



Anthropological approaches to food decisions and practices cover the spectrum from adaptationist, structuralist, and culturalist points of view. In agrarian societies, dietary choices encompass logics that range from nutritional balance, cost limitations, and political or economic considerations to flavor and complex traditions of food preferences and practices that may or may not align with those practical considerations (Hastorf Reference Hastorf2017). In complex cultural systems, food choices entail political and economic factors constrained by agrarian labor, ecological utility, cost efficiencies, and “prevailing ideologies of food preference” (Smith Reference Smith2006:488–489).
The dialectic between practical considerations and cultural traditions of culinary practice is especially salient in contexts of diaspora involving the migration or translocation of people to new regions. Hastorf (Reference Hastorf2017:253) writes, “Culinary traditions are put to the test when people move”; culinary transformations capture immigrant histories and measure the “level of integrity of a group's identity.” Indeed, in diasporic contexts, the interplay of dietary modification with dietary traditionalism can be an important proxy in considering change and continuity in migrant cultural identities. We explore this topic using data drawn from frontier communities of the Tiwanaku polity, one of the earliest states to develop in the south-central Andes during what is called the Middle Horizon period (around AD 600–1100).
The Tiwanaku civilization originated in the Bolivian altiplano in a unique highland agropastoral niche located 3,800 m asl (Janusek Reference Janusek2008; Kolata Reference Kolata1986, Reference Kolata and Kolata2003; Stanish Reference Stanish2003). At the eponymous type site and in its hinterland, altiplano Tiwanaku people herded llamas and alpacas and grew frost-resistant crops, such as quinoa (Chenopodium quinoa), kiwicha (Amaranthus caudatus), potatoes (Solanum tuberosum), and other tuber crops: oca (Oxalis tuberosa), ulluco (Ullucus tuberosus), and mashua (Tropaeolum tuberosum; Bruno Reference Bruno2008, Reference Bruno2014; Goldstein Reference Goldstein2005; Hastorf et al. Reference Hastorf, Whitehead, Bruno, Wright, Staller, Tykot and Benz2006; Kolata Reference Kolata1986, Reference Kolata and Kolata2003; Lennstrom et al. Reference Lennstrom, Hastorf and Wright1991a, Reference Lennstrom, Hastorf and Wright1991b; Towle Reference Towle1961; Wright et al. Reference Wright, Hastorf, Lennstrom and Kolata2003).
Throughout the Middle Horizon (AD 600–1100), the Tiwanaku civilization expanded into lowland coastal valleys (from around 600–1,500 m asl), such as the Moquegua and Locumba Valleys of Peru (Figure 1), where settlers established large colonies to cultivate temperate-zone crops that could not be grown in the Tiwanaku homeland (Goldstein Reference Goldstein2005). Lowland cultivars include tropical fruits, such as avocado (Persea americana), psychotropic plants like coca (Erythroxylum spp.), cotton (Gossypium spp.), molle pepper (Schinus molle), peanuts (Arachis hypogaea), beans (Phaseolus spp.), sweet potatoes (Ipomoea batatas), chili peppers (Capsicum spp.), and, particularly, maize (Zea mays) (Hernández Bermejo and León Reference Hernández Bermejo and León1994; Towle Reference Towle1961).

Figure 1. Tiwanaku capital (around 3,860 m asl) and coastal-valley Tiwanaku colonial sites, L1 (600 m asl) and M43 (900 m asl) (figure created by Matthew J. Sitek).
In this article, we present a comprehensive analysis of archaeobotanical remains from the Andean Tiwanaku culture's provincial site of Cerro San Antonio (L1) in the Locumba Valley and compare this to data from the contemporary Tiwanaku site of Rio Muerto (M43) in the Moquegua Valley and from the homeland site of Tiwanaku (TIW) in the Bolivian altiplano of the south-central Andes (Figure 1). Highland Tiwanaku settlers occupied the lowland sites of L1 and M43 during the period of state expansion (AD 600–1100). Our goal is to compare the adoption of new agrarian and culinary possibilities in the Tiwanaku diaspora with the persistence of cultivars and preferences from their traditional homeland.
Because of its central importance to Tiwanaku's economic and ritual systems through “vertical complementarity” (Goldstein Reference Goldstein2005; Murra Reference Murra and Murra1972), maize has been the focus of dietary reconstructions of the Middle Horizon Tiwanaku expansion. We posit here that other cultivars are also significant and focus our discussion on the frost-resistant Amaranthaceae cultivars, Chenopodium quinoa (quinoa) and Amaranthus spp. Quinoa was a native and staple food for the Tiwanaku residents of the southern Titicaca Basin (Wright et al. Reference Wright, Hastorf, Lennstrom and Kolata2003). Although traditionally grown at lower elevations (2,000–3,000 m asl) than the altiplano (around 3,800 m asl), Amaranthus spp. would have been recognizable to the Tiwanaku highland people and was likely a “casual food source” to them (Lennstrom et al. Reference Lennstrom, Hastorf and Wright1991a:6, Reference Lennstrom, Hastorf and Wright1991b:6). Given that maize's unique role has captured much of the attention in the discussion of Tiwanaku food investigations, it is helpful to first review the critical position of this lowland crop that bore such inordinate importance in the world's highest-altitude, pristine-state society.
Maize Mania: The Focus on Maize in the Narrative of Tiwanaku Expansion
The Tiwanaku valued maize because it could be brewed into the alcoholic beverage chicha (Biwer and VanDerwarker Reference Biwer and VanDerwarker2015; Goldstein Reference Goldstein2005; Hastorf et al. Reference Hastorf, Whitehead, Bruno, Wright, Staller, Tykot and Benz2006). Chicha is highly significant to Andean people and is consumed during labor parties, planting and harvesting ceremonies, and feasts in both the Andean past and ethnographic present (Hastorf and Johannessen Reference Hastorf and Johannessen1993:118, citing Cavero Carrasco Reference Cavero Carrasco‚1986; Skar Reference Skar‚1981; Wagner Reference Wagner1978). Allen (Reference Allen1988) describes chicha as a central bonding material, and Hastorf and Johannessen (Reference Hastorf and Johannessen1993:118) agree, writing, “It is a symbolic seal to contracts—spiritual (e.g., asking for fertility of the herds and land), economic (e.g., work and exchange of goods), and social (e.g., marriage).”
