



Introduction
Chicken (Gallus gallus domesticus) production, both egg and meat, has steadily increased over the past decade, with 76.7 million tonnes of eggs produced in 2018 (van Horne Reference van Horne2018) and about 120 million tonnes of meat produced in 2019 (Windhorst Reference Windhorst2017). Global demand is only expected to increase further, which is also expected to include that for free-range, pasture-raised, and organic options. In the European Union (EU), free-range and organic poultry systems account for 18.7% of operations (Windhorst Reference Windhorst2017), and in the United States (US) 37.8% of egg layer chickens are currently estimated to now be in some form of a cage-free environment while 15% are raised in organic settings with outdoor access. An additional unknown number of birds are also free-range or pasture-raised (Ibarburu Reference Ibarburu2016). Collectively, these patterns indicate ongoing shifts toward alternative production systems in the coming years.
The roundworm Ascaridia galli is an internal parasite of galliform birds with a global distribution. It can cause production losses and clinical symptoms including weight loss, damage to the intestinal mucosa, intestinal impaction, alterations in hormone levels, and death. Additionally, there have also been reports of A. galli worms appearing within chicken eggs, which, although not a health concern for humans, may be a source of consumer complaints and possibly discourage people from consuming eggs (Piergili Fioretti et al. Reference Piergili Fioretti, Veronesi, Diaferia, Pia Franciosini and Casagrande Proietti2005; Höglund & Jansson Reference Höglund and Jansson2011; Bautista-Vanegas et al. Reference Bautista-Vanegas, Esteban-Mendoza and Cala-Delgado2023). Due to regulatory changes banning the use of traditional battery cages for laying hens in the EU (Appleby Reference Appleby2003) and increased demand for organic and free-range options, the prevalence of A. galli infection has risen. In Denmark, for example, 63.8% of free-range/organic systems contained A. galli versus only 5% of battery cage systems (Permin et al. Reference Permin, Bisgaard, Frandsen, Pearman, Kold and Nansen1999). A recent meta-analysis also indicates a higher prevalence of A. galli in backyard and free-range systems versus cage systems, as well as an increase in prevalence over time in developed countries (Shifaw et al. Reference Shifaw, Feyera, Walkden-Brown, Sharpe, Elliott and Ruhnke2021).
Taken together, the increased demand for organic, free-range, and pasture-raised options and the inherently higher A. galli infection rate in these systems is an alarming prospect. Anthelmintics cannot be used to treat infections in organic systems, and in non-organic systems, the only anthelmintics available for treatment of A. galli infections both in the US and the EU are benzimidazoles. Two close relatives of A. galli that infect turkeys, A. dissimilis and Heterakis gallinarum, have both developed resistance to fenbendazole (Collins et al. Reference Collins, Jordan, Baldwin, Hebron, Paras, Vidyashankar and Kaplan2019; Collins et al. Reference Collins, Jordan, Vidyashankar, Bishop and Kaplan2022), and decreased efficacy of benzimidazoles against A. galli has also been documented (Yazwinski et al. Reference Yazwinski, Tucker, Wray, Jones and Clark2013).
Given the dearth of available drugs, development of anthelmintic resistance, and potential health risks to the birds, a targeted approach to anthelmintic treatment in chickens is necessary. Faecal egg counts (FECs) are used in equine, small ruminant, and cattle industries to restrict treatment only to cases where it is necessary, whether it be with individuals or pooled samples exceeding a chosen eggs per gram (EPG) threshold in faeces. When a treatment threshold of 200 EPG for pooled samples was chosen in chickens, not only were FECs lower for the targeted treatment group, but overall worm burdens were also reduced (Tarbiat et al. Reference Tarbiat, Jansson, Tydén and Höglund2016). Treatment should only be administered based on information from FECs that are repeated every eight weeks, enabling a sustainable system for anthelmintic treatment in these birds (Tarbiat et al. Reference Tarbiat, Jansson and Höglund2022). In addition, using FECs affords the opportunity to monitor anthelmintic efficacy using faecal egg count reduction tests (FECRTs); in organic systems FECRTs can also be used to assess the effect of natural treatment methods.
Traditionally, FECs are performed using manual methods (Nielsen Reference Nielsen2021), most commonly the McMaster (MM) method (Gordon & Whitlock Reference Gordon and Whitlock1939) for agricultural animals. Manual methods are subject to variation due to the human element involved in sample preparation and counting (Went et al. Reference Went, Scare, Steuer and Nielsen2018; Slusarewicz et al. Reference Slusarewicz, Slusarewicz and Nielsen2019; Cain et al. Reference Cain, Peters, Suri, Roher, Rutledge and Nielsen2021) and often use a subsample of a subsample, meaning that a small proportion of the overall sample is examined. Homogeneity is assumed, despite the large variation that occurs between subsamples from the same animal as well as between animals (Nielsen Reference Nielsen2021). Manual methods also require extensive training of analysts (Cain et al. Reference Cain, Peters, Suri, Roher, Rutledge and Nielsen2021) and often take more than ten minutes per sample to perform. Recently, automated FEC methods have been introduced to the market, one of which is the Parasight System (PS; Parasight System, Inc., Lexington, Kentucky). This method is unique in that it fluorescently stains parasite eggs via the carbohydrate polymer chitin, which is a constituent of the shells of nematode ova (Slusarewicz et al. Reference Slusarewicz, Pagano, Mills, Popa, Chow, Mendenhall, Rodgers and Nielsen2016), including those of A. galli (Figure 1).

