



Non-technical Summary
Monstrocrinus is a most unusual crinoid from the Devonian of Germany. It has long, simple to complex spines attached to most calyx plates. A stem was not associated with the initial specimens on which this genus was named. Thus, it was assumed that Monstrocrinus was a stemless crinoid that sat directly on the sea floor and, when alive, rolled around on its spines. For a variety of reasons, this seems unlikely, but the rolling crinoid lifestyle interpretation has persisted in the literature. More thorough cleaning of historical specimens and, more importantly, a new specimen with an attached column clearly refute the classical lifestyle interpretation and demonstrate that Monstrocrinus was a typical crinoid attached to a long column that was presumably anchored to the sea floor. In addition to revising the lifestyle of this crinoid, the varieties of spine types are redefined with names that more accurately reflect their morphology. Monstrocrinus occurs in Europe and South America, and one previously described species is placed in synonymy so that the genus is now comprised of three species.
Introduction
The historical type localities of numerous crinoid faunas in the Devonian of the Rhenish Slate Mountains are from distinct depositional facies. During the Early Devonian (late Siegenian [late Pragian] to the end of the early Emsian) ~70 crinoid species assigned to 28 genera are known from the famous “Hunsrückschiefer,” exposed between Koblenz, Trier, and Mainz (Bartels et al., Reference Bartels, Briggs and Brassel1998; Hess, Reference Hess, Hess, Ausich, Brett and Simms1999; Südkamp, Reference Südkamp2017). Characteristic pyritized fossils of four primary crinoid clades are represented by the genera Codiacrinus Schultze, Reference Schultze1866; Imitatocrinus Schmidt, Reference Schmidt1934; and Parisangulocrinus Schmidt, Reference Schmidt1934 (Eucladida); Calycanthocrinus Follmann, Reference Follmann1887; and Triacrinus Münster, Reference Münster1839 (Disparida); Hapalocrinus Jaekel, Reference Jaekel1895; and Thallocrinus Jaekel, Reference Jaekel1895 (Camerata); and Eutaxocrinus Springer, Reference Springer1906 (Flexibilia).
The crinoids of the sandy Lower Devonian up to the Cultrijugatus Zone, which were summarized by Schmidt (Reference Schmidt1941), are the second important regional crinoid fauna. They occur at numerous localities along the western and eastern Rhenish Massif in a time slice between the late Siegenian (respectively late Pragian) to the early Eifelian. One hundred and twenty-five species assigned to 34 genera are discussed in this classic monograph, with most specimens preserved as hollow molds. Especially characteristic are the camerate genera Ctenocrinus Bronn, Reference Bronn1840; Acanthocrinus Roemer, Reference Roemer1850; Monstrocrinus Schmidt, Reference Schmidt1941; and Orthocrinus Jaekel, Reference Jaekel1895; as well as the eucladid Eifelocrinus Wanner, Reference Wanner1916 (see Bohatý, Reference Bohatý2009, p. 195). Monstrocrinus, Orthocrinus, and Eifelocrinus also dominate the crinoid fauna of the Emsian–Eifelian boundary within the Rhenish Massif (Bohatý, Reference Bohatý2009, p. 195). This boundary is characterized by the sedimentological change from the sandy sedimentation of the Emsian to the carbonate sedimentation of the Middle Devonian, which is associated with the establishment of extensive reefoid formation during the beginning of the Eifelian (Bohatý, Reference Bohatý2009). This change also meant that the Emsian crinoid soft-bottom dwellers were increasingly associated with hard- and firmground dwellers. Monstrocrinus, Orthocrinus, and Eifelocrinus characterize a crinoid transitional fauna between the Lower and Middle Devonian facies realms, which locally only had a short stratigraphic range.
Supraregionally, Monstrocrinus is the youngest representative of the three genera mentioned with occurrences extending into the middle to upper Eifelian in Brazil, where the genus has the youngest known stratigraphic occurrence (Scheffler et al., Reference Scheffler, Fernandes and Da Fonseca2006, Reference Scheffler, Dias-Da-Silva, Gama, Da Fonseca and Fernandes2011). Monstrocrinus is an enigmatic crinoid with long, elaborate spines attached to numerous calyx plates. Most preserved remains of this crinoid are isolated spines; thus, spine morphology has largely been the basis for species differentiation. Also, because neither an associated column nor a preserved external view of the infrabasal circlet were known previously, a free-living benthic lifestyle was proposed for this unusual crinoid (e.g., Schmidt, Reference Schmidt1941; Ettensohn, Reference Ettensohn1984). The new partially articulated specimens reported here with variable spine shapes on a single individual and an attached column allow a reassessment of Monstrocrinus systematics and paleoecology.
Location and stratigraphy
In the present work, new specimens of the spectacular, if enigmatic, Monstrocrinus are described from the uppermost Lower to Middle Devonian rocks of the Rhenish Slate Mountains (North Rhine-Westphalia and Rhineland-Palatinate, Germany; Fig. 1), which allow important insights into the mode of life and the functional morphology of this unusual crinoid. The material comes from the uppermost section of the Emsian Orthocrinus Beds (Polygnathus costatus patulus Conodont Biozone) of the locus typicus of the type species Monstrocrinus securifer Schmidt, Reference Schmidt1941, at the former brickworks mine (slopes north, east, and south of the former brick factory, UTM: 51°02′22.73″N, 7°51′51.02″E) in the Osterbachtal on today's federal road B54, northwest of Olpe-Lütringhausen (administrative district Arnsberg, Sauerland, North Rhine–Westphalia). The classic site is located at the southeastern flank of the Middle Devonian Attendorn–Elspe Double-Syncline in the transition to the Lower Devonian Siegen Anticline. Additional material is from two outcrops within the Eifel synclines. The first locality is the northern embankment at the “Eschfelder Seifen” on the southwestern flank of the Middle Devonian Ahrdorf Syncline, in the transition area to the Lower Devonian Wiesbaum Anticline (southeast of Dollendorf, NNW of Leudersdorf, North Rhine–Westphalia; UTM: 50°21′45.62″N, 6°43′44.14″E). The second location is in the northern Ahbach Valley, on the north-northeastern flank of the Middle Devonian Hillesheim Syncline, in the transition area to the Lower Devonian Hoffeld Anticline (north-northeast of Üxheim–Ahütte, Rhineland–Palatinate; UTM: 50°20′56.25″N, 6°46′35.71″E). Stratigraphically, this material is from the Eifel synclines in the Lauch Formation (Polygnathus costatus partitus to lowermost P. costatus costatus Conodont biozones) of the lower Eifelian and, therefore, are somewhat younger than those from the Sauerland type locality north of Olpe.
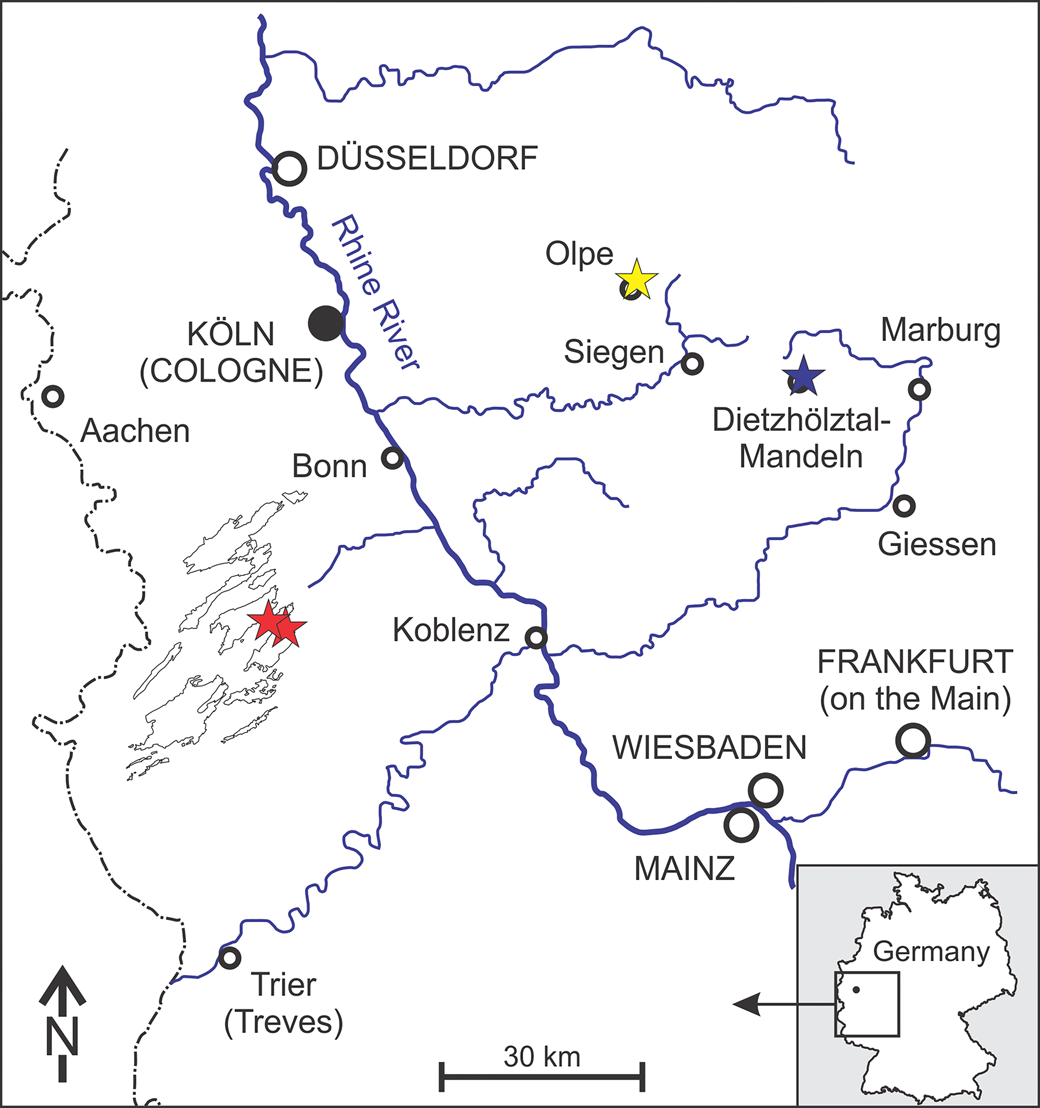
Figure 1. Geographical overview of the Rhenish Massif (modified from Korn, Reference Korn2008, after Walter, Reference Walter1995). The German Monstrocrinus localities are marked with stars: red stars mark the sites within the Eifel synclines (“Eschfelder Seifen,” in the west, and northern Ahbach Valley, in the east); the yellow star indicates the fossil locality in the Sauerland (former brick factory in the Osterbachtal); and the blue star marks the fossil site within the Lahn–Dill district (“Old Municipality Quarry” in Dietzhölztal–Mandeln).
Whereas the type species Monstrocrinus securifer Schmidt, Reference Schmidt1941, is the only recorded Monstrocrinus to date within the Eifel, both species known in Germany occur together within the Emsian Orthocrinus Beds (Polygnathus costatus patulus Conodont Biozone) in the Sauerland (Olpe). However, Monstrocrinus granosus Schmidt, Reference Schmidt1941, already occurs within the upper Emsian Mandeln Formation (Polygnathus serotinus Conodont Biozone) of Dietzhölztal–Mandeln in the Lahn–Dill district in central Hesse, north-northeast of Dillenburg (type locality at UTM: 50°51′22.52″N, 8°20′51.41″E).
