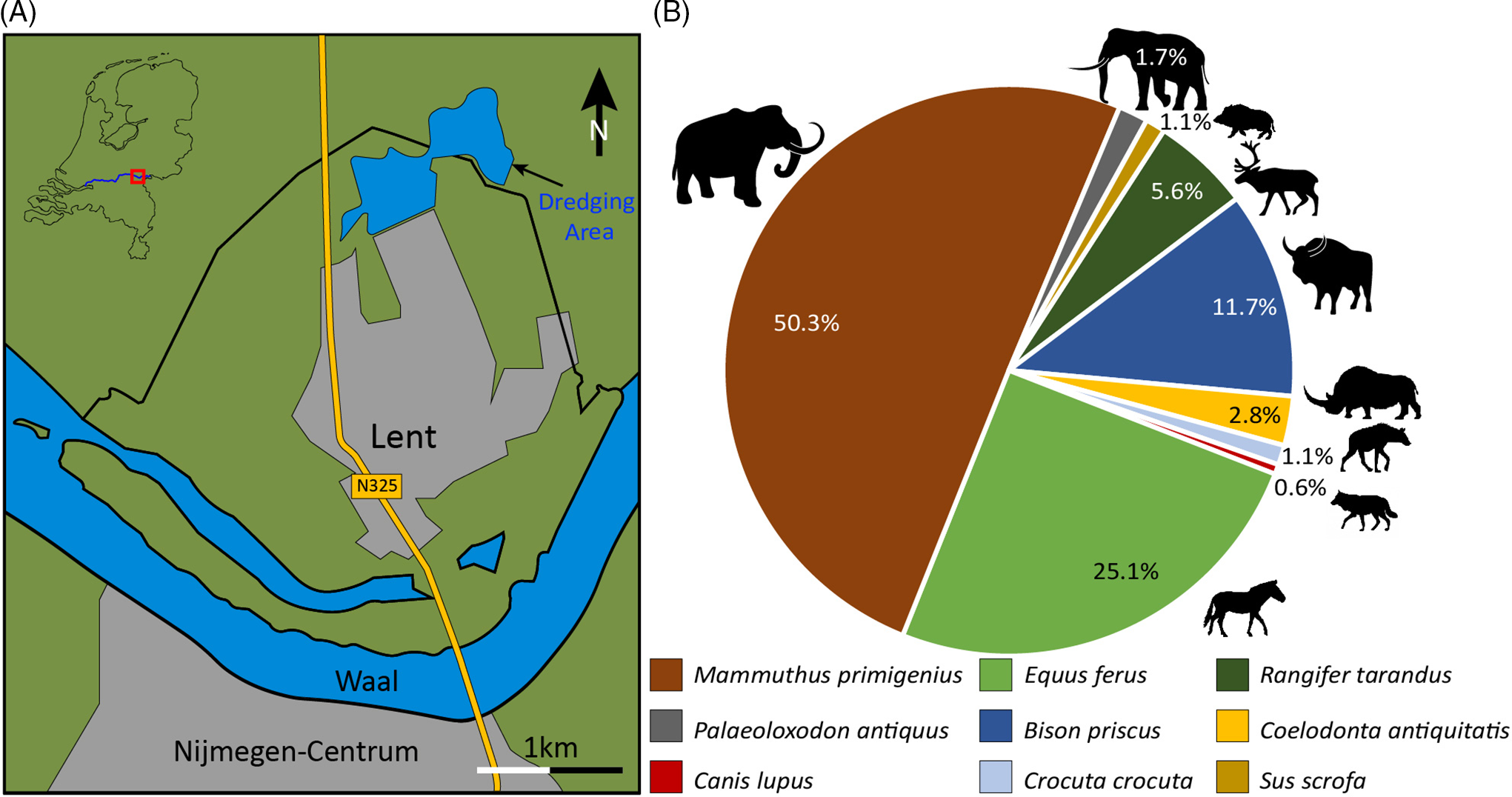
Figure 1. Location of the dredging site and the abundance of Pleistocene megafauna fossils.