Figure 1. Representative images of fluorescently-stained Ascaridia galli eggs under A) brightfield and B) fluorescence illumination at 100x.
In previous studies, the system has been validated for the quantification of parasite eggs in horse (Scare et al. Reference Scare, Slusarewicz, Noel, Wielgus and Nielsen2017; Cain et al. Reference Cain, Slusarewicz, Rutledge, McVey, Wielgus, Zynda, Wehling, Scare, Steuer and Nielsen2020) and small ruminant (Slusarewicz et al. Reference Slusarewicz, Slusarewicz and Nielsen2019) faecal samples. The purpose of this study was to validate PS as, to our knowledge, the first automated system for the detection and quantification of A. galli ova in chicken faeces. Such a system could be used to help expedite and simplify the use of FECs as a health surveillance tool in commercial chicken operations globally.
Materials and methods
Chickens and sample collection
Samples were collected in a commercial indoor aviary cage-free layer farm. The building was a Fienhage aviary with two levels, housing a total of 200,000 white leghorns. Both levels provided free access to a scratch area with a coated marine flooring and 1–3 inches of standing litter. Samples were collected from a flock verified to harbour only Ascaridia and to be Heterakis-free.
Faecal egg counts
Three sample preparation tools (Figure 2A) were used to prepare each sample. Six grams of faeces were placed in each tool, which was then filled with 54 mL of flotation solution (specific gravity 1.30; FecaMed, Vedco Inc., Saint Joseph, Missouri). The samples were thoroughly homogenised using the tool’s spring-actuated handle, and then all three were strained through the filter cap (Figure 2B) and pooled in a beaker. The resulting faecal slurry was mixed and swirled to ensure homogenisation prior to each sample being dispensed.

Figure 2. Parts comprising the Parasight System for Ascaridia galli quantitative counts. A) sample prep tool with filter top, B) homogeniser top, C) egg chamber, D) reagent dispensing unit where chemistry takes place, E) imaging unit, and F) a representative image of stained A. galli eggs identified by the algorithm and circled in magenta.
Samples were prepared for PS by filling each of ten 15 mL tubes with 8 mL of faecal slurry. The samples were then centrifuged in an MXU Centrifuge (LW Scientific, Lawrenceville, Georgia) for 2 min at 626 x g, and the resulting supernatant was poured into an egg chamber (Figure 2C) following the manufacturer’s instructions for small ruminant samples. After staining took place on the reagent dispending unit (Figure 2D), the egg chamber was then placed into the imaging unit (Figure 2E) and analysed (Figure 2F) using a custom application containing a machine learning algorithm trained using 5000+ images of stained A. galli oocysts.
For manual counts, the same faecal slurry was also used to fill both chambers of ten separate MM slides (FEC Source, Grand Ronde, Oregon), which were allowed to sit for 10 min before they were manually counted by a parasitologist using a Nikon Eclipse E200 microscope (Tokyo, Japan). The multiplication factor for MM counts using this method is 33.3.
Statistical analysis
Correlation between MM and PS using both raw and mean values was assessed with the coefficient of determination (R2) using the Pearson’s product−moment correlation. Raw values were paired by sorting the 10 individual counts for each method from lowest to highest value. A correlation was considered strong when R2 ≥ 0.75 and statistically significant when α ≤ 0.05. Precision was analysed by determining the coefficient of variation (CV) for each set of 10 replicate counts performed each of the 20 slurries generated. CVs were calculated as the standard deviation multiplied by 100 and divided by the mean of the replicate set. The mean CVs for MM and PS were then compared using student’s T-test, with results considered statistically significant when α ≤ 0.05. Raw egg magnitudes were compared with paired-samples sign tests. All statistical analysis was completed using R version 4.1.2. (R Core Team 2024).
Results
Overall, a total of 20 samples were analysed for use in this study. Ten repeated counts were performed for each sample and method for a total of 200 individual counts per method. Counts ranged from 0–1898 EPG based on MM.
On average, PS detected 15 times more eggs than did MM (Figure 3A), which was statistically significant (P < 0.0001). This factor could be used to normalise PS data to produce EPG results comparable to those of MM (Figure 3B). In addition, there was a strong, statistically significant correlation between individual (R2 = 0.7914, P < 0.0001) and mean (R2 = 0.7879, P < 0.0001) PS and MM counts (Figure 4) after raw PS results were refactored to produce MM-equivalent EPG counts.