Materials and methods
The majority of the Monstrocrinus specimens from the historic M. securifer type locality within the Sauerland (Olpe) and the M. granosus type locality within the Lahn–Dill district in Hesse (Dietzhölztal–Mandeln) are preserved as molds, which has historically led to misinterpretations of Monstrocrinus morphology. For this study, some new preparation work was carried out on the holotypes of both species in order to be able to create new latex molds. These were whitened and digitally photographed with low-angle lighting. The new finds from the lower Eifelian of the Eifel synclines are in calcite preservation. These specimens were mechanically dissected using preparatory needles, micro sand-streaming methods, and fine pneumatic probes.
Repositories and institutional abbreviations
The original material of Monstrocrinus securifer is deposited with the abbreviation IGPB in the Steinmann Institute for Geology, Mineralogy, and Paleontology of the Rhenish Friedrich Wilhelm University of Bonn, Germany. The type material of M. granosus is deposited with the abbreviation SMF in the Senckenberg Research Institute and Natural History Museum Frankfurt on the Main, Germany. Specimen no. NHMMZ PWL 2021/6145-LS is deposited in the State Collection of Natural History of Rhineland–Palatinate at the Natural History Museum Mainz (NHMMZ), Germany. The type material of M. incognitus is housed in the paleontological collection of the Departamento de Geologia e Paleontologia do Museu Nacional, Universidade Federal do Rio de Janeiro (UFRJ), Brazil, under institutional abbreviation no. MN8277-Id. (holotype) and nos. MN8277-Ia, and MN8277-Ib (paratypes).
Systematic paleontology
The superfamilial classification used here follows Cole (Reference Cole2017), Wright (Reference Wright2017), and Wright et al. (Reference Wright, Ausich, Cole, Peter and Rhenberg2017). Family-level classifications follow Moore and Teichert (Reference Moore and Teichert1978). Morphologic terminology follows Webster (Reference Webster1974), Ubaghs (Reference Ubaghs, Moore and Teichert1978a), Ausich et al. (Reference Ausich, Wright, Cole and Sevastopulo2020), and Ausich and Donovan (Reference Ausich and Donovan2023). The plating of interrays is given by the number of plates in each range from proximalmost plate to the last range before the tegmen. In the posterior interray, the primanal is indicated by “P;” and in regular interrays, the first interradial plate is indicated by “1.” A “?” indicates that more distal plating is unknown (Ausich, Reference Ausich2021). Abbreviations used in designating measurements include: CrH, crown height; CaH, calyx height; CaW, calyx width; CoH, column height. An asterisk (“*”) indicates an incomplete measurement.
Class Crinoidea Miller, Reference Miller1821
Subclass Camerata Wachsmuth and Springer, Reference Wachsmuth and Springer1885
Infraclass Eucamerata Cole, Reference Cole2017
Order Diplobathrida Moore and Laudon, Reference Moore and Laudon1943
Superfamily Rhodocrinitoidea Roemer, Reference von Roemer and Bronn1855
Family Rhodocrinitidae Roemer, Reference von Roemer and Bronn1855
Genus Monstrocrinus Schmidt, Reference Schmidt1941
Type species
Monstrocrinus securifer Schmidt, Reference Schmidt1941.
Other species
Monstrocrinus granosus Schmidt, Reference Schmidt1941; and M. incognitus Scheffler et al., Reference Scheffler, Dias-Da-Silva, Gama, Da Fonseca and Fernandes2011.
Emended diagnosis
Rhodocrinitid with a medium bowl- to globe-shaped calyx, distal corners of infrabasal plates not visible in lateral view, proximal regular interrays with 1–3 plating, proximal CD interrays with 1–3 plating, median ray ridges absent, anitaxial ridge absent, primaxil hexagonal, calyx plates with various types of long spines attached to calyx plates, fixed pinnules absent, 20(?) free arms that divide once, brachials chisel biserial.
Occurrence
Upper Emsian (Lower Devonian) to lower Eifelian (Middle Devonian) of Europe (Rhenish Slate Mountains, North Rhine-Westphalia, Rhineland-Palatinate, and Hesse, Germany; and province of Teruel, Aragon, Spain). Middle and upper Eifelian (Middle Devonian) of South America (states of Pará and Tocantins, Brazil) (Ferreira and Fernandes, Reference Ferreira and Fernandes1989; Fernandes et al., Reference Fernandes, Fonseca and Ferreira2000; Scheffler et al., Reference Scheffler, Fernandes and Da Fonseca2006, Reference Scheffler, Fernandes and Da Fonseca2009, Reference Scheffler, Dias-Da-Silva, Gama, Da Fonseca and Fernandes2011, Reference Scheffler, Da Fonseca and Fernandes2015; Gama Jr. and Scheffler, Reference Gama and Scheffler2007; Scheffler, Reference Scheffler2010, Reference Scheffler2015). The occurrence of the genus Monstrocrinus in the Emsian–Eifelian boundary interval of the lowermost part of the Chefar El Ahmar Formation of the Ougarta Mountains in the Sahara province of Béni Abbès in southwestern Algeria in the Maghreb region of North Africa (see Le Menn, Reference Le Menn1990, Reference Le Menn1997) is not proven taxonomically.
Remarks
In addition to complete specimens of Orthocrinus tuberculatus Schmidt, Reference Schmidt1913, and Arthroacantha Williams, Reference Williams1883, Schmidt (Reference Schmidt1924, p. 17) mentioned a new crinoid genus “with unusually monstrous, axe-shaped spines on the basals and radials” from the former brickworks mine in the Osterbachtal, northwest of Olpe-Lütringhausen (see “Location and stratigraphy”). Seventeen years later, Schmidt (Reference Schmidt1941, p. 213–215) described this as Monstrocrinus n. gen. and considered this new discovery to have been a free-living benthic rhodocrinitid genus, which is characterized by “monstrous floating spines” (he interpreted the paddle-like, equatorially projecting spiny appendages of the calyx as useful elements for passive drifting on the sea bottom; see Fig. 2.3) and “very flexible, two-lined arms.” However, he pointed out that the genus was incompletely known and required a full description (Schmidt, Reference Schmidt1941, p. 213). His diagnosis was based on various disarticulated plates of the calyx as well as remains of the “floating spines.” This material was from different specimens and localities, which systematically cannot be assigned unequivocally to a genus or to one of the two Monstrocrinus species described by him. Nevertheless, Schmidt (Reference Schmidt1941, p. 213) combined these individual parts into a genus and species description. In addition, Schmidt (Reference Schmidt1941, p. 213) justified the lack of a stem with the alleged lack of a “stem scar” on the aboral side of the calyx; the column lumen was said to be closed by the fact that the proximal plates of the calyx are supposed to have grown together until the “column hole” was completely closed (Schmidt, Reference Schmidt1941, p. 215). The inconclusive evidence for this is a gelatin cast of the interior of a single calyx remnant preserved as a mold and required a full description (Schmidt, Reference Schmidt1941, p. 216). However, our re-examination of the type material revealed that this assumption was incorrect. Furthermore, Schmidt (Reference Schmidt1941, p. 215) deduced from disarticulated arm remains of his type specimen for M. granosus that the arms of all Monstrocrinus species must have been relatively short.
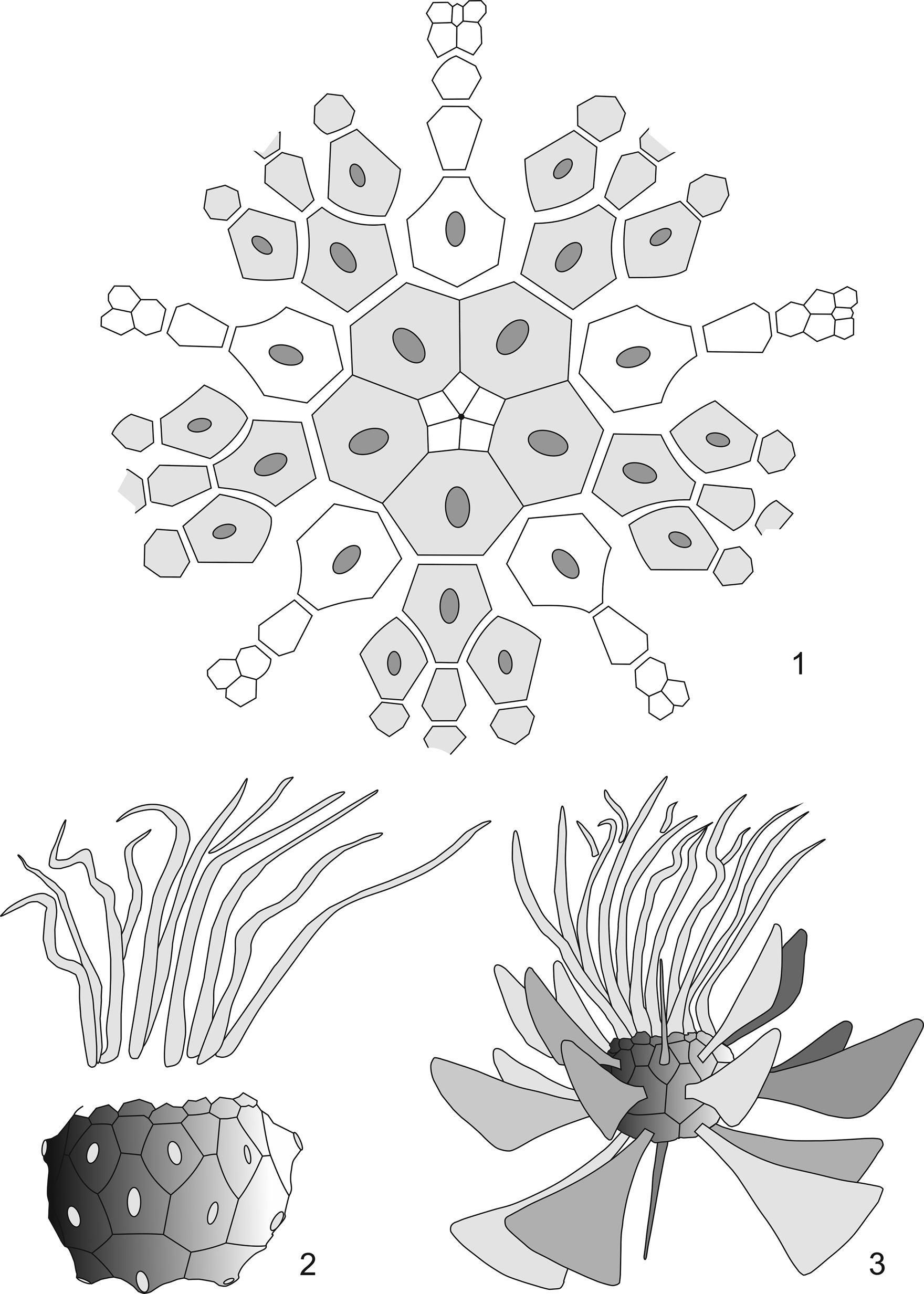
Figure 2. (1) Calyx plate diagram of Monstrocrinus securifer according to Schmidt (Reference Schmidt1941, p. 214, Fig. 60a). (2) Schematic reconstruction drawing of a Monstrocrinus calyx with “broken off floating spines” and “indicated arms” after Schmidt (Reference Schmidt1941, p. 214, Fig. 60b). (3) Reconstruction drawing of Monstrocrinus securifer, previously interpreted as a stemless crinoid, with “completely preserved floating spines” (after Schmidt, Reference Schmidt1941, p. 214, Fig. 60b); arms are simplified.