The ancient Tiwanaku of the Bolivian altiplano considered maize to be a luxury imported food. It has poor resistance to frost and thus was nonlocal to the altiplano; however, it is native to the Peruvian coastal valleys (Hastorf et al. Reference Hastorf, Whitehead, Bruno, Wright, Staller, Tykot and Benz2006). Throughout the Middle Horizon (AD 600–1100), the Tiwanaku settlers expanded from the altiplano and established colonies in the Peruvian coastal valleys to acquire maize and other lowland-valley crops (Berenguer Rodríguez Reference Berenguer Rodríguez1998; Goldstein Reference Goldstein1989, Reference Goldstein1990, Reference Goldstein and Aldenderfer1993, Reference Goldstein and Bray2003, Reference Goldstein2005; Hastorf et al. Reference Hastorf, Whitehead, Bruno, Wright, Staller, Tykot and Benz2006; Janusek Reference Janusek2002; Knudson et al. Reference Knudson, Goldstein, Dahlstedt, Somerville and Schoeninger2014; Kolata Reference Kolata1986, Reference Kolata1993; Ponce Sanginés Reference Ponce Sanginés1980). Although maize was mainly imported to the altiplano from coastal-valley colonies, Tiwanaku altiplano farmers learned to grow the crop in the “microclimatic pockets” near Lake Titicaca (Langlie Reference Langlie2018:170).
Vessels for chicha have served as markers for the Middle Horizon Tiwanaku expansion. The Tiwanaku corporate ceramic style was defined by the new culinary and social practices associated with chicha that emerged in the Titicaca Basin in the earlier Terminal Late Formative (around AD 350–600), and vessels for chicha making and drinking, such as the kero, quickly diffused into the maize-growing coastal valleys with the spread of Tiwanaku after AD 600 (Goldstein Reference Goldstein and Bray2003, Reference Goldstein2005). Goldstein (Reference Goldstein and Bray2003) emphasizes the economic role of chicha and how the tradition of sponsored work-party feasting in which chicha was consumed could have been a way for the Tiwanaku polity to control production, one in which alcohol played a vital function in encouraging labor (Dietler Reference Dietler1990:368; Goldstein Reference Goldstein and Bray2003; Janusek Reference Janusek2008; Kolata Reference Kolata and Kolata2003). The asymmetrical exchange of labor for chicha began with the Tiwanaku, and the political value of maize developed just as the Tiwanaku expanded into the maize-producing valleys (Goldstein Reference Goldstein and Bray2003:148).
The presence of maize in the Tiwanaku homeland and colonial sites also supports the model of Tiwanaku expansion. Maize kernels and cupules have been recovered from various contexts of Tiwanaku's urban and ceremonial center, demonstrating a 25% ubiquity in a region where maize does not readily grow; in addition, cupule morphology suggests that at least some of this maize was imported from the western valleys (Hastorf et al. Reference Hastorf, Whitehead, Bruno, Wright, Staller, Tykot and Benz2006:430). Maize has been recovered in abundance and high ubiquity from Tiwanaku colonial sites in the Moquegua Valley, specifically from domestic and funerary contexts at Rio Muerto (Vergel Rodriguez and León Reference Vergel Rodriguez and León2009), domestic contexts at Omo M10 (Muñoz Rojas et al. Reference Muñoz Rojas, Boswell, Somerville, Goldstein and Goldstein2009), and the Omo Temple (Gaggio Reference Gaggio2014; Gaggio and Goldstein Reference Gaggio and Goldstein2015). In 2008, the Rio Muerto project at the site of M43 recovered 140 whole maize cobs and several human coprolites showing maize consumption from a single 2 × 3 m domestic unit, suggesting surplus production of maize within the Moquegua Valley (Boswell Reference Boswell2008; Somerville et al. Reference Somerville, Goldstein, Baitzel, Bruwelheide, Dahlstedt, Yzurdiaga, Raubenheimer, Knudson and Schoeninger2015).
Isotopic studies at the Tiwanaku type site suggest that elites enjoyed differential access to imported maize (Berryman Reference Berryman2010), and recent stable isotopic analysis of human skeletal remains spanning 2,000 years shows an increase in C4 signals, suggesting maize became more important in the Lake Titicaca Basin throughout the Middle Horizon (Miller et al. Reference Miller, Iain Kendall, Capriles, Bruno, Evershed and Hastorf2021). Isotope analyses of Tiwanaku settlers in the lowland valleys have demonstrated maize-heavy Tiwanaku colonial diets, particularly among male settlers (Santillan Goode Reference Santillan Goode2018; Somerville et al. Reference Somerville, Goldstein, Baitzel, Bruwelheide, Dahlstedt, Yzurdiaga, Raubenheimer, Knudson and Schoeninger2015). Somerville and colleagues (Reference Somerville, Goldstein, Baitzel, Bruwelheide, Dahlstedt, Yzurdiaga, Raubenheimer, Knudson and Schoeninger2015) found that the Tiwanaku colonists consumed more C4 plants than both earlier non-Tiwanaku people in Moquegua and Tiwanaku people living in the altiplano homeland, correlating the high C4 dietary signal to maize because of the overwhelming presence of maize at Moquegua Valley Tiwanaku sites.
The maize-heavy diets of Tiwanaku colonists support a model of how Tiwanaku people settled lower-elevation zones to grow frost-intolerant maize and export the cultivar back to the altiplano homeland (Somerville et al. Reference Somerville, Goldstein, Baitzel, Bruwelheide, Dahlstedt, Yzurdiaga, Raubenheimer, Knudson and Schoeninger2015:418). We caution, however, that maize may be overrepresented in isotopic analyses of the provincial Tiwanaku diet, which attribute the C4 dietary signal to maize even though Amaranthus spp. may also contribute to C4 signatures (Cadwallader et al. Reference Cadwallader, Beresford-Jones, Whaley and O'Connell2012; Santana-Sagredo et al. Reference Santana-Sagredo, Schulting, Méndez-Quiros, Vidal-Elgueta, Uribe, Loyola and Maturana-Fernández2021). Similarly, maize's role might be accentuated in macrobotanical samples because of its large seed and cob size and thus visibility in archaeological-coarse screening in hyper-arid desert conditions. Sampling under these conditions often fails to recover smaller seed-size cultivars like members of the Amaranthaceae family. In what follows, we argue that using systematic archaeobotanical sampling and graduated-fine dry-screening techniques reveals other cultivars’ equally important role in the provincial Tiwanaku diet. We suggest that the inhabitants of these lowland Tiwanaku sites consumed large quantities of higher-valley and highland Amaranthaceae cultivars in addition to lowland maize.