Late Pleistocene faunal abundance
One of the most common mammals found in Late Pleistocene deposits throughout the Netherlands is the woolly mammoth (Mol et al., Reference Mol, Van den Bergh and De Vos1999), and Lent is no exception (Fig. 2F further below). Mammoth remains make up 50.3% of the Late Pleistocene assemblage from this locality (Fig. 1B) and are represented mainly as fragmentary skulls, mandibles and pelvises. This high percentage is due in part to their colossal size, which increases the chances of preservation. Additionally, this taxon was abundant and widely distributed during the Late Pleistocene, having been well adapted to grazing and cold temperatures.

Figure 2. Late Pleistocene fossils found at the Lent site: A. dextral radius of a cave hyena (Crocuta crocuta spelaea); B. dextral humerus of a cave hyena; C. atlas of a wild boar (Sus scrofa); D. pelvic fragment of a wolf (Canis lupus lupus); E. dextral lower M3 fragment of a straight-tusked elephant (Palaeoloxodon antiquus); and F. sinistral upper M1 molar of a woolly mammoth (Mammuthus primigenius). Black bar represents 5 cm.
Wild horse Equus ferus Boddaert, Reference Boddaert1785 is also common (25.1%) at Lent, followed by steppe bison Bison priscus Bojanus, Reference Bojanus1827 (11.7%) and reindeer Rangifer tarandus (Linnaeus, Reference Linnaeus1758) (5.6%). Horse remains are abundant, likely because their ecological niche was relatively large compared to that of reindeer or steppe bison. Horses were able to cope with open steppe conditions as well as with more forested environments, while bison and reindeer were restricted to open steppe environments. This flexibility explains why horses are represented not only in interstadial and stadial faunas, but possibly even in interglacial ones (Von Koenigswald, Reference Von Koenigswald2003; Saarinen et al., Reference Saarinen, Eronen, Fortelius, Sepp and Lister2016). Furthermore, reindeer material is less likely to be preserved compared to other herbivores in the present collection, as their thin-walled humeri and radii were easily eroded or gnawed away by predators and scavengers (Van der Jagt, Reference Van der Jagt2005; Zimov et al., Reference Zimov, Zimov, Tikhonov and Chapin2012).
This collection contains only a few remains of the woolly rhinoceros, Coelodonta antiquitatis (Blumenbach, Reference Blumenbach1799), accounting for 2.8%. This low percentage likely represents the true number of individuals of this species that lived during the Late Pleistocene. Woolly rhinos are thought to have been solitary, with males having territories similar to extant forms (Garutt, Reference Garutt1999; Diedrich, Reference Diedrich2008; Becker et al., Reference Becker, Dini and Scherler2014). By contrast, mammoths, horses, steppe bison and reindeer lived in herds, and thus had considerably higher population densities compared to the woolly rhinoceros.
The geology at the site suggests these herbivores are to be associated with cold climatic conditions during the Late Pleistocene, and in particular with the Weichselian Glacial (DINOloket, 2020). They were occupants of the ‘mammoth steppe’, the cold, dry grass steppe that extended over much of the Northern Hemisphere during the Late Pleistocene (Zimov et al., Reference Zimov, Zimov, Tikhonov and Chapin2012).
Predators are not well represented in this assemblage. Of the three bones recovered, two belong to the cave hyena (Crocuta crocuta spelaea Goldfuss, Reference Goldfuss1823; Fig. 2A–B). The third is a heavily mineralised pelvic fragment of a wolf (Canis lupus, Linnaeus, Reference Linnaeus1758; Fig. 2D). Fossils of cave hyenas are not particularly rare in the Netherlands (Reumer et al., Reference Reumer, Mol and Borst2010), while the absence of skeletal remains of other predators suggests that these were too rare in the ecosystem for them to be represented in this assemblage, as predators often are. It is difficult to connect the presence of predators with a particular climatic condition as they can be found both in warm and cold periods of the Late Pleistocene (Von Koenigswald, Reference Von Koenigswald2003). Other predators missing from the present assemblage are notably cave lions, brown bears and cave bears (cf. Bocherens, Reference Bocherens2015).