The type species, Monstrocrinus securifer Schmidt, Reference Schmidt1941, p. 215–217, was described as a large representative of the genus, which was characterized by vertically positioned, axe-shaped “floating spines.” The surface of the crown ossicles is unornamented. The specimens described by Schmidt (Reference Schmidt1941) are from the uppermost section of the Emsian Orthocrinus Beds in the area around Olpe; these are: (1) the former brickworks mine NW of Olpe-Lütringhausen (see Location and stratigraphy); (2) the abandoned “Waukemicke Quarry” (Olpe-Wauckemicke); (3) several fossil localities near Olpe-Oberveischede; and (4) the fossil localities at the “Siele Farmyard” (between Olpe-Lütringhausen and Olpe-Waukemicke). Schmidt also mentioned M. securifer remains from localities outside Olpe (Bad Laasphe–Feudingen in the Siegen–Wittgenstein district in North Rhine-Westphalia and the “Old Paper Mill Haiger Hut,” NE-Haiger in the northern part of the Lahn–Dill district in central Hesse).
The second species, M. granosus Schmidt, Reference Schmidt1941, p. 217–218, was described as a medium-sized species that is characterized by a granular ornamentation of the calyx, by single-pointed and forked “floating spines,” and by “two-lined arms.” The species occurs in the upper Emsian Mandeln Formation within the “iron-shed greywacke schist” in the “Old Municipality Quarry” at the northern Hauberg Hill in Dietzhölztal–Mandeln in the Lahn–Dill district in central Hesse, north-northeast of Dillenburg. The original material was discovered by the late geologist Gerhard Solle (1911–1981), who found the specimen in 1935. It is deposited in the Senckenberg Research Institute and Natural History Museum Frankfurt under catalog number SMF-XXIII-113a. Forty-three years later Ettensohn (Reference Ettensohn1984) discussed genus Monstrocrinus as a stemless crinoid genus that was passively moved by current.
Le Menn (Reference Le Menn1990) described an isolated plate with preserved appendages from the upper Emsian Pena Negra Formation northeast of the municipality of Loscos in the province of Teruel, Aragon, Spain (Le Menn, Reference Le Menn1990, p. 151–153). This specimen has a striking elk-antler-like morphology (Le Menn, Reference Le Menn1990, p. 152, figs. 1a, b; see Fig. 3.3). This extreme morphology was considered distinct from the “floating spines” of the type species, M. securifer, described by Schmidt (Reference Schmidt1941). Thus, Le Menn (Reference Le Menn1990) described the new species M. aliformis Le Menn, Reference Le Menn1990. However, new specimens of M. securifer described herein from the lowermost Eifelian of the Ahrdorf- and Hillesheim Eifel synclines demonstrate that the spine morphology of M. aliformis falls within the range of variation of the spine processes of M. securifer. Therefore, M. aliformis is treated herein as a subjective junior synonym of M. securifer. These new specimens also have an effect on our understanding of the evolutionary development of the “floating spines” as postulated by Le Menn (Reference Le Menn1990, p. 156, fig. 3). According to Le Menn's model, the tapering Acanthocrinus spines (Reference Le Menn1990, p. 156, fig. 3a) were first modified to form the forked Monstrocrinus granosus spines (Le Menn, Reference Le Menn1990, p. 156, fig. 3b; see Fig. 3.2), which then became the elk-antler-like spines of the species M. aliformis (Le Menn, Reference Le Menn1990, p. 156, fig. 3c, see Fig. 3.3), and finally adopted the “paddle-like floating prong shape” of the species M. securifer (Le Menn, Reference Le Menn1990, p. 156, fig. 3d). This development was said to have taken place between the lower to the uppermost Emsian. Beside the fact that the M. granosus spine shown in Schmidt (Reference Schmidt1941, p. 217, fig. 61, no. b2) has no particular taxonomic relevance, it was wrongly interpreted by Le Menn (Reference Le Menn1990, p. 156, fig. 3b). The prong-shaped extension (Fig. 3.2) was interpreted as pointing to the calyx plate (proximally). However, it is not a backward-pointing prong but a distally tapering bifurcation (cf., Schmidt, Reference Schmidt1941, p. 217). The proposed evolutionary model is furthermore refuted by the common occurrence of different forms of spinous calyx plate appendages of all Monstrocrinus species.
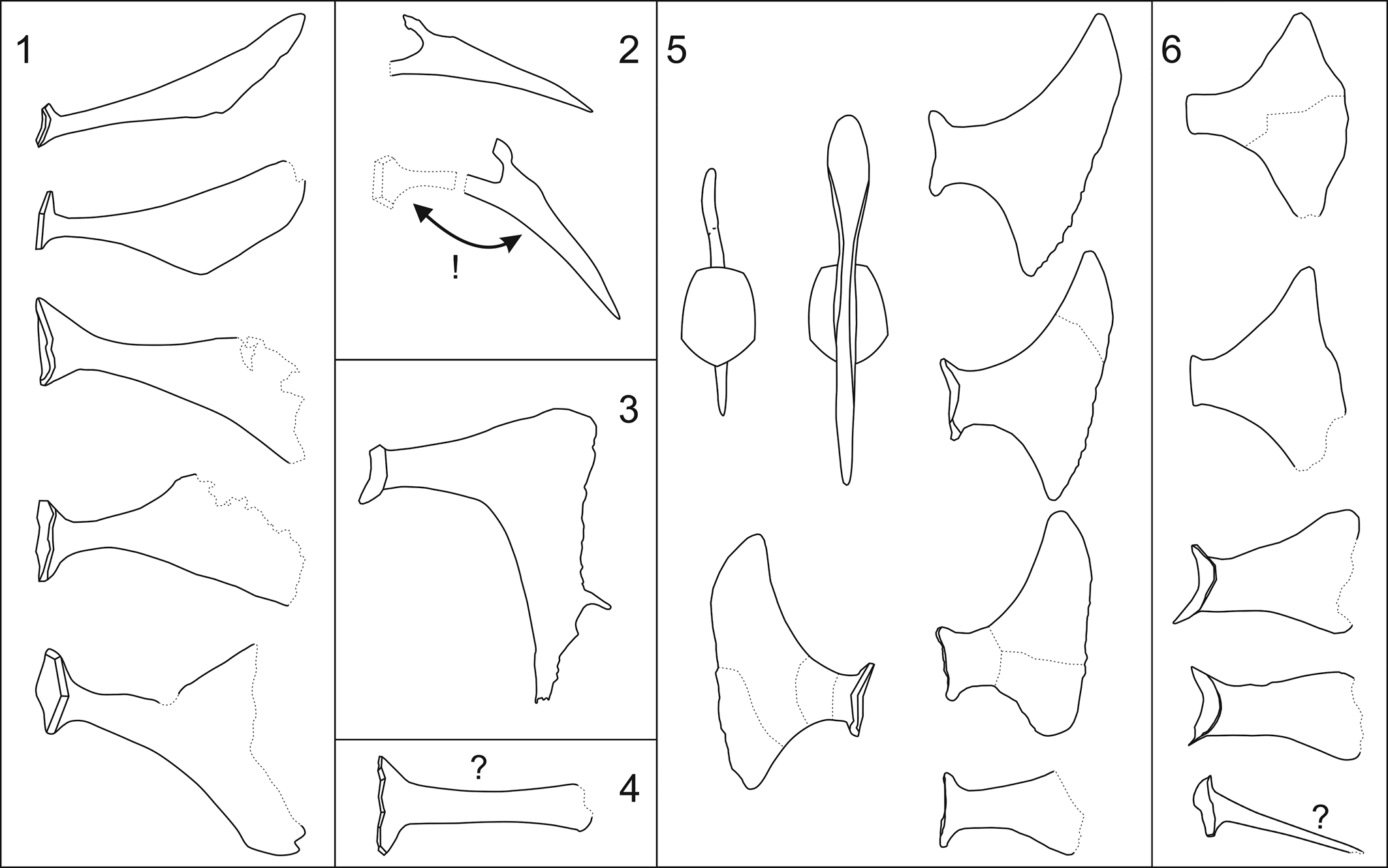
Figure 3. The isolated spiny calyx plate appendages of genus Monstrocrinus depicted in the literature. (1) Monstrocrinus securifer (after Schmidt, Reference Schmidt1941, pl. 16). (2) Monstrocrinus granosus (after Schmidt, Reference Schmidt1941, p. 217, fig. 61, no. b2), the upper figure shows a disarticulated distally forking spiny calyx plate appendage. Le Menn (Reference Le Menn1990, p. 156, fig. 3b) wrongly interpreted the prong-shaped extension as pointing proximally toward the calyx plate (see lower figure); however, the prong-shaped extension is not a backward-pointing prong, but a distally tapering bifurcation (see Schmidt, Reference Schmidt1941, p. 217). (3) “Monstrocrinus aliformis” (after Le Menn, Reference Le Menn1990, p. 152, fig. 1b). The species is interpreted in the present work as a subjective younger synonym of M. securifer. (4) “Monstrocrinus sp.” according to Le Menn (Reference Le Menn1990, p. 152, fig. 1d; 1997, pl. IV, fig. 14). The plate fragment cannot be clearly assigned to the genus Monstrocrinus. (5) Monstrocrinus incognitus (after Scheffler et al., Reference Scheffler, Dias-Da-Silva, Gama, Da Fonseca and Fernandes2011, p. 1191–1192, figs. 3, 4.1–4.6). (6) “Monstrocrinus securifer” (after Scheffler et al., Reference Scheffler, Fernandes and Da Fonseca2006, p. 238, fig. 3A–3E), either belonging to M. incognitus or to another species (further investigations are needed). The isolated spine shown at the bottom of the figure cannot be unequivocally assigned to the genus Monstrocrinus.
Furthermore, Le Menn (Reference Korn1990, p. 152, figs. 1c, d) figured two isolated calyx plates with narrow appendages as Monstrocrinus sp. from Emsian–Eifelian boundary of the lowermost part of the Chefar El Ahmar Formation of the Ougarta Mountains in the Sahara province of Béni Abbès in southwestern Algeria (also illustrated in Le Menn, Reference Le Menn1997, pl. 4, fig. 14). The plate fragment (Fig. 3.4) shown in Le Menn (Reference Le Menn1990, p. 152, figs. 1c, d) cannot be clearly assigned to the genus Monstrocrinus. Le Menn postulated that Monstrocrinus was stratigraphically restricted to the upper Emsian and that none of the species crossed the Lower–Middle Devonian boundary (Le Menn, Reference Le Menn1990, p. 149–150), which is refuted by specimens from the Lauch Formation of the lower Eifelian of the Eifel synclines and by the Monstrocrinus discoveries from the Eifelian strata of Brazil (see below).
Scheffler et al. (Reference Scheffler, Fernandes and Da Fonseca2006) described “Monstrocrinus securifer” from the middle Eifelian Maecuru Formation along the Maecuru River in the northwestern part of the State of Pará in Brazil. In addition to the characteristic “floating spines,” the authors also depicted an Acanthocrinus-like spine for the first time and assigned it to “M. securifer” (Scheffler et al., Reference Scheffler, Fernandes and Da Fonseca2006, p. 238, fig. 3E). This isolated calyx plate can only be assigned to the genus Monstrocrinus with reservation because numerous members of the family Rhodocrinitidae Roemer, Reference von Roemer and Bronn1855, have similar plates. Nevertheless, the articulated specimen described in the present work proves that Monstrocrinus had Acanthocrinus-like spines in addition to differently designed “floating spines.”