Amaranthaceae Cultivars and the Tiwanaku Civilization
Amaranthaceae cultivars may grow in many niches, but we focus here on species originating in the upper-elevation zones of the Andes (2,000–4,000 m asl). Quinoa domestication (Chenopodium quinoa, from the Quechua “kinua”) originated in the Andes (Aguilar and Jacobsen Reference Aguilar and Jacobsen2003; Garcia Reference Garcia2003; Hernández Bermejo and León Reference Hernández Bermejo and León1994). Throughout the Archaic period (8000–3000 BC), hunters and gatherers in Peru, Argentina, and Chile consumed wild species of Chenopodium, and their handling likely led to morphological changes in the cultigen (Planella Reference Planella2019; Planella et al. Reference Planella, López, Bruno, Bazile, Bertero and Nieto2015).
Morphological markers of Chenopodium domestication include an increase in seed size, a more prominent “beak,” and a smooth, thinner testa, which makes the perisperm visible and results in a lighter seed color (Bruno Reference Bruno, Zeder, Bradley, Emshwiller and Smith2006; Langlie Reference Langlie2019; Langlie et al. Reference Langlie Brie, Hastorf, Bruno, Bermann, Bonzani and Condarco2011). Archaeobotanical studies have uncovered charred Chenopodium seeds with morphological characteristics showing emerging human manipulation from two late Archaic Chilean-Andean contexts, dating to 1250–980 BC and 1460–1340 BC (Planella et al. Reference Planella, Cornejo and Tagle2005, Reference Planella, Scherson and Mcrostie2011, Reference Planella, López, Bruno, Bazile, Bertero and Nieto2015).
Altiplano people cultivated domesticated Chenopodium quinoa (quinoa) as early as 1500 BC at Chiripa on the Taraco Peninsula of Lake Titicaca (Bruno Reference Bruno, Zeder, Bradley, Emshwiller and Smith2006; Bruno and Whitehead Reference Bruno and Whitehead2003; Bruno et al. Reference Bruno, Capriles, Hastorf, Fritz, Marie Weide, Domic and Baker2021) and by 1300 BC in La Barca, Bolivia (Langlie et al. Reference Langlie Brie, Hastorf, Bruno, Bermann, Bonzani and Condarco2011). Early Formative (1600–800 BC) altiplano people practiced small-scale quinoa agriculture (Bruno Reference Bruno2008:22). Quinoa became the primary staple food by the Late Formative (250 BC–AD 500) when its ubiquity surpassed 90% in altiplano sites on the Taraco Peninsula (Bruno et al. Reference Bruno, Capriles, Hastorf, Fritz, Marie Weide, Domic and Baker2021). At Tiwanaku, studies suggest that the proportions of quinoa increased throughout the Late Formative and that raised fields were created for quinoa cultivation (Wright et al. Reference Wright, Hastorf, Lennstrom and Kolata2003).
Quinoa was likely the most important food source for the Tiwanaku residents of the southern Titicaca Basin (Wright et al. Reference Wright, Hastorf, Lennstrom and Kolata2003). Seeds may have thickened soups, been ground into flour, boiled to make chicha de quinoa (aloja), or malted for alcohol (Cutler and Cárdenas Reference Cutler and Cárdenas1947:34, 39; Goldstein et al. Reference Goldstein, Coleman, Williams, Jennings and Bowser2009; Towle Reference Towle1961:36). Although quinoa was found in everyone's homes, research suggests that lower-status residents at Tiwanaku consumed quinoa and potatoes more frequently than higher-status residents did (Wright et al. Reference Wright, Hastorf, Lennstrom and Kolata2003).
Along with quinoa and other frost-resistant crops, amaranth was traditionally grown at 2,000–3,000 m asl (Hernández Bermejo and León Reference Hernández Bermejo and León1994:128) and in high elevation along with potatoes (Pearsall Reference Pearsall, Silverman and Isbell2008:107). Domesticated several times, Amaranthus includes three domesticated species, A. hypochondriacus, A. cruentus, and A. caudatus (Gómez Pando and Ríos Alfaro Reference Gómez Pando and Alfaro2020; Mallory et al. Reference Mallory, Hall, McNabb, Pratt, Jellen and Maughan2008; Maughan et al. Reference Maughan, Smith, Fairbanks and Jellen2011; Sauer Reference Sauer1967), and was an important food source in the precontact New World (Sauer Reference Sauer and Steward1950). Amaranthus caudatus (kiwicha) is grown in the Peruvian, Bolivian, and northwestern Argentinian Andes (Towle Reference Towle1961). By 1500–1000 BC, quinoa, cañihua (Chenopodium pallidicaule), and achita or coimi (Amaranthus caudatus, kiwicha) were part of the “altiplano economy,” along with three kinds of legumes—tarwi (Lupinus mutabilis), jiquima (Pachyrhizus ahipa), and the common bean (Phaseolus vulgaris)—and four types of tubers: potato (Solanum tuberosum), oca (Oxalis tuberosa), mashua (Tropaeolum tuberosum), and ulluco (Ullucus tuberosus; Browman Reference Browman1981:410; Pickersgill and Heiser Reference Pickersgill‚, Heiser and Browman1978). Wild Amaranthus spp. seeds have been recovered from the Taraco Peninsula and at Tiwanaku (Bruno and Hastorf Reference Bruno, Hastorf, Capriles and Tripcevich2016) and, as mentioned, are described as a “possible casual food source” (Lennstrom et al. Reference Lennstrom, Hastorf and Wright1991a:6, Reference Lennstrom, Hastorf and Wright1991b:6). According to Sauer (Reference Sauer1967:104), all amaranth seeds are likely to be edible and taste like cereals when prepared properly, and amaranth leaves and stalks may be boiled as food.