Species associated with warmer climatic conditions are few in the assemblage. Three molars of the straight-tusked elephant Palaeoloxodon antiquus (Falconer & Cautley, Reference Falconer and Cautley1846; Fig. 2E) and two heavily mineralised bones of wild boar (Sus scrofa Linnaeus, Reference Linnaeus1758; Fig. 2C) have been identified. The geology at the dredging site suggests that the remains of these animals date back to the Eemian and are possibly reworked (Laban & Van der Meer, Reference Laban and Van der Meer2011; DINOloket, 2020).
Fly puparia
In Pleistocene mammalian bones, blowfly puparia, typically the species Protophormia terraenovae Robineau-Desvoidy, Reference Robineau-Desvoidy1830, are occasionally found (Gautier & Schumann, Reference Gautier and Schumann1973; Vervoenen, Reference Vervoenen1991; Gautier, Reference Gautier1995; Verhagen & Mol, Reference Verhagen and Mol2009; Van der Plicht et al., Reference Van der Plicht, Post and Mol2012; Mähler et al., Reference Mähler, Wappler, Sanmugaraja, Menger and von Koenigswald2016). This Holarctic species of the Calliphoridae family is absent from the Netherlands at present (Erzinçlioğlu, Reference Erzinçlioğlu2009; Vanin et al., Reference Vanin, Turchetto, Galassi and Cattaneo2009; Verhagen & Mol, Reference Verhagen and Mol2009). This species breeds on the carcasses of vertebrates, on which their larvae feed and pupate (Roux et al., Reference Roux, Gers, Telmon and Legal2006; Erzinçlioğlu, Reference Erzinçlioğlu2009). Until the 1970s, remains of puparia in Pleistocene mammalian fossils were thought to be rare; subsequently, there have been more observed (Gautier & Schumann, Reference Gautier and Schumann1973; Vervoenen, Reference Vervoenen1991; Gautier, Reference Gautier1995; Verhagen & Mol, Reference Verhagen and Mol2009; Van der Plicht et al., Reference Van der Plicht, Post and Mol2012; Mähler et al., Reference Mähler, Wappler, Sanmugaraja, Menger and von Koenigswald2016).
A skull of a woolly mammoth (MAB10673) from Lent comprises 43 separate fragments (e.g. Fig. 2F), 15 of which contained puparia. Only five puparia (MAB11313; Table 1) were deemed to be sufficiently preserved for identification in accordance with criteria presented in the relevant literature (Gautier & Schumann, Reference Gautier and Schumann1973; Erzinçlioğlu, Reference Erzinçlioğlu2009; see Fig. 3). All five have been identified as P. terraenovae on the basis of length and morphology. Two of these have larger widths than is typical of this species (Erzinçlioğlu, Reference Erzinçlioğlu1988), but this is likely due to infilling sediment having flattened the puparia inside the skull fragments.
Table 1. Length, width and shape factor of the Lent puparia, compared with the data from Erzinçlioğlu (Reference Erzinçlioğlu1988)
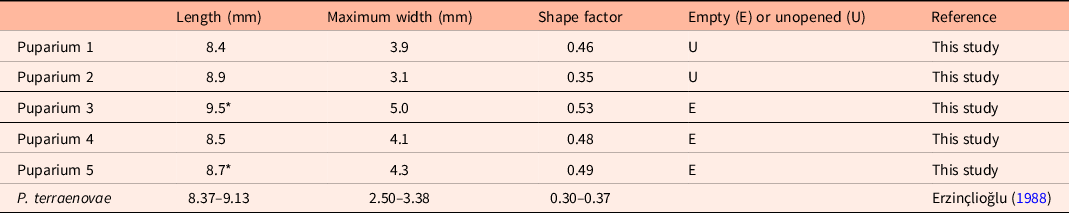
* Estimated length because these puparia were incomplete. Shape factor calculated by dividing the greatest width by overall length.

Figure 3. MAB11313 Puparium 1 (unopened) ventral (A) and dorsal (B) view; Puparium 3 (opened) ventral (C) and dorsal (D) view. Black bar represents 1 mm.