Scheffler et al. (Reference Scheffler, Fernandes and Da Fonseca2009) described additional remains of the calyx and crown of the genus Monstrocrinus and stated that each arm divided twice, resulting in four free arms per ray. Scheffler et al. (Reference Scheffler, Dias-Da-Silva, Gama, Da Fonseca and Fernandes2011) erected the new species Monstrocrinus incognitus from the middle to upper Eifelian Pimenteira Formation of the Parnaíba Basin in the State of Estado do Tocantins in Brazil. To date, this is the youngest species of the genus and was considered an intermediate form between “M. securifer” (with regard to the “floating spines” of Scheffler et al., Reference Scheffler, Fernandes and Da Fonseca2006, identified from the Maecuru Formation in Brazil) and “M. aliformis” from the Pena Negra formation of Spain (Scheffler et al., Reference Scheffler, Dias-Da-Silva, Gama, Da Fonseca and Fernandes2011, p. 1189). In contrast to these species, the “floating spines” should be characterized by short webs between the calyx plate attachment and the distal “wings” as well as by distally widened “wing appendages.” Note that the incomplete calyx plate in Scheffler et al. (Reference Scheffler, Dias-Da-Silva, Gama, Da Fonseca and Fernandes2011, p. 1192, fig. 4.2) has a longer web between the base of the plate and the distal “wing end” in comparison with the other examples illustrated (Scheffler et al., Reference Scheffler, Dias-Da-Silva, Gama, Da Fonseca and Fernandes2011, p. 1192, figs. 4.1, 4.3–4.6). This could indicate an insufficiently recorded morphological variability, as demonstrated for the type species in the present work. According to Scheffler et al. (Reference Scheffler, Dias-Da-Silva, Gama, Da Fonseca and Fernandes2011, p. 1192), the widened outer edge of the distal “floating spines” could have been useful for both an increase in the water turbulence in the direction of the arms in the parabolic filter position and for anchoring on the substrate (Scheffler et al., Reference Scheffler, Dias-Da-Silva, Gama, Da Fonseca and Fernandes2011, p. 1197).
The first two steps toward an understanding of Monstrocrinus is a standardized method for describing calyx spines, which then allows a means to describe spine variability more precisely on a single individual, from a single fossiliferous horizon, and intra- and interspecific variability. Figure 4 is a model for description of calyx spine shapes that builds on Scheffler et al. (Reference Scheffler, Dias-Da-Silva, Gama, Da Fonseca and Fernandes2011, fig. 3). The height and width of the entire calyx spine can be measured. The spine base has an outline shape and a height and width. Similarly, the spine neck (immediately abaxial from the spine base) has a cross-sectional shape and a height and width. The remainder of the calyx spine is referred to as the spine blade. This is the highly variable portion of the spine. The shape of spines is largely a function of different shapes of the upper edge, external edge, and lower edge (Fig. 4).
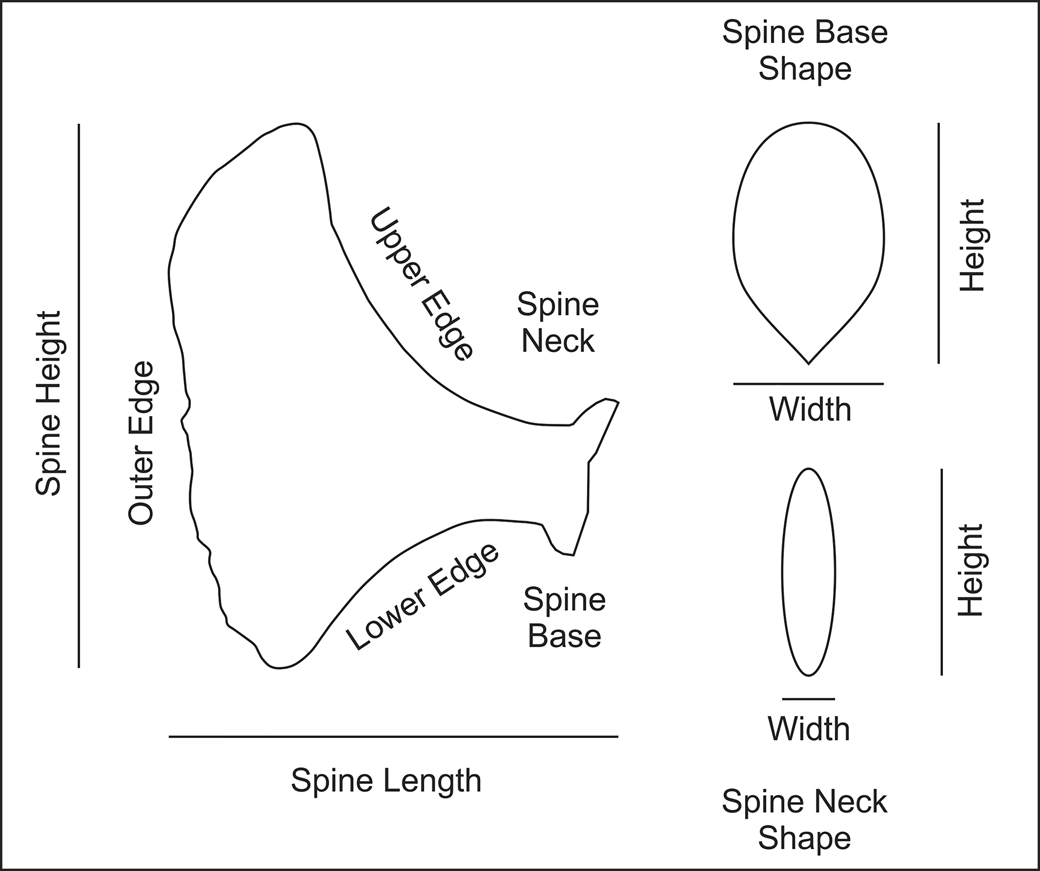
Figure 4. Morphological features of Monstrocrinus spines (modified from Scheffler et al., Reference Scheffler, Dias-Da-Silva, Gama, Da Fonseca and Fernandes2011, p. 1191, fig. 3).
There is a wide range of spine shapes in Monstrocrinus. Based on the examined fossil material it is now possible to distinguish eight standardized morphological spine types (Fig. 5). They are the following types:
-
Type 1: tapering, pointed spine.—Entire spine circular to subcircular in cross section; tapers to a point at terminus (Figs. 6.2, 9.2, 10, 12.1, 12.3, 12.4).
-
Type 2: blunt spine.—Entire spine higher than wide, elliptical in cross section; upper and lower edges either slightly expanding, parallel, or slightly narrowing; tapers to a flat, blunt terminus (Fig. 9.2).
-
Type 3: narrow paddle-shaped spine.—Neck long, much higher than wide, flat ellipse in cross section, gradually increases in height toward outer edge; upper and lower edges concave; entire spine expands gradually toward outer edge, lower edge longer, strongly asymmetrical; outer edge at an acute angle to horizontal; smooth or serrated (Fig. 8.1–8.3).
-
Type 4: broad paddle-shaped spine.—Neck short, much higher than wide, flat ellipse in cross section, rapidly increases in height toward outer edge; upper and lower edges sharply concave; entire spine expands rapidly in height, upper and lower edges very convex and expand at different rates yielding an asymmetrical spine; outer edge at a high angle to horizontal; straight or broadly convex; smooth or serrated. Type 4 spine shapes can be subdivided into two subtypes: (1) subtype 4a—upper edge longer and higher, spine asymmetrical upward; and (2) subtype 4b—lower edge longer and lower, spine asymmetrical downward (Fig. 8.4).
-
Type 5: flat, high fan.—Neck variable in shape, much higher than wide, flat ellipse in cross section; upper and lower edges parallel or broadly concave; entire spine shape variable; outer edge at high angle to horizontal, smooth. Type 5 spine shapes can be subdivided into two subtypes: (1) subtype 5a—neck short; upper and lower edges broadly concave, entire spine relatively short, gradually expanding in height toward outer edge; outer edge broadly convex; and subtype 5b—neck long, short, or undifferentiated; upper and lower edges very broadly concave or subparallel; entire spine gradually expanding from the neck, shortest at mid-spine position, or rectilinear; outer edge concave with or without upper and lower blunt extensions.
-
Type 6: bifurcating spine.—Neck long or short, much higher than wide, flat ellipse in cross section; entire fan gradually expanding in height toward outer edge, upper and lower edges broadly concave, bifurcates into two spines near or away from the spine base, entire spine symmetrical or asymmetrical; terminal extensions circular to subcircular in cross section, taper to a blunt point (Figs. 8.5, 12.2).
-
Type 7: forked spine.—Neck of various lengths, much higher than wide, flat ellipse in cross section; entire spine gradually expanding in height toward outer edge, upper and lower edges broadly concave; outer margin with various spine sizes and arrangements, spines may be long or short, may assume a serrated appearance, are arranged symmetrically or asymmetrically; spine cross section circular to elliptical. (Figs. 8.6, 8.7, 9.2, 10, 12.2, 12.4).
-
Type 8: strongly asymmetrical paddle shaped spine with spinose outer margin.—Neck short, much higher than wide, flat ellipse in cross section, rapidly increases in height toward outer edge; entire spine with upper and lower edges concave, expanding in height distally, lower edge more convex, longer, and lower; highly asymmetrical; outer edge at a high angle to horizontal, straight or broadly concave, serrated or with small spines (Fig. 8.8, 8.9).
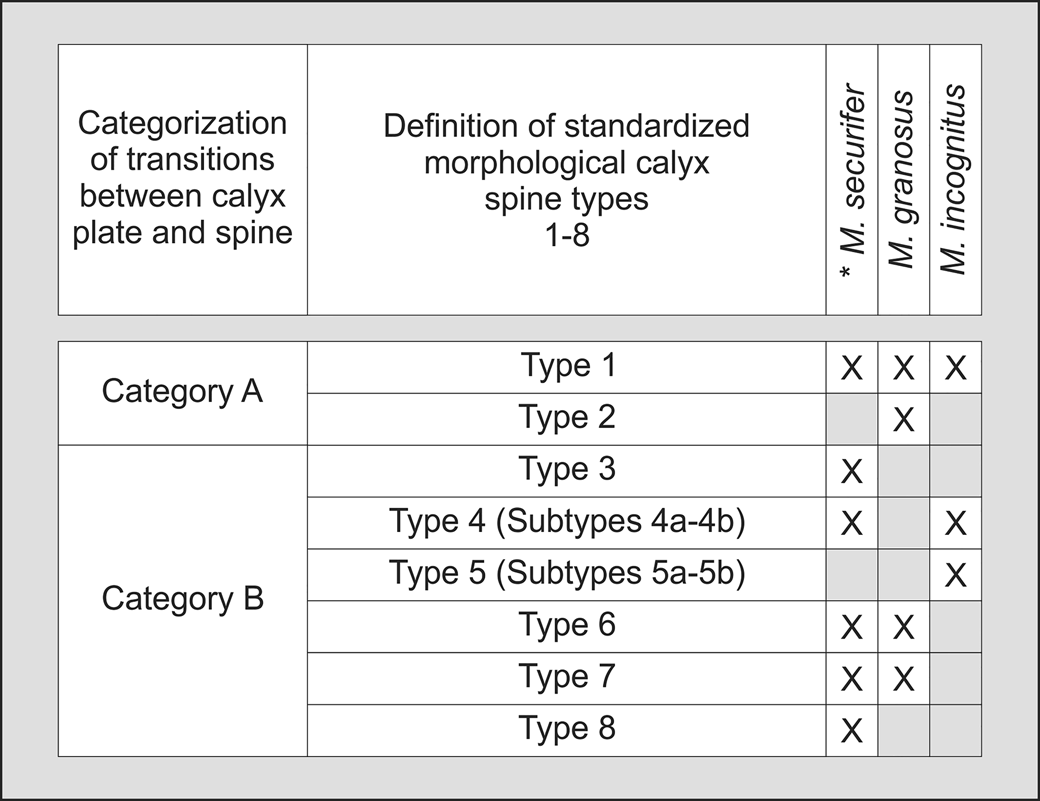
Figure 5. Categorization of transitions between calyx plate and spine and definition of standardized morphological calyx spine types and subtypes.