The uses of quinoa and amaranth go beyond food, and the taxa may enter the archaeological record in various ways. For example, Aymara farmers burn dry quinoa grain stalks to create an ash that activates the alkaloids in coca while chewing (Bruno Reference Bruno2008:193, 273), and amaranth seeds have medicinal uses (Biwer Reference Biwer2019:126, citing Brack Egg Reference Brack Egg1999:27). Each taxon may be thought of as a field weed that is found in “canal-fed small holdings” between 1,400 and 2,500 m asl and in disturbed habitats (Biwer Reference Biwer2019:125; Bruno Reference Bruno2014:7; Bruno and Whitehead Reference Bruno and Whitehead2003:340; Lennstrom et al. Reference Lennstrom, Hastorf and Wright1991a:6, Reference Lennstrom, Hastorf and Wright1991b:6; Sauer Reference Sauer1967:104). This is especially true for Amaranthus species because only three of around 60 species are domesticated (Stallknecht and Schultz-Schaeffer Reference Stallknecht, Schultz-Schaeffer, Janick and Simon1993; Towle Reference Towle1961:37) and for wild Chenopodium sp. seeds, such as quinoa negra, or “quinoa's weedy counterpart” (Bruno Reference Bruno2008:152). As a result, the presence of wild Amaranthus spp. or Chenopodium spp. at a site might indicate that these seeds were inadvertently brought into the space through field-processing activities (Biwer Reference Biwer2019:125) or might suggest that camelids consumed the seeds, which then entered the site as camelid dung that was burned as fuel (Bruno and Hastorf Reference Bruno, Hastorf, Capriles and Tripcevich2016; Hastorf and Wright Reference Hastorf and Wright1998).
Study Sites
The Locumba Valley is in the present-day department of Tacna, south of the Moquegua (Osmore) Valley and north of the Sama, Caplina, and the Chilean Lluta and Azapa Valleys. The sector of the Locumba Valley under consideration is no higher than 1,500 m asl in the Peruvian desert region (Figure 1). This area is suitable for irrigating lowland crops, including tropical fruits, coca, peppers, peanuts, beans, chili peppers, and maize. The site of Cerro San Antonio (L1) is in the middle Locumba Valley and is 35 km from the coast at 600 m asl. This multicomponent site covers 166 ha, of which the Tiwanaku occupation comprises three sectors defined by domestic materials (Sectors A, L, and U) and 10 associated mortuary sectors (Sitek Reference Sitek2022; Sitek and Goldstein Reference Sitek and Goldstein2016). Our analysis of Tiwanaku provincial foodways focuses on Sectors A and L, defined by dense surface midden deposits and remnants of domestic architecture, whereas Sector U is characterized by sparse surface scatter.
We compare L1 findings to data from a Tiwanaku colonial site in the neighboring Moquegua Valley and the central Tiwanaku site in the altiplano. The Tiwanaku homeland is located about 3,860 m asl and 20 km south of Lake Titicaca. There, people gathered lacustrine resources on small totora reed boats, relied on domesticated llamas and alpacas as beasts of burden and as sources of wool and food, and grew frost-resistant crops like quinoa and potatoes (Goldstein Reference Goldstein2005). Throughout the Middle Horizon (AD 600–1100), Tiwanaku altiplano people traveled directly to Moquegua for lowland resources and settled at Rio Muerto (Knudson et al. Reference Knudson, Goldstein, Dahlstedt, Somerville and Schoeninger2014), one of the largest settlements in the valley (Somerville et al. Reference Somerville, Goldstein, Baitzel, Bruwelheide, Dahlstedt, Yzurdiaga, Raubenheimer, Knudson and Schoeninger2015:409). The Rio Muerto site of M43 is in the Moquegua Valley at 900 m asl, sharing a similar ecological placement to the site of L1 in the Locumba Valley.
Methods
Under the Proyecto Arqueológico Locumba (PAL) in 2016 and 2019, Sitek and Goldstein conducted surface collections and household-archaeology excavations in the three Middle Horizon (AD 600–1100) residential sectors at L1. Field excavation in L1 employed a fine-screening method and captured most botanical materials larger than 1 mm in size. Maize remains (kernels and cobs) were the most ubiquitous domesticate recovered, found in 53% of excavated contexts. Both by weight (94.24%) and count (68.11%), most of these fine-screened maize remains were cob fragments. In addition, Sitek collected 134 sediment samples from excavated domestic contexts (Goldstein and Oquiche Hernani Reference Goldstein and Hernani2019; Sitek Reference Sitek2022). From August to September 2019, 36 sediment samples from the primary residential sectors L1A and L1L were selected for archaeobotanical analysis (Figure 2).

Figure 2. Site of Cerro San Antonio (L1). Archaeobotanical samples analyzed are from the primary residential sectors L1A and L1L (figure created by Matthew J. Sitek).
Analyzing 0.5 L of each sample was sufficient because the desert environment of the Locumba Valley allows for excellent macrobotanical preservation. Decomposers do not grow in this desiccated context (Gallagher Reference Gallagher, Marston, Guedes and Warinner2014), and dry-sieving techniques are most appropriate. Implementing flotation would have required unnecessary labor; more importantly, adding water to dry sediment samples may damage delicate, desiccated macrobotanical specimens (Pearsall Reference Pearsall2000; Wagner Reference Wagner, Popper and Hastorf1988; White and Shelton Reference White, Shelton, Marston, Guedes and Warinner2014). In the recovery process, each sample was sifted through 4.0 mm, 2.0 mm, 1.0 mm, and 0.5 mm sieves, using a brush to ease sediment and remains through the mesh.
After the samples were sieved, inorganic and organic materials were manually extracted from the fractions of each sample, using a stereomicroscope for the smaller fractions. The findings were identified using a preliminary photographic seed reference guide that Cindy Vergel created for Moquegua Tiwanaku sites in 2008 (Vergel Rodriguez and León Reference Vergel Rodriguez and León2009) and that Giacomo Gaggio supplemented in 2014; this guide has been extensively expanded and amended with the help of Jade d'Alpoim Guedes, Matthew Biwer, Maria Bruno, and Christine Hastorf. Finally, identified specimens were counted, weighed, and recorded (Supplemental Table 1).
To explore the importance of homeland altiplano foods at the site of L1, we contrasted the proportions of higher-valley and highland (about 2,000–4,000 m asl) Amaranthaceae seeds to lowland-associated (around 0–2,000 m asl) cultivars, molle fruits, peanut seeds, bean seeds, algarrobo endocarps, maize kernels and cobs, and ají seeds (Figure 3). Considering taphonomy, our analysis involved (1) maize kernels and (2) both maize kernels and cobs. Maize kernels might seem more appropriate for taxa comparisons involving consumable seeds and fruits. One might argue, however, that the tiny yet durable Amaranthaceae seeds—which also exist in large numbers on one panicle and easily bounce and fall—are more likely to be preserved than maize kernels, which were consumed and often boiled to make chicha. Maize cobs were discarded and thus more likely to be preserved than maize kernels.