These puparia were recovered from skull fragments close to natural body orifices such as the eyes (zygomatic arch), which are used by blowflies to enter the body, as has been documented for colonisation of human carcasses (Haskell et al., Reference Haskell, Hall, Cervenka, Clark, Haglund and Sorg1997). The larvae likely travelled further into the carcass to feed (Roux et al., Reference Roux, Gers, Telmon and Legal2006).
The mammoth host was likely a subadult bull, as the wear patterns observed on the lower dextral M2 suggest that it had reached the age of 24.5 ± 2 African Equivalent Years (Laws, Reference Laws1966), an estimate based on the procedure outlined by Haynes (Reference Haynes1993) for extinct proboscideans. Sex was determined by comparing the dimensions of the isolated occipital pars basalis to those of the analogous bone in the complete skull of an adult female (MAB11628; Van der Merwe et al., Reference Van der Merwe, Bezuidenhout and Seegers1995; Averianov, Reference Averianov, Shoshani and Tassy1996).
By applying modern forensic methods to the analysis of puparia, one can approximate the minimum temperature at the time of the mammoth’s death (Gilbert & Bass, Reference Gilbert and Bass1967; Vanin et al., Reference Vanin, Turchetto, Galassi and Cattaneo2009). Protophormia terraenovae can develop its eggs only in temperatures higher than 9°C (Nuorteva, Reference Nuorteva1987; Grassberger & Reiter, Reference Grassberger and Reiter2002). Reconstructions by Caspers & Freund (Reference Caspers and Freund2001) suggest that mean July temperatures during the Weichselian ranged from approximately 17°C during the early Weichselian to 3°C during the Late High Glacial stadials. This suggests that this mammoth died either during the early Weichselian or during an interstadial of the High Glacial when temperatures were high enough for egg development.
Three of the five puparia analysed were empty, suggesting that the flies fully developed and left their puparium (Nuorteva, Reference Nuorteva1987). Hence, the carcass was likely exposed to the surface long enough for them to become fully developed. Studies on the periods of development of extant P. terraenovae suggest that full development takes between 35 and 39 days (Marchenko, Reference Marchenko2001; Grassberger & Reiter, Reference Grassberger and Reiter2002). However, this varies according to region and temperature (Grassberger & Reiter, Reference Grassberger and Reiter2002). Other heat sources, such as the carcass itself (Henssge, Reference Henssge1988) and ‘maggot mass heating’, should be taken into account, as they can cause development times to be considerably shorter (Haskell et al., Reference Haskell, Hall, Cervenka, Clark, Haglund and Sorg1997; Erzinçlioğlu, Reference Erzinçlioğlu2009).
Conclusions
Late Pleistocene megafaunal remains from Lent document a considerable preservational bias in favour of mammoths and against species such as reindeer, while accurately representing the abundance of horses, steppe bison and woolly rhinoceros throughout climatic changes during the Late Pleistocene. The presence of Protophormia terraenovae puparia in the woolly mammoth bull skull MAB10763 suggests carcass colonisation occurred during the early Weichselian, or during an interstadial of the High Glacial, during which temperatures met the minimum requirement for egg development.
Acknowledgements
We wish to thank Werner Peters for collecting and donating the Lent material over a period of 3 years, with impeccable consideration for all types of fossil material at the site. Identification of the mammalian material was made possible thanks to Charles Schouwenburg and Dick Mol, while puparia identification was aided by comparative material from Lambert Van Es. Geological interpretations of the area were made possible thanks to Wim Hoek. The senior author thanks Wilma Wessels for supervision throughout this research project. Comments from Wilma Wessels, Charles Schouwenburg, Tom Giltaij, Jan van der Made and John Jagt on early drafts were extremely useful and essential in the writing process. Credit to Ruben Winter for taking the photographs for this project. We thank Dick Mol, Hans van Essen and an anonymous reviewer for pertinent comments and feedback. Special thanks to the MSB zand en grind (Beijer Bouwgrondstoffen, Boskalis and Spaansen collective) for allowing fossil collection at the dredging site with excellent hospitality.
Declaration of interest
The authors declare adherence to Committee on Publication Ethics (COPE) guidelines and have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.





 Open access
Open access