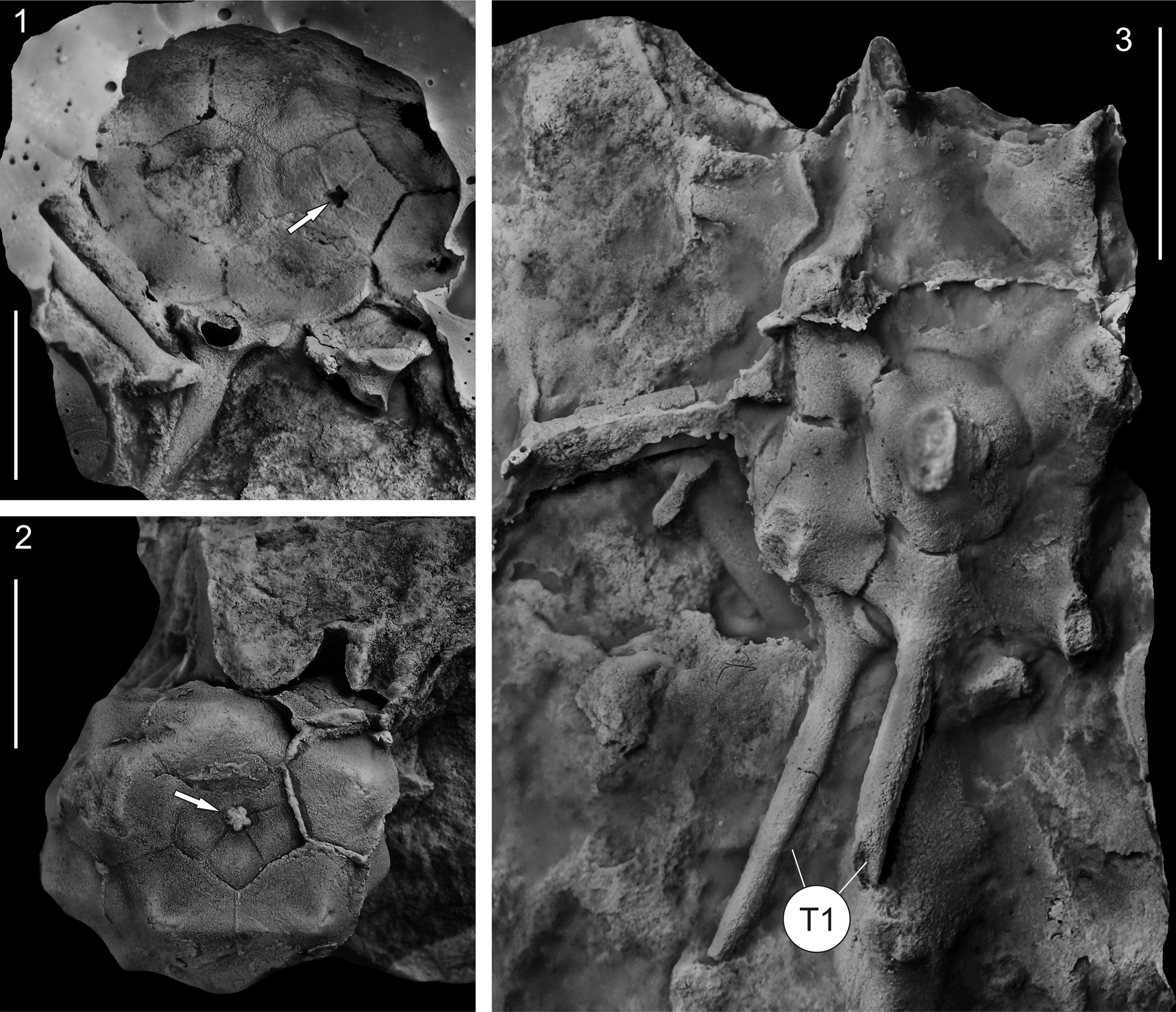
Figure 6. Recast holotype of Monstrocrinus securifer Schmidt, Reference Schmidt1941 (no. IGPB-SCHMIDT, W.E.-18a) from the uppermost section of the Emsian Orthocrinus Beds (Polygnathus costatus patulus Conodont Biozone) of the former brick factory in the Osterbachtal, northwest of Olpe-Lütringhausen (administrative district Arnsberg, Sauerland, North Rhine–Westphalia, Germany). (1, 2) Aboral view of the calyx, showing pentalobate axial canal (white arrows), which were not shown in the original illustration by Schmidt (Reference Schmidt1941, pl. 16, fig. 2b); (3) lateral view of the calyx with rudimentarily preserved type 1 spines (white T1). Scale bars = 1 cm.
These eight morphological types can be divided into two additional categories (Category A and B), because calyxes of Monstrocrinus have two different types of transitions between the proximally positioned calyx plate and the distally projecting end of the spine. Category A is a spine that truly seems to be completely a physical part of the calyx plate, whereas Category B appears to be an attachment onto the calyx plate, which is either positioned immediately proximal to a more planar calyx plate (e.g., in M. granosus), or somewhat higher, at the distal end of a funnel-shaped calyx plate projecting externally (e.g., in M. securifer; see Fig. 8.10–8.12). In M. granosus, the outer portion of the calyx plate is coarsely granulated, whereas the center of the plate has much less granulation. Here it seems that both the spine and spinous process were able to detach immediately from the flat and heavily granulated calyx plate. Category B possibly can be interpreted as a kind of “secondary spine.” Category A can be observed in morphological spine Types 1 and 2, and category B occurs in Types 3–8.
This standardized typing and categorization, which is distributed among the examined Monstrocrinus species as shown in Figure 5, enables the subsequent differential diagnosis of the individual Monstrocrinus species. Species-diagnostic characters include the relative size of the calyx, calyx plate sculpturing, presence, or absence of depressions at calyx plate triple junctions, spine categories, and spine types.
Monstrocrinus securifer Schmidt, Reference Schmidt1941
Figures 2, 3.1, 3.3, 6–8
- Reference Schmidt1941
Monstrocrinus securifer Schmidt, p. 214–217, fig. 60a–c, pl. 16, figs. 1–7.
- Reference Ubaghs, Moore and Teichert1978b
Monstrocrinus securifer; Ubaghs, p. T428, T429, fig. 237a, b.
- Reference Le Menn1990
Monstrocrinus securifer; Le Menn, p. 156, fig. 3d.
- Reference Le Menn1990
Monstrocrinus aliformis Le Menn, p. 151–153, 156, figs. 1a, b, 3c.
- Reference Hauser2012
Monstrocrinus securifer; Hauser, p. 1–4, text-figs. 4–6 (non text-fig 3).
- non Reference Hauser2012
Monstrocrinus granosus or Acanthocrinus sp.; Hauser, p. 4, text-fig. 6, no. 2a (= holotype of M. securifer).
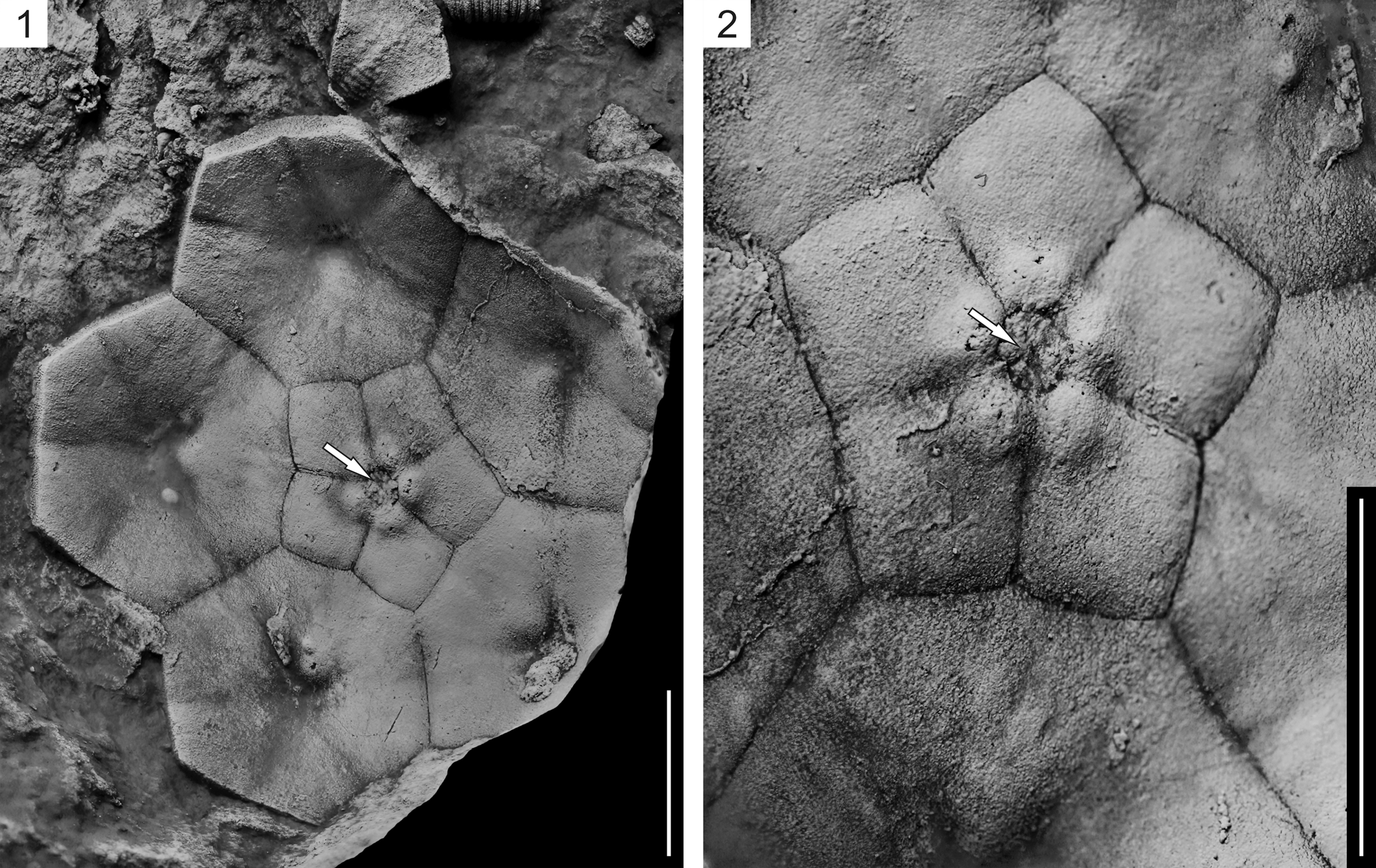
Figure 7. Recast paratype of Monstrocrinus securifer Schmidt, Reference Schmidt1941 (no IGPB-SCHMIDT, W.E.-18c) from the uppermost section of the Emsian Orthocrinus Beds (Polygnathus costatus patulus Conodont Biozone) of the former brick factory in the Osterbachtal, northwest of Olpe-Lütringhausen (administrative district Arnsberg, Sauerland, North Rhine–Westphalia, Germany). (1, 2) Internal aboral view of isolated base of a calyx, showing narrow stem axial canal impression (white arrows), which Schmidt mistakenly interpreted as closed (see Schmidt, Reference Schmidt1941, pl. 16, fig. 3). Scale bars = 1 cm.