Figure 3. L1 food taxa: (a) Amaranthus sp. seed; (b) Chenopodium quinoa seed; (c) wild Chenopodium sp. seed (labeled as “cf. Chenopodium sp.” in Garvin_PAL2019PEB_data) from L1 with reference to a wild Chenopodium sp. seed from M43 (photograph by Cindy Vergel); (d) Schinus molle fruit; (e) Prosopis sp. endocarp; (f) Zea mays kernels; (g) Zea mays fragment and whole cob; (h) cf. Arachis hypogaea seed; (i) Phaseolus spp. seeds; and (j) Capsicum spp. seeds (except where noted, photographs by Daniel Echecopar). (Color online)
Figures 4 and 5 show the relative proportions of highland to lowland foods at L1 based on taxa counts and weights. In the count-based analyses, each whole and fragmented seed has a value of 1 because a fragment includes more than 50% of the seed. The Fabaceae category includes seeds split down the axis of the hilum, so a “half-seed” is given a value of 0.5. Whole and fragments of maize cobs are each given a value of 1. The graphs comparing weights are most helpful in illustrating the diversity in the set of lowland foods and highlighting the significance of more massive seeds with low counts but with high nutritional or caloric value.

Figure 4. L1 count-based comparisons (n = 36). Graphs A and C show the total proportion of higher-valley and highland taxa (Chenopodium spp. and Amaranthus sp.) and the total proportion of lowland taxa. Graphs B and D show individual-taxa proportions. Graphs A and B use only maize kernels for calculations, and Graphs C and D include both maize kernels and cobs.

Figure 5. L1 weight-based comparisons (n = 36). Graphs E and G show the total proportion of higher-valley and highland taxa (Chenopodium spp. and Amaranthus sp.) and the total proportion of lowland taxa. Graphs F and H show individual-taxa proportions. Graphs E and F use only maize kernels for calculations, and Graphs G and H include both maize kernels and cobs.
We then compared the L1 food taxa proportions to the same set of cultivars at the altiplano Tiwanaku type site (Hastorf et al. Reference Hastorf, Whitehead, Bruno, Wright, Staller, Tykot and Benz2006; Wright et al. Reference Wright, Hastorf, Lennstrom and Kolata2003) and the Rio Muerto M43 site in Moquegua (Goldstein Reference Goldstein2005; Vergel Rodriguez and León Reference Vergel Rodriguez and León2009). Archaeobotanical comparisons with the Tiwanaku capital are complicated by the radically different preservation conditions of the wet Bolivian altiplano, which necessitated flotation methods for archaeobotanical analysis. Nonetheless, it was possible to draw some comparisons from the Tiwanaku database (Wright et al. Reference Wright, Hastorf, Lennstrom and Kolata2003; Christine Hastorf, personal communication 2022). Our analysis used counts of Chenopodium spp., Amaranthus sp., maize kernels, Capsicum sp., and domesticated legumes from 664 Tiwanaku (TIW) samplesFootnote 1 from six units. TIW tubers were purposely excluded from the analysis, because Amaranthaceae cultivars are the focus of this study.
Excavation of Rio Muerto M43 Unit 6, an exceptionally well-preserved stratified household midden in the M43F domestic sector, produced comparable sediment samples for archaeobotanical analysis. As at L1, large quantities of macrobotanical remains recovered from M43 field-screened collections emphasized larger cultigens, like maize and beans, whereas the archaeobotanical data tell a different story.
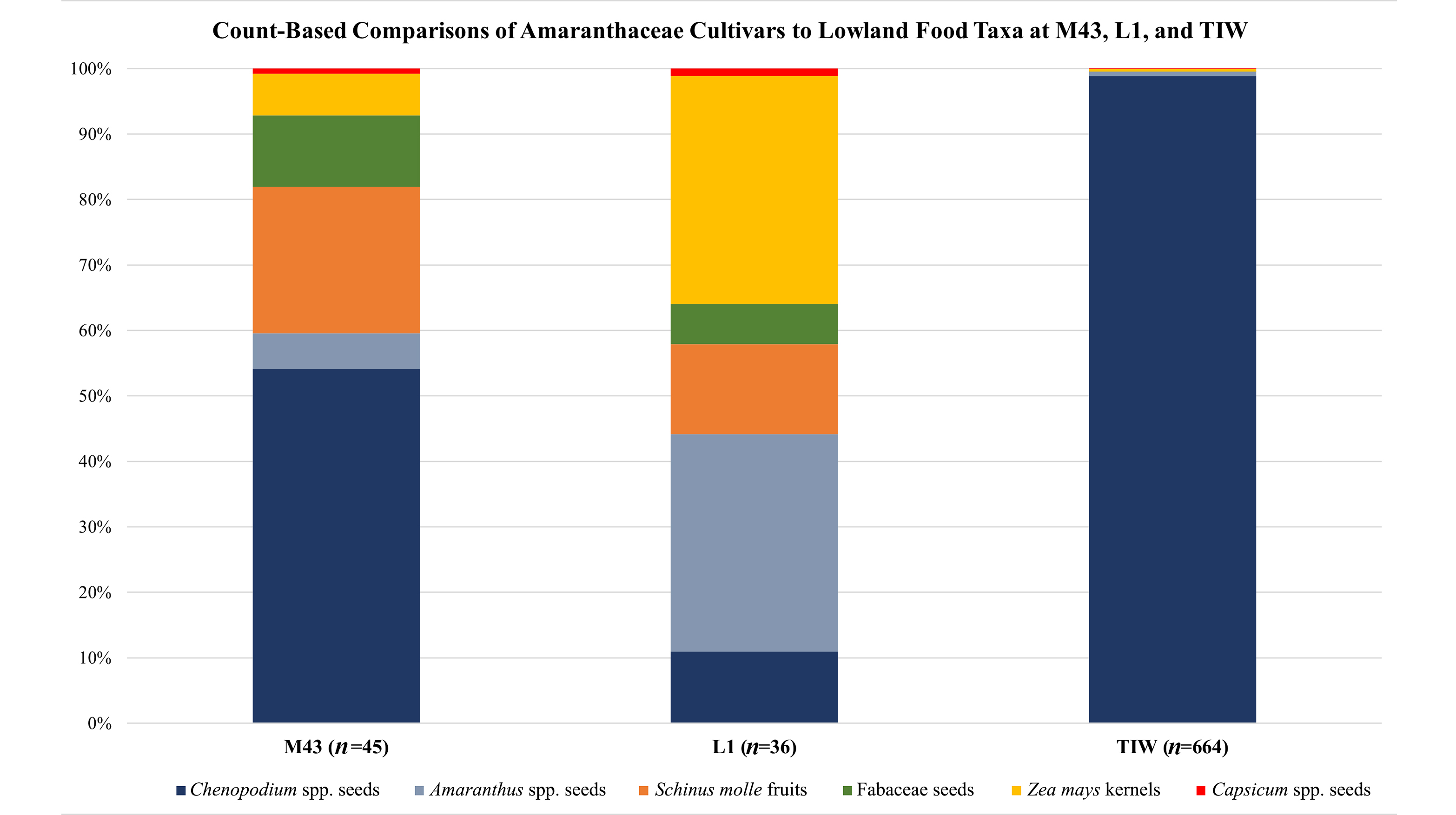
The L1, M43, and TIW comparative analysis involved counts and only maize kernels (Figure 6). We note that L1 includes an outlier sample excluded from the L1, M43, and TIW comparative analysis. Sample L1-4161 contained 481 maize kernels (11 whole and 470 fragments) of the 498 kernels (16 whole and 482 fragments), or 96.59% of the total kernel count at L1 (Supplemental Figure 1). The TIW data included a domesticated legume category (“DOMLEGUM”), so to deal with seed-identification uncertainties, we combined Arachis hypogaea seeds, Phaseolus spp. seeds, and Prosopis spp. endocarps to form a Fabaceae category for L1 and M43. Moreover, TIW wild Leguminosae/Fabaceae (“WILDLEG”) were excluded from the analysis. After reviewing the 2008 Moquegua Tiwanaku photographic collection, we also recognize that the M43 Amaranthus spp. category may have included some wild Chenopodium sp. seeds.