Figure 8. Calcite-preserved spines (1–9) and isolated calyx plates (10–12) of Monstrocrinus securifer Schmidt, Reference Schmidt1941, from the Lauch Formation (Polygnathus costatus partitus to lowermost P. costatus costatus Conodont biozones) of the lower Eifelian of the Eifel synclines, Germany; (1, 4–12) are from the “Eschfelder Seifen,” southeast of Dollendorf, north-northwest of Leudersdorf, North Rhine–Westphalia; (2, 3) are from the northern Ahbach Valley, north-northeast of Üxheim–Ahütte, Rhineland–Palatine). (1–3) Type 3 spines (T3); (4) subtype 4b spine (T4b); (5) type 6 spine (T6); (6, 7) type 7 spines (T7); (8, 9) type 8 spines (T8); (10, 11) isolated calyx plates with “Category B” transitions between the proximally positioned calyx plates and the distally projecting end of the spines (“secondary spines,” not preserved). The spine attachment shows eight-shaped concavities. Scale bars = 1 cm.

Figure 9. Recast holotype of Monstrocrinus granosus Schmidt, Reference Schmidt1941 (no. SMF XXIII 113a) from the upper Emsian Mandeln Formation (Polygnathus serotinus Conodont Biozone) of Dietzhölztal–Mandeln in the Lahn–Dill district in central Hesse, north-northeast of Dillenburg. (1) Strongly disarticulated crown with longer part stem in matrix (until the present study, the stem was not interpreted as belonging to the individual); (2) detailed view, showing remains of spine types 1, 2, and 7 (T1, T2, T7) and granulated calyx plates. Scale bars = 3 cm in (1) and 1 cm in (2).
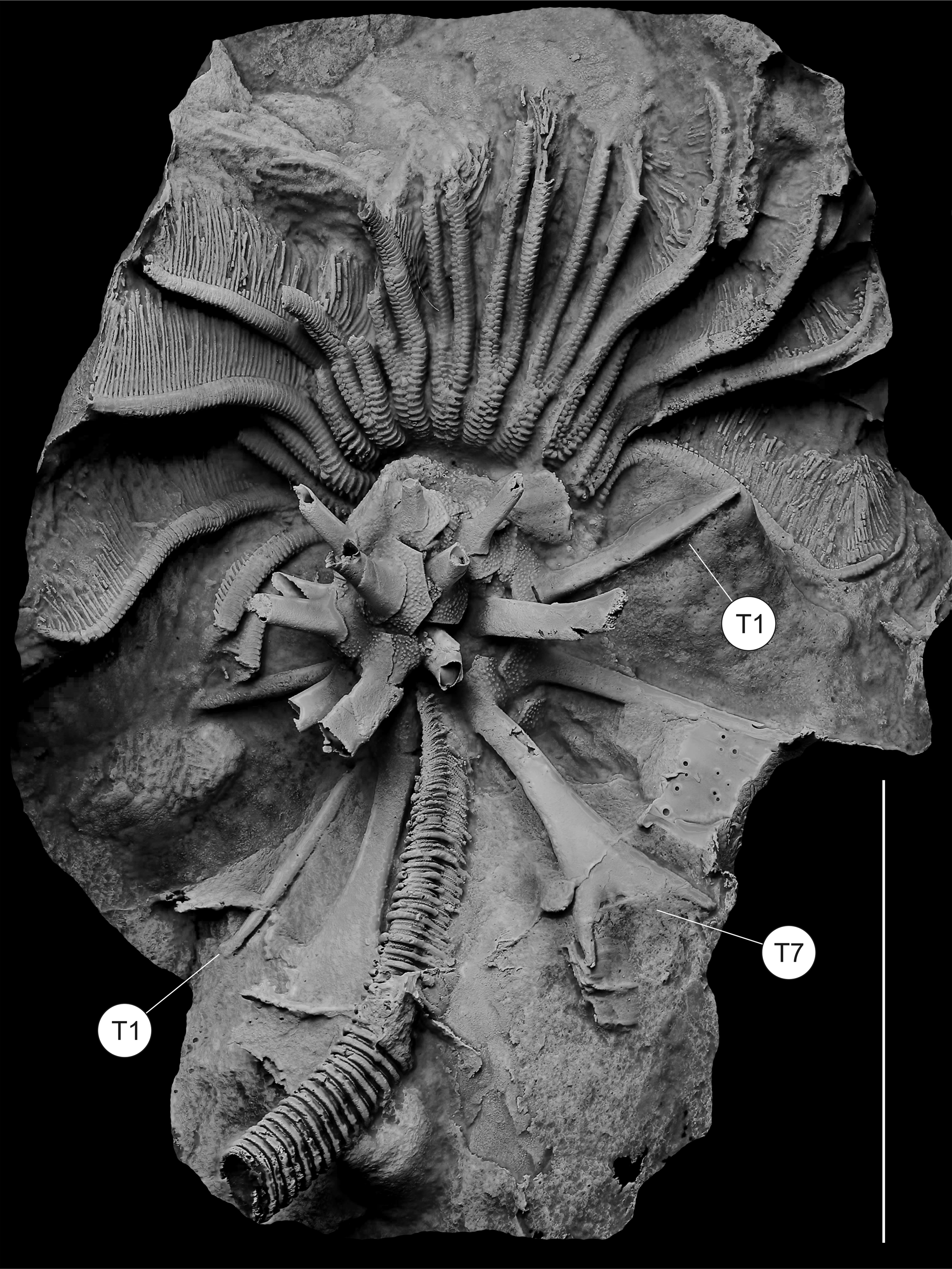
Figure 10. Lateral view of the cast of the most complete known Monstrocrinus (no. NHMMZ PWL 2021/6145-LS), M. granosus Schmidt, Reference Schmidt1941, from the uppermost section of the Emsian Orthocrinus Beds (Polygnathus costatus patulus Conodont Biozone) of the former brick factory in the Osterbachtal, northwest of Olpe-Lütringhausen (administrative district Arnsberg, Sauerland, North Rhine–Westphalia, Germany). The specimen in matrix shows a longer part of stem, partly articulated spines, especially spine types 1 and 7 (T1 and T7), which are well preserved, and brachials preserved. The specimen shows evidence of non-lethal predation of its arms in which the distal portions are narrower than the proximal arms. Similarly, in two rays on the right portion of the organism, the free arm bifurcation is lower than in other arms, and the arms above the bifurcation are narrower. Scale bar = 3 cm.
Types
Holotype: mold of an isolated calyx with rudimentarily preserved spines in matrix, no. IGPB-SCHMIDT, W.E.-18a (recast and re-illustrated in Fig. 6), original of Schmidt (Reference Schmidt1941, pl. 16, figs. 2a, b). Paratype: mold of an isolated base of a calyx in matrix, no. IGPB-SCHMIDT, W.E.-18c (recast and re-illustrated in Fig. 7), original of Schmidt (Reference Schmidt1941, pl. 16, fig. 3).
Emended diagnosis
Monstrocrinus with a relatively large calyx, smooth plate sculpturing, shallow depressions at calyx plate triple junctions, spine categories (A, spine that is physically part of a calyx plate; B, spine articulated with calyx plate); spine types (1, tapering point; 3, narrow paddle shaped; 4, broad paddled shape; 6, bifurcating; 7, forked; 8, strongly asymmetrical with spines on outer margin).
Occurrence
The locus typicus of M. securifer is the former brickworks mine (slopes north, east, and south of the former brick factory, at UTM: 51°02′22.73″N, 7°51′51.02″E) in the Osterbachtal on today's federal road B54, northwest of Olpe-Lütringhausen (administrative district Arnsberg, Sauerland, North Rhine-Westphalia, Germany). The classic site is located at the southeastern flank of the Middle Devonian Attendorn–Elspe Double Syncline in the transition to the Lower Devonian Siegen Anticline and stratigraphically belongs to the uppermost section of the Emsian (Lower Devonian) Orthocrinus Beds (Polygnathus costatus patulus Conodont Biozone). Additional specimens described by Schmidt (Reference Schmidt1941) are from the abandoned “Waukemicke Quarry” (Olpe-Wauckemicke); from several fossil localities near Olpe-Oberveischede; and from the fossil localities at the “Siele Farmyard” (between Olpe-Lütringhausen and Olpe-Waukemicke). Schmidt also mentioned M. securifer remains from localities outside Olpe (Bad Laasphe–Feudingen in the Siegen–Wittgenstein district in North Rhine–Westphalia and the “Old Paper Mill Haiger Hut,” NE-Haiger in the northern part of the Lahn–Dill district in central Hesse, Germany).
Additional material studied in the course of this work is from two outcrops within the Eifel synclines. The first locality is the northern embankment at the “Eschfelder Seifen” on the southwestern flank of the Middle Devonian Ahrdorf Syncline, in the transition area to the Lower Devonian Wiesbaum Anticline (southeast of Dollendorf, NNW of Leudersdorf, North Rhine–Westphalia; UTM: 50°21′45.62″N, 6°43′44.14″E). The second location is in the northern Ahbach Valley, on the north-northeastern flank of the Middle Devonian Hillesheim Syncline, in the transition area to the Lower Devonian Hoffeld Anticline (north-northeast of Üxheim–Ahütte, Rhineland–Palatinate; UTM: 50°20′56.25″N, 6°46′35.71″E) and stratigraphically belongs to the Lauch Formation (Polygnathus costatus partitus to lowermost P. costatus costatus Conodont biozones) of the lower Eifelian.
Because “M. aliformis” sensu Le Menn, Reference Le Menn1990 is treated herein as a subjective junior synonym of M. securifer, the species also occurs in the upper Emsian part of the Pena Negra Formation, northeast of the municipality of Loscos in the province of Teruel, Aragon, Spain.
Emended description
Calyx medium in size, medium to high globe-shaped calyx with a shallow basal concavity, calyx height:width ratio 1.25; depressions at calyx plate triple junctions present; plate sculpturing around spine bases smooth. Infrabasal plates confined to basal concavity, not visible in lateral view; five infrabasal plates. Five basal plates form part of calyx base and proximal portion of the calyx vertical wall. Radial circlet with ten plates (all radial plates alternate with interradial plates); presumably five radial plates, large, hexagonal, approximately as high as wide, with broad plications that connect to like plications on adjoining plates. Calyx plates with various long spines including spine categories A (spine that is physically part of a calyx plate) and B (spine articulated with calyx plate), and spine types 1 (tapering point), 3 (narrow paddle shaped), 4 (broad paddled shape), 6 (bifurcating), 7 (forked), and 8 (strongly asymmetrical with spines on outer margin). Proximal regular interradial plates large, approximately the same size as radial plates. First interradial plate higher than wide. CD interray not known. Fixed brachials present but the number is unknown, median ray ridges absent. Tegmen and free arms unknown. Column shape presumably circular, nodals wider than internodals, pentalobate lumen.
Remarks
Re-examination of the historical type material of Schmidt (Reference Schmidt1941) revealed that the base of the calyx of Monstrocrinus securifer has a stem attachment with a central pentalobate axial canal, which confirms that the M. securifer crown was attached to a column as an adult (see Figs. 6.1, 6.3, 7). Thus, a free-living life mode of a passively rolling or drifting crinoid is refuted, and the type species is reinterpreted as an attached, stalked crinoid.
The longest spine of M. securifer examined by us is 7.5 cm long, with the base missing (Fig. 8.8). So, it was originally about 8.0 cm long. Our measurements show that the largest-known calyxes (without preserved spines) have at least 4.0 cm in equatorial cross-section. If this calyx is now placed between two of our approximately 8.0-cm long spines on the equatorial plane, the result is a maximum total diameter of a calculated adult calyx of 20.0 cm. This value would probably be somewhat lower (~15 cm) if the spines were slightly inclined in the aboral direction. The ratio of the maximum calyx diameter with preserved spines to maximum crown length (calyx with arms) is at least 1:1. On this basis, we calculated total crown height of 40.0 cm (or 30.0 cm with slightly aborally inclined spines), as well as a truly “monstrous” fan when the arms were deployed in a feeding position.
Monstrocrinus securifer is distinguished from other species because it has a relatively large calyx, smooth calyx plate sculpturing, and shallow depressions at calyx plate triple junctions. It also had spine categories A (spines that are physically part of a calyx plate) and B (spine articulated with calyx plate), and the following spine types: (1) tapering point, (3) narrow paddle shaped, (4) broad paddled shape, (6) bifurcating, (7) forked, and (8) strongly asymmetrical with spines on outer margin. In contrast, M. granosus has a relatively small calyx, fine granulose calyx plate sculpturing, and shallow depressions at calyx plate triple junctions are absent. It also had spine categories A (spines that are physically part of a calyx plate) and B (spines articulated with calyx plate), and the following spine types: (1) tapering point, (2) blunt spine, (6) bifurcating, and (7) forked. The morphology of the calyx in M. incognitus is poorly preserved, but diagnostic characters that are known include smooth calyx plate sculpturing, spine categories A (spine that is physically part of a calyx plate) and B (spines articulated with calyx plate), and spine types (1) tapering point, (4) broad paddle-shaped spine, and (5) flat high spine.
Monstrocrinus granosus Schmidt, Reference Schmidt1941
Figures 3.2, 9–12
- Reference Schmidt1941
Monstrocrinus granosus Schmidt, p. 217–218, text-figs. 61a, 61b1, 61b2, 61c, 61d, 61e1, 61e2.
- Reference Le Menn1990
Monstrocrinus granosus; Le Menn, p. 151, 153–154, 156 (non fig. 3b).

Figure 11. Digitally cropped view of the specimen shown in Figure 10. Scale bar = 3 cm.
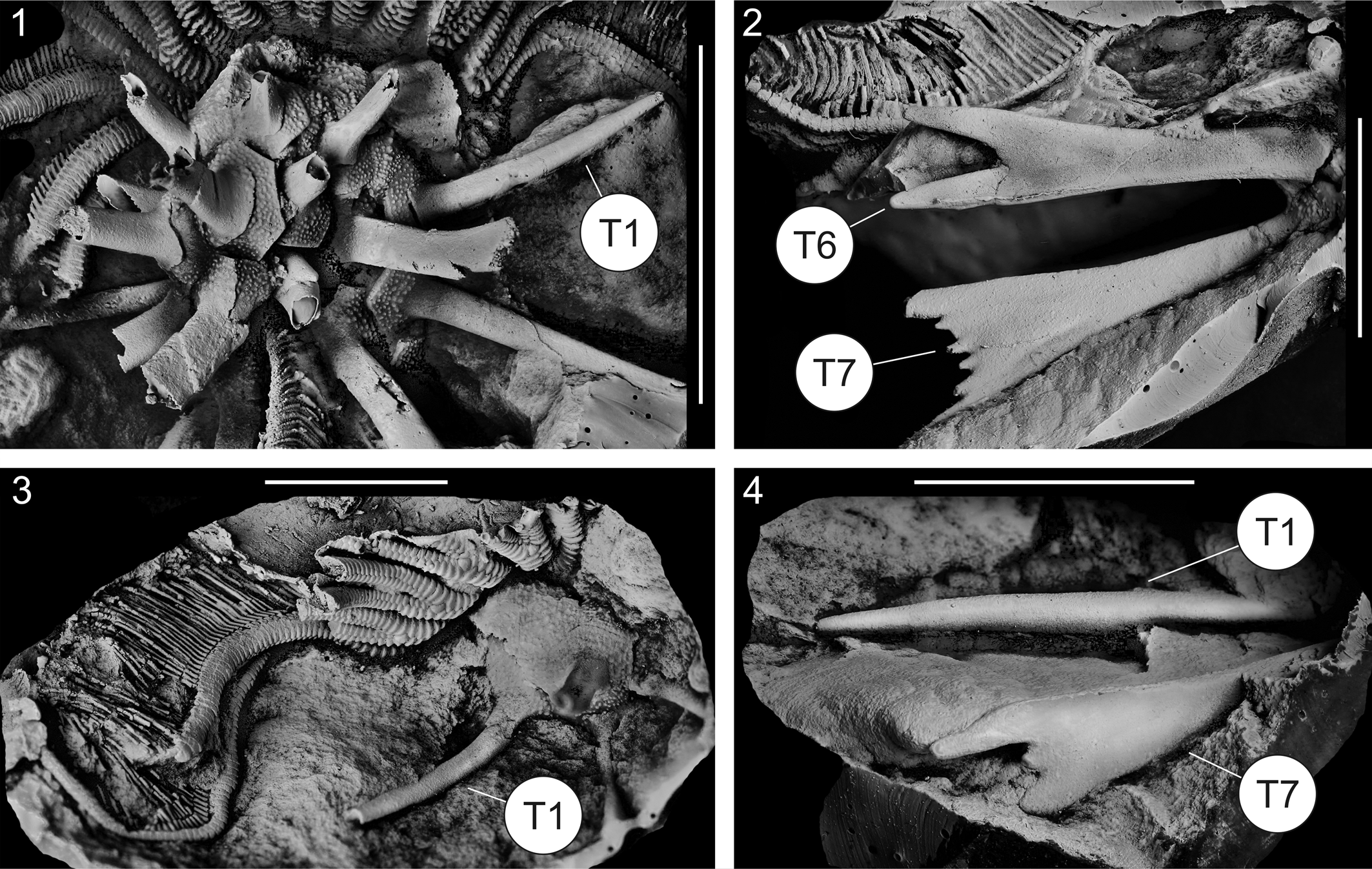
Figure 12. (1–4) Detail views of the specimen from Figures 10 and 11. (1) Lateral view of the spined calyx with well-preserved spine type 1 (T1). (2–4) Casts of the counter plates: (2) lateral view of two well-preserved spines of types 6 and 7 (T6, T7); (3) lateral view of part of the arms and calyx, showing spine type 1 (T1); (4) lateral view of spine types 1 and 7 (T1, T7). Scale bars = 2 cm in (1) and 1 cm in (2–4).
Holotype
Mold of a disarticulated crown with longer stem part in matrix, no. SMF XXIII 113a (recast and re-illustrated in Fig. 9), original of Schmidt (Reference Schmidt1941, p. 217, text-figs. 61.a, 61.b1, 61.b2, 61.c, 61.d, 61.e1, 61.e2).
Emended diagnosis
Monstrocrinus with a relatively small calyx; fine granulose calyx plate sculpturing; shallow depressions at calyx plate triple junctions absent; spine categories (A) spine that is physically part of a calyx plate and (B) spine articulated with calyx plate; and spine types (1) tapering point, (2) blunt spine, (6) bifurcating, and (7) forked.
Occurrence
The species occurs in the upper Emsian Mandeln Formation (Polygnathus serotinus Conodont Biozone) within the “iron-shed greywacke schist” in the “Old Municipality Quarry” at the northern Hauberg Hill in Dietzhölztal–Mandeln in the Lahn–Dill district in central Hesse (Germany), north northeast of Dillenburg (type locality at UTM: 50°51′22.52″N, 8°20′51.41″E). The examined material is from the locus typicus of the type species M. securifer, the former brickworks mine (slopes north, east, and south of the former brick factory, at UTM: 51°02′22.73″N, 7°51′51.02″E) in the Osterbachtal on today's federal road B54, northwest of Olpe-Lütringhausen (administrative district Arnsberg, Sauerland, North Rhine–Westphalia, Germany). The classic site is located at the southeastern flank of the Middle Devonian Attendorn–Elspe Double Syncline in the transition to the Lower Devonian Siegen Anticline and stratigraphically belongs to the uppermost section of the Emsian (Lower Devonian) Orthocrinus Beds (Polygnathus costatus patulus Conodont Biozone).
Emended description
Calyx small in size, medium globe-shaped calyx with a shallow basal concavity, calyx height:width ratio 1.25; depressions at calyx plate triple junctions absent; finely granulose calyx plate sculpturing on plates and may be on adaxial portion of spines. Infrabasal plates confined to basal concavity, not visible in lateral view; presumably five infrabasal plates, pentagonal. Five basal plates form part of calyx base and proximal portion of the calyx vertical wall. Radial circlet with ten plates (all radial plates alternate with interradial plates); radial plates large, presumably five, hexagonal, approximately as high as wide, lacking broad folds. Calyx plates with long spines on at least basal plates, radial plates, proximal interradial plates, and some fixed brachials; spine categories (A) spine that is physically part of a calyx plate and (B) spine articulated with calyx plate; spine types (1) tapering point, (2) blunt spine, (6) bifurcating, and (7) forked. Proximal regular interradial plates large, hexagonal (more distal regular interray plates not known). First interradial plate in regular interrays approximately the same size as radial plates and as high as wide. CD interray not known. Fixed brachials present but the number is unknown, median ray ridges absent. Tegmen unknown. Free arms presumably 20; high; one isotomous division; initially project obliquely downward, then abaxially, and finally obliquely upward. Proximal free brachials subrectangular, free brachials after division sharply convex, all brachials chisel biserial. Pinnules very long and slender, proximal pinnulars rectangular, higher than wide. Column circular, pentalobate lumen; nodal–internodal pattern N1; nodals wider than internodals. Proportions of different features on columnal facets: 12% narrow crenularium, 52% aereola, 8% perilumen, 28% lumen.
Remarks
After recasting the holotype of Monstrocrinus granosus Schmidt, Reference Schmidt1941 (no. SMF XXIII 113a), which was kindly provided by the Senckenberg Research Institute and Natural History Museum Frankfurt on the Main (Germany), important morphological characters came to light that were not recorded in the original description (compare Schmidt, Reference Schmidt1941, p. 217, figs. 61a–e with our Fig. 9). The holotype of M. granosus is far more complete than previously reported. It is a highly disarticulated crown embedded together with an associated long part of the stem (Fig. 9). In addition, further details of the spine morphology could be recorded. The previously unappreciated knowledge about the spine morphology led to an incorrect interpretation of a granosus spine by Le Menn (Reference Le Menn1990, p. 156, Fig. 3b). The latter author understood the prong-shaped extension of Schmidt's fig. 61b2 (1941, p. 217) as pointing proximally toward the calyx plate. In fact, it is not a backward-pointing prong, but a distally tapering bifurcation (see Schmidt, Reference Schmidt1941, p. 217).
Whereas the holotype of M. granosus was found within the upper Emsian Mandeln Formation of the “Old Municipality Quarry” (Paleontological Ground Monument of the Federal State of Hesse, no. “Mandeln 991″) at the northern Hauberg Hill in Dietzhölztal–Mandeln in the Lahn–Dill district in central Hesse (Germany), the material examined by us comes from the uppermost section of the Emsian (Lower Devonian) Orthocrinus Beds of the M. securifer type locality, the former brickworks mine northwest of Olpe-Lütringhausen (administrative district Arnsberg, Sauerland, North Rhine-Westphalia, Germany). To date, this locality in Germany is the only fossil site where both species co-occur.
It should be noted that the endoskeletons of M. granosus are consistently smaller in size than in the comparatively larger type species M. securifer. Monstrocrinus granosus is compared to other species of Monstrocrinus in the remarks section of M. securifer.