Figure 6. Count-based, food-taxa comparisons at the Tiwanaku colonial sites M43 (900 m asl; Goldstein Reference Goldstein2005; Vergel Rodriguez and León Reference Vergel Rodriguez and León2009), L1 (600 m asl), and the Tiwanaku altiplano capital (around 3,860 m asl; Hastorf et al. Reference Hastorf, Whitehead, Bruno, Wright, Staller, Tykot and Benz2006; Wright et al. Reference Wright, Hastorf, Lennstrom and Kolata2003).
Food Taxa Recovered at Cerro San Antonio (L1)
We compared the proportions of eight historically and ethnographically important food taxa at the site of L1 in the Locumba Valley (Figure 3).Footnote 2 In addition to quinoa, amaranth, and maize cultivars, we explored Arachis hypogaea (peanut), Capsicum spp. (ají), Phaseolus spp. (bean), Prosopis sp. (algarrobo), and Schinus molle (molle).
Peanuts served as a nutritious complement to the coastal diet of precontact Peru. They were toasted, fried, boiled, and ground into chicha de maní (Cutler and Cárdenas Reference Cutler and Cárdenas1947:34; Fernández Honores and Rodríguez Rodríguez Reference Fernández Honores and Rodríguez Rodríguez2007:107; Masur et al. Reference Masur, Millaire and Blake2018; Stalker Reference Stalker1997). Ají flavors largely contribute to Andean cuisine (Chiou et al. Reference Chiou, Hastorf, Vásquez, Tham, Bonavia, Dillehay and Dillehay2017; Pearsall Reference Pearsall, Silverman and Isbell2008). Peruvian peoples regularly grew beans in the temperate coastal and middle valleys by the Middle Horizon (AD 600–1100). The bean, a nitrogen-fixing plant high in the amino acid lysine, agriculturally and nutritionally complements maize, a nitrogen-depleting plant deficient in lysine but higher than the bean in caloric value (Biwer Reference Biwer2019; Mt. Pleasant Reference Mt. Pleasant2016; Towle Reference Towle1961).
Algarrobo and molle are notable multipurpose plants of coastal Peru (Fernández Honores and Rodríguez Rodríguez Reference Fernández Honores and Rodríguez Rodríguez2007; Goldstein and Coleman Reference Goldstein and Coleman2004; Hernández Reference Hernández1942:280; Towle Reference Towle1961). Archaeological evidence suggests that people ingested raw algarrobo pods and processed pods into syrup. They may also have ground pods into flour for beverages or a porridge mix (Capparelli and Lema Reference Capparelli and Lema2011). Molle seeds may have served as a condiment, resembling black or white pepper (Goldstein and Coleman Reference Goldstein and Coleman2004:525).
L1 Comparisons of Amaranthaceae Cultivars versus Lowland Cultivars
Table 1 includes raw counts and weights of the eight food taxa at L1. Count-based taxa comparisons show that highland-associated Amaranthaceae cultivars account for 40%–44% of the archaeobotanical assemblage. These figures do not change dramatically when either using only maize kernels or combining maize kernels and cobs (Figure 4). When weight is measured, the proportion of Amaranthaceae cultivars drops considerably, comprising 0.73%–1.01% (Figure 5). As in the count-based graphs, molle and maize are significant, and the importance of Prosopis sp. becomes more apparent in the weight-based graphs. Finally, the low counts of cf. Arachis hypogaea and Phaseolus spp. seeds that are not visible in the count-based graphs are visible in the weight-based comparisons.
Table 1. L1 Taxa Counts and Weights.

a The Garvin_PAL2019PEB_data sheet includes “whole” and “fragment” categories, where each fragment is greater than 50% of the taxon. Whole and fragment taxa are each given a value of 1 to create the sum counts for this table.
b Wild Chenopodium sp. seeds are labeled as cf. Chenopodium sp. in Garvin_PAL2019PEB_data.
c The Garvin_PAL2019PEB_data sheet includes “half” Phaseolus spp. seeds, each given a value of 0.5 to create the sum counts for this table.
d Prosopis sp. endocarps are labeled as seeds in Garvin_PAL2019PEB_data.
Comparing Diets at L1 to the Moquegua Tiwanaku Colony M43 and the Tiwanaku Capital
Figure 6 overwhelmingly emphasizes Chenopodium spp. as the dominant cultigens at the Tiwanaku capital (98.88%). At TIW, Amaranthus spp. represents only 0.67%, and combined lowland cultigens maize, Capsicum sp., and Fabaceae comprise 0.45%. L1 and M43 colonial residents had remarkably similar diets, relying heavily on Amaranthaceae cultivars: L1 shows 67.06% Amaranthaceae (16.09% Chenopodium spp. and 50.97% Amaranthus spp.), and M43 shows 59.54% Amaranthaceae (54.16% Chenopodium spp. and 5.38% Amaranthus spp.).