Noteworthy is that the distal portion of some distal arms are of variable widths. Also, the position of the free arm division is variable. One hypothesis is that these differences among free arms were a consequence of non-lethal predation on this specimen.
Monstrocrinus incognitus Scheffler et al., Reference Scheffler, Dias-Da-Silva, Gama, Da Fonseca and Fernandes2011
Figure 3.5, 3.6(?)
- Reference Scheffler, Dias-Da-Silva, Gama, Da Fonseca and Fernandes2011
Monstrocrinus incognitus Scheffler et al., p. 1189–1192, fig. 5.1–5.6.
Types
The type material of M. incognitus is housed in the paleontological collection of the Departamento de Geologia e Paleontologia do Museu Nacional, Universidade Federal do Rio de Janeiro (UFRJ), Brazil, under institutional abbreviation no. MN8277-Id. (holotype) and nos. MN8277-Ia, and MN8277-Ib (paratypes).
Emended diagnosis
Monstrocrinus with calyx size unknown; smooth calyx plate sculpturing; calyx plate triple junctions character unknown; spine categories (A) spine that is physically part of a calyx plate and (B) spine articulated with calyx plate; and spine types (1) tapering point, (4), broad paddle-shaped, (5), flat, high fan.
Occurrence
Monstrocrinus incognitus was reported from the western edge of the Parnaíba Basin from three outcrops (Fazenda Encantada II, Estância Cantilena, and Mirante Taquaruçu) in the district of Taquaruçu (Municipality de Palmas, state of Tocantins, Brazil), and, according to Scheffler et al. (Reference Scheffler, Dias-Da-Silva, Gama, Da Fonseca and Fernandes2011), occurs in outcrops positioned near the base of the middle to upper Eifelian Pimenteira Formation.
Remarks
Monstrocrinus incognitus is characterized in particular by the distinctive type 4a spines, which have a shorter length and a generally shorter and taller neck compared to those in M. granosus and M. securifer. This feature clearly justifies designation of a distinct species.
Scheffler et al. (Reference Scheffler, Fernandes and Da Fonseca2006, p. 238, fig. 3A–3E) described “Monstrocrinus securifer” from the middle Eifelian Maecuru Formation along the Maecuru River in the northwestern part of the State of Pará in Brazil. In our opinion, this material belongs either to M. incognitus or to another species of Monstrocrinus (compare Fig. 3.5 and 3.6 of the present study). Therefore, it could be possible that the species diversity of Monstrocrinus in Brazil is higher than currently known (further investigations are necessary).
Results and discussion
Schmidt (Reference Schmidt1941) described the base of the Monstrocrinus calyx from a specimen that is primarily an internal mold of articulated infrabasal and basal plate circlets (a mold of this specimen is illustrated in Fig. 7). The original interpretation of Monstrocrinus was that this crinoid lacked a column as an adult; however, the calyx–proximal columnal attachment would be visible on the opposite surface of the infrabasal circlet. The most complete, new specimen has a long segment of column projecting outward from the basal concavity, which confirms that the Monstrocrinus crown was attached to a column as an adult. Thus, a free-living life mode of a passively rolling or drifting crinoid (Schmidt, Reference Schmidt1941; Ettensohn, Reference Ettensohn1984; Le Menn, Reference Le Menn1990; Hauser, Reference Hauser2012) is refuted. Instead, the life mode Monstrocrinus should be interpreted as an attached, stalked echinoderm. Therefore, the previous interpretations of the calyx spines as floating spines, crawling spines, or skeletal elements resting on the sea floor must be reinterpreted, because the spines were most likely lifted above the bottom substrate, as was the entire crown.
Three functional hypotheses for the long spines are: 1, antipredatory; 2, modification of currents to enhance feeding; and 3, stability of the crown in a current. We regard the antipredatory interpretation for the spines to be the most probable, which would be consistent with increasing predation pressure during the Mid Paleozoic Marine Revolution (Meyer and Ausich, Reference Meyer, Ausich, Tevesz and McCall1983; Signor and Brett, Reference Signor and Brett1984; Brett and Walker, Reference Brett, Walker, Kowalewski and Kelley2002; Baumiller and Gahn, Reference Baumiller, Gahn, Kowalewski and Kelley2004, Reference Baumiller and Gahn2012; Gahn and Baumiller, Reference Gahn and Baumiller2005, Reference Gahn and Baumiller2010). Further evidence of non-lethal predation on Monstrocrinus is the occurrence of arms in which the distal portion of the arms are narrower than the proximal arms. This condition occurs in several arms (Fig. 10) and is consistent with non-lethal predation. Similarly, in two rays on the right portion of the organism, the free arm bifurcation is lower than in other arms and the arms above the bifurcations are narrower. This is also consistent with regeneration following non-lethal predation (see Meyer and Ausich, Reference Meyer, Ausich, Tevesz and McCall1983; Signor and Brett, Reference Signor and Brett1984; Brett and Walker, Reference Brett, Walker, Kowalewski and Kelley2002; Baumiller and Gahn, Reference Baumiller, Gahn, Kowalewski and Kelley2004, Reference Baumiller and Gahn2012; Gahn and Baumiller, Reference Gahn and Baumiller2005, Reference Gahn and Baumiller2010).
Spines producing eddying that may have enhanced feeding is a second hypothesis for the spines. In this case, perhaps eddying of aboral–oral flow of water through the spines slowed water that passed into the arms and directed a larger sample of water than only straight-line currents (Fig. 13).
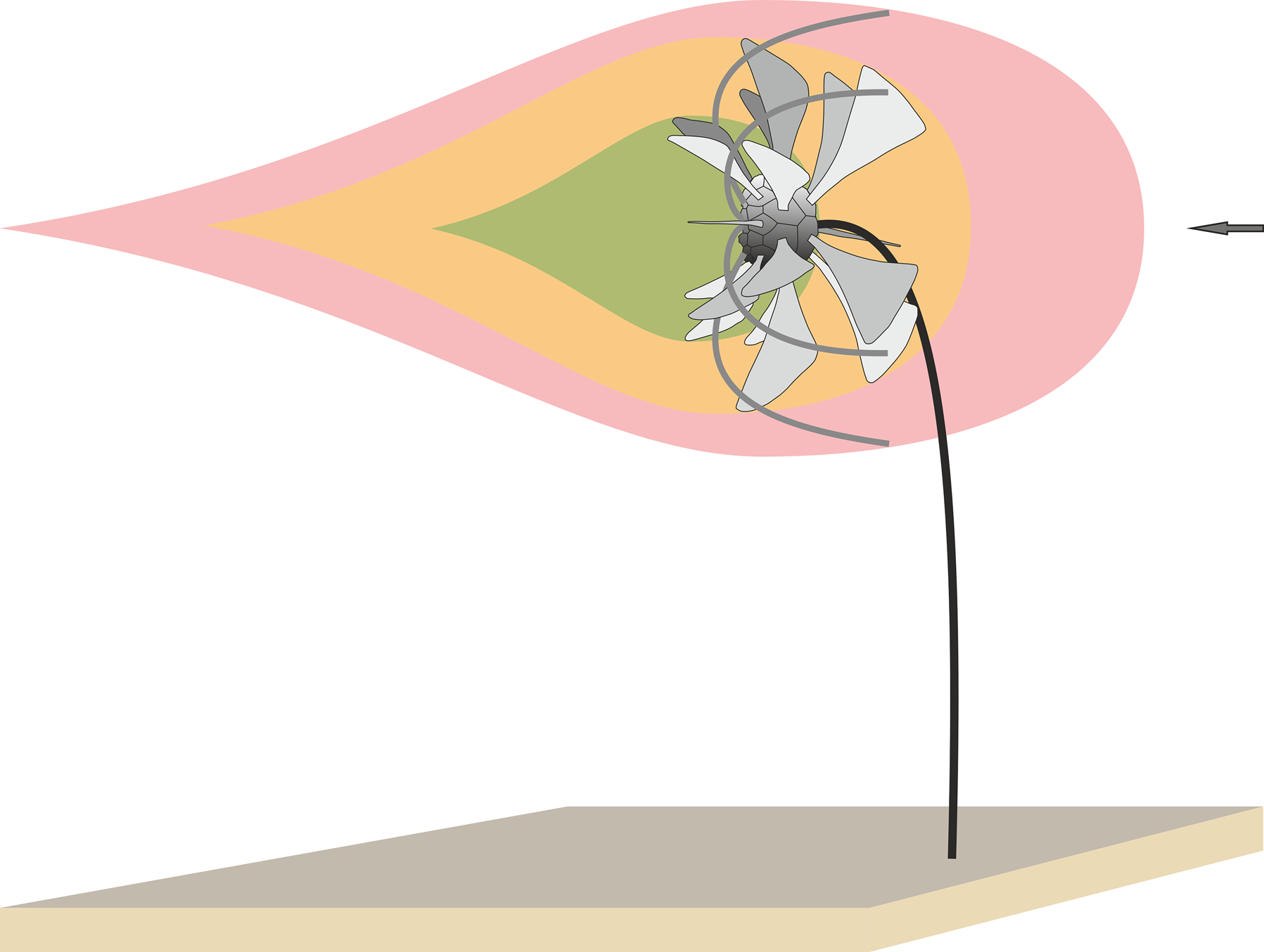
Figure 13. Schematic reconstruction of Monstrocrinus in feeding posture (based on the model of Pterotocrinus depressus by Baumiller and Plotnick, Reference Baumiller and Plotnick1989, p. 323, fig. 10); note arms are not in this reconstruction. The arrow indicates the current direction. In this model, the spines of Monstrocrinus serve as stabilizing fins or rudders, allowing the passive maintenance of an efficient feeding posture in moving water. While the crown is lifted into the water column by the stem and the arms were stretched out in the filtration fan feeding position, an aboral-to-oral water flow through the spines create a zone of slower recirculating water downstream toward the arms and the oral side of the calyx. The zone shown in pink, yellow, and green indicate fast, medium, and slower recirculating water downstream.
It is also possible that the spines provided rotational stability for the crown, similar to that recognized by Baumiller and Plotnick (Reference Baumiller and Plotnick1989) in a study on the function of wing plates of the Mississippian camerate crinoid Pterotocrinus depressus Lyon and Casseday, Reference Lyon and Casseday1860. However, the fact that spine placement, size, and shape varied around the calyx may not support this third hypothesis as an adaptive strategy.
Acknowledgments
We are especially grateful to R. Bersch (Dortmund) who discovered specimen NHMMZ PWL 2021/6145-LS from Olpe about 40 years ago and donated it to one of us (MJP). E. Schindler and M. Ricker (Senckenberg Research Institute and Natural History Museum Frankfurt on the Main, Germany) kindly provided the holotype of Monstrocrinus granosus for reexamination. The same thanks go to G. Heumann (Goldfuß-Museum, Institute for Geosciences, Rhenish Friedrich Wilhelm University of Bonn, Germany), who made the type material of M. securifer available to us. We thank S.M. Scheffler (Federal University of Rio de Janeiro, UFRJ, Brazil) for his willingness to discuss and for important stratigraphic and taxonomic information on the Brazilian Monstrocrinus. We are curious to learn further insights and details of Brazilian occurrences. We thank U. Hein (Solingen), and H.-P. Hein (Wermelskirchen) for discovery and donation of several studied crinoids, as well as for numerous important comments. We thank S. Zamora (editor, Journal of Paleontology) and P. Gorzelak (associate editor, Journal of Paleontology), and F. J. Gahn, and one anonymous reviewer for their reviews that greatly enhanced this manuscript.
Declaration of competing interest
There is no conflict of interest with any author.














 Open access
Open access