Although more L1 sediment samples should be analyzed, the L1 preliminary findings (2019) and the M43 archaeobotanical findings (2008) highlight Amaranthaceae cultivars in the diet. This contrasts with the emphasis on maize in the Moquegua diet derived from excavation findings (Gaggio Reference Gaggio2014; Gaggio and Goldstein Reference Gaggio and Goldstein2015; Muñoz Rojas et al. Reference Muñoz Rojas, Boswell, Somerville, Goldstein and Goldstein2009; Vergel Rodriguez and León Reference Vergel Rodriguez and León2009), maize-cupule analysis (Hastorf et al. Reference Hastorf, Whitehead, Bruno, Wright, Staller, Tykot and Benz2006), and isotope analyses (Santillan Goode Reference Santillan Goode2018; Somerville et al. Reference Somerville, Goldstein, Baitzel, Bruwelheide, Dahlstedt, Yzurdiaga, Raubenheimer, Knudson and Schoeninger2015). In short, the archaeobotanical findings subvert our perspective and show that inhabitants of Tiwanaku colonial sites heavily consumed cultivars from their homeland and upper Andean zone.
Discussion: The Implications of Amaranthaceae at L1
The preeminent cultigen of Tiwanaku, Chenopodium quinoa, and a similar cultigen, Amaranthus spp., contributed significantly to the Tiwanaku colonial diets at Cerro San Antonio (L1) and Rio Muerto (M43). At L1, Amaranthaceae seeds are found in samples recovered from hearths, domestic-waste contexts, and a storage pit containing maize and beans.
Amaranthus sp. comprises 75.16% of the L1 Amaranthaceae assemblage (Figure 7). The large proportion suggests that this weedy seed grew locally at L1 and that Tiwanaku colonists came across a familiar crop or at least one that resembled quinoa. Weedy Amaranthus spp. grow in disturbed habitats (Lennstrom et al. Reference Lennstrom, Hastorf and Wright1991a:6, Reference Lennstrom, Hastorf and Wright1991b:6) and at sea level (Hernández Bermejo and León Reference Hernández Bermejo and León1994). No domesticated Amaranthus sp. seeds, like kiwicha, have been recovered from L1 (Figure 8). Weedy Amaranthus seeds were likely growing among field crops or in pastures where camelids graze, so these seeds may have entered domestic spaces in camelid dung that was burned for cooking (Garvin Reference Garvin2020); this was a common Tiwanaku altiplano practice (Bruno and Hastorf Reference Bruno, Hastorf, Capriles and Tripcevich2016). It is also possible that Tiwanaku colonists consumed the Amaranthus sp. seeds, along with the leaves and stalks (Sauer Reference Sauer1967:104).

Figure 7. L1 Amaranthaceae proportions. (Color online)

Figure 8. L1 Amaranthaceae domesticated seeds versus wild/weedy seeds (n = 36). (Color online)
Chenopodium quinoa comprises 19.94% of the Amaranthaceae assemblage (Figure 7), and its presence at L1 suggests that Tiwanaku colonists continued to consume the highland-associated cultivar by either acquiring harvested altiplano quinoa through trade or growing quinoa in the Locumba Valley.
One might argue that L1 residents acquired quinoa through trade. According to John V. Murra's (Reference Murra and Murra1972) vertical archipelago model, the Andean nonmarket system of production and exchange developed from small groups of people colonizing complementary ecological zones of the vertically complex ecological landscape, engaging in economic activities suitable to the zones, and acquiring resources from the different ecological zones through exchange. We might speculate that the Tiwanaku colonists left the altiplano (3,800 m asl) and established permanent or long-term residence at L1 in the Locumba Valley (600 m asl). At L1, the colonists cultivated food crops, beans, and maize that readily grew in this lowland zone and then exchanged these cultivars for highland quinoa. As mentioned, quinoa is native to the altiplano and was the most important food source for Late Formative and Tiwanaku residents of the Titicaca Basin (Wright et al. Reference Wright, Hastorf, Lennstrom and Kolata2003). Considering this long-standing tradition and the demonstrated Tiwanaku trade of lowland crops to the highlands, it is possible that the same caravans could have brought highland chenopod produce to the lowlands or that lowland settlers brought highland crops home after visiting Tiwanaku for seasonal festivities.
Yet Tiwanaku colonists instead may have grown quinoa locally in Locumba. First, quinoa and amaranth are known to adapt to various agroclimatic and edaphic conditions (Table 2). This is because of their high genetic variability developed over their history of domestication in the altiplano and dynamic Andean environments, where crops are susceptible to frost, limited rainfall, a high rate of evapotranspiration, low soil-water retention, and saline soils (Garcia et al. Reference Garcia, Condori, Castillo, Murphy and Matanguihan2015:26–27; Jacobsen et al. Reference Jacobsen, Mujica and Jenson2003; Nascimento et al. Reference Nascimento, Mota, Coelho, Gueifão, Santos, Matos, Gimenez, Lobo, Samman and Castanheira2014). Quinoa grows in places like the altiplano salt desert of Bolivia, with just 200 mm of rainfall (Aguilar and Jacobsen Reference Aguilar and Jacobsen2003:39, citing Mujica et al. Reference Mujica‚, Jacobsen and Ortiz1998; Jacobsen et al. Reference Jacobsen, Mujica and Jenson2003:102) and even in the Atacama Desert (Fuentes and Bhargava Reference Fuentes and Bhargava2011; Garcia et al. Reference Garcia, Raes, Jacobsen and Michel2007; Jacobsen et al. Reference Jacobsen, Mujica and Jenson2003).
Table 2. Quinoa Cultivation and Adaptability.

Present-day, traditional altiplano farmers are intimately connected to quinoa biodiversity and use practices adapted to variable, adverse conditions. Andrews (Reference Andrews2017:20–21) compiled a list of 195 quinoa names as only part of an even more extensive ethno-taxonomy used by Aymara farmers. In dry conditions, present-day altiplano farmers strategically mix quinoa varieties, such as the drought-resistant Kcoitos native gray variety, frost-resistant Witullas and Wilas varieties, and the more desirable but less hardy white quinoa (Aguilar and Jacobsen Reference Aguilar and Jacobsen2003). Such strategies can hedge bets between desirable and productive varieties and those adapted to potential adverse conditions. Additionally, traditional communities grow quinoa in a spatial and temporal rotation known as the aynoqas system. Although temporal rotation alternates different crops and fallow to avoid soil depletion, the sociospatial aspect of aynoqa also scatters each family's holdings within the larger communities to include parcels in lower and higher microenvironments with different susceptibilities to frost and drought risks (Aguilar and Jacobsen Reference Aguilar and Jacobsen2003:33–34, 36).
It is likely that ancient Tiwanaku farmers, like present-day altiplano farmers, would have known of quinoa biodiversity and the relative drought, salt, pH, and temperature resistance of different varieties and would have brought a comparable set of strategies for quinoa cultivation under lowland desert conditions. Moreover, it does appear that quinoa was present in at least some lowland and desert contexts outside its center of domestication by the Formative (1800 BC–AD 400) or even earlier (Table 3).
Table 3. Archaeological Evidence of Quinoa in Lowland and Desert Contexts.

The mixture of domesticated and wild Chenopodium at L1 offers insight into whether quinoa arrived via trade or was cultivated locally. López and Recalde (Reference López and Recalde2016:432) present the first reliable evidence of quinoa and ajara, or quinoa negra, from the Sierras del Norte of Central Argentina (around 700–300 BP); they suggest that the “Andean crop/weed complex” mitigated crop failure. They also note that precontact people consumed quinoa and weed seeds (López and Recalde Reference López and Recalde2016:431, citing Lagiglia Reference Lagiglia‚2005; López Reference López‚2012; Ratto et al. Reference Ratto, Lema and López2014). During the Early Formative (1600–800 BC), altiplano people harvested and consumed quinoa's weedy counterpart, quinoa negra (Chenopodium quinoa var. melanospermum) in equal or greater proportions than domesticated quinoa (Bruno Reference Bruno2008:22, citing Bruno Reference Bruno‚2001:96–98). In contrast, in the Middle Formative (800–250 BC) people selected seeds and separated quinoa negra from the food supply (Bruno and Whitehead Reference Bruno and Whitehead2003:352).
If quinoa arrived at L1 via trade, quinoa would likely have been carefully processed and cleaned before trade (Ren et al. Reference Ren, Dong, Liu, Guedes, Flad, Ma and Li2020), leaving few wild Chenopodium spp. seeds in the L1 assemblage. Instead, we found that wild Chenopodium sp. comprised 4.91% of the Amaranthaceae assemblage (Figure 7) and 19.75% of the Chenopodium spp. seeds at L1 (Figure 8). We argue that the significant presence of wild Chenopodium, along with weedy Amaranthus seeds, suggests that the L1 Tiwanaku colonists cultivated, prepared, and consumed Amaranthaceae in the Locumba Valley (AD 600–1100) and ultimately reinforced their Tiwanaku homeland identities through these food-related activities.
Conclusion
Dietary investigations of the Middle Horizon Tiwanaku expansion (AD 600–1100) have primarily focused on maize because of its unique economic and ritual roles (Gaggio Reference Gaggio2014; Gaggio and Goldstein Reference Gaggio and Goldstein2015; Hastorf et al. Reference Hastorf, Whitehead, Bruno, Wright, Staller, Tykot and Benz2006; Muñoz Rojas et al. Reference Muñoz Rojas, Boswell, Somerville, Goldstein and Goldstein2009; Santillan Goode Reference Santillan Goode2018; Somerville et al. Reference Somerville, Goldstein, Baitzel, Bruwelheide, Dahlstedt, Yzurdiaga, Raubenheimer, Knudson and Schoeninger2015; Vergel Rodriguez and León Reference Vergel Rodriguez and León2009). We argue that Tiwanaku colonial diet studies should focus more on the presence of Amaranthaceae and other cultivars through archaeobotanical fine screening and taxa comparisons based on counts and weights. Although more samples from L1 should be analyzed, our archaeobotanical analysis found that L1 and M43 residents shared remarkably similar mixed diets, largely including Amaranthaceae cultivars. L1 colonists reinforced their Tiwanaku altiplano identities and cultivated, prepared, and consumed Amaranthaceae cultivars on the lowland frontier because of the strong presence of domesticated, wild, and weedy Amaranthaceae seeds. By cultivating quinoa in a region well outside its natural range, Tiwanaku colonization marked a significant step in developing this important cultivar's ability to adapt to various agroclimatic and edaphic conditions.
Acknowledgments
We thank Drs. Christine Hastorf, Maria Bruno, Matthew Biwer, and Rick Jellen for their help with seed identifications, and Daniel Echecopar for the seeds photographed. We are grateful to Dr. Hastorf for sharing the Tiwanaku data and to Cindy Vergel, Giacomo Gaggio, and the codirector of the Rio Muerto project, Patricia Palacios Filinich, for their contributions to the M43 research referenced in this article. We also thank the codirector of the Proyecto Arqueológico Locumba, Antonio Oquiche Hernani. This research was conducted under MNC Resolucion Viceministerial N°172-2018-VMPCIC-MC.
Funding Statement
This research was funded by the NSF Doctoral Dissertation Improvement Award #1841909 (Goldstein, Sitek) and the UC, San Diego Center for Iberian and Latin America Studies 2019 Tinker Field Research Grant (Garvin Suero).
Data Availability Statement
Original data are available in supplemental materials and in Garvin's (Reference Garvin2020) master's thesis, Department of Anthropology, University of California, San Diego, retrieved from https://escholarship.org/uc/item/4ff0×80c.
Competing Interests
The authors declare none.
Supplemental Material
For supplemental material accompanying the article, visit https://doi.org/10.1017/laq.2023.46.
Supplemental Figure 1. Original archaeobotanical data from the Tiwanaku colonial site of Cerro San Antonio (L1; Garvin Reference Garvin2020).
Supplemental Table 1. Count-Based, Food-Taxa Comparisons at the Tiwanaku Colonial Sites M43 (900 m asl; Goldstein Reference Goldstein2005; Vergel Rodriguez and León Reference Vergel Rodriguez and León2009), and L1 (600 m asl), and the Tiwanaku Altiplano Capital (~3,860 m asl; Hastorf et al. Reference Hastorf, Whitehead, Bruno, Wright, Staller, Tykot and Benz2006; Kolata Reference Kolata and Kolata2003; Wright et al. Reference Wright, Hastorf, Lennstrom and Kolata2003). The L1 outlier sample 4161 is included.














 Open access
Open access