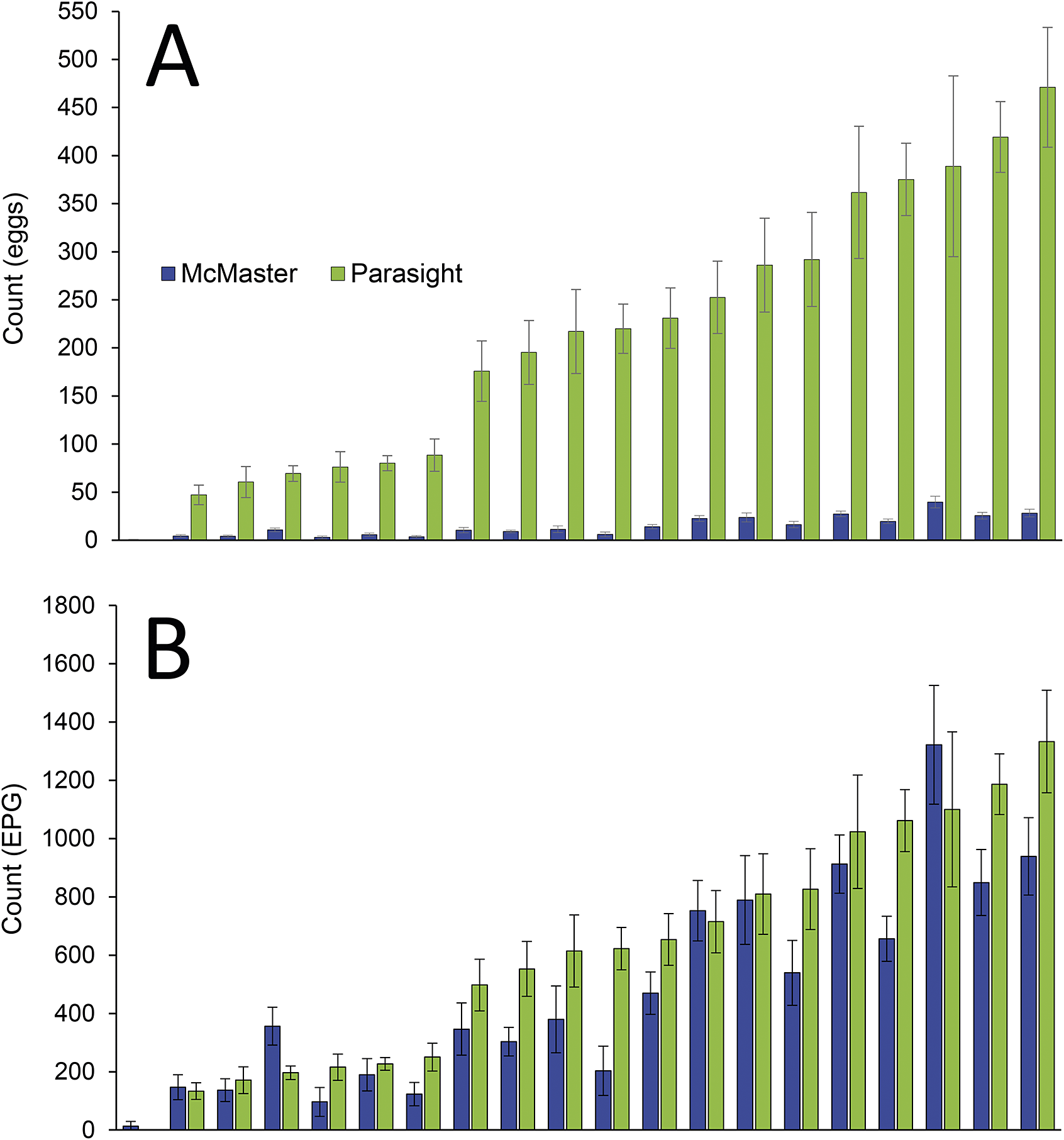
Figure 3. Comparison of A. galli faecal egg counts (FEC) for both the Parasight System and McMaster counts using A) counts converted to eggs per gram of faeces and B) raw number of eggs counted.

Figure 4. Scatterplots of A) all individual counts for the Parasight System and McMaster and B) means of ten repeated counts performed for each of the twenty individual samples.
Precision was determined from the CVs of each of the 20 sets of 10 replicate counts. PS had a mean CV of 22.0% (range 0–37.4%) whereas MM had a mean CV of 38.9% (range 15.3–165.8%), and this difference was statistically significant (P = 0.0391).
Discussion
This comparative study found that larger numbers of eggs were detected by PS compared with MM (Figure 3A). This was likely a reflection of the large volume of sample that can be examined by PS because the faecal slurry is poured through the mesh of the egg chamber, thereby trapping the ova on its surface prior to chemical processing and imaging. A total of 8 mL of slurry is examined using PS versus only 0.3 mL under the grid of the MM slide, i.e. nearly 27 times more faeces is analysed with PS.
Despite analysing 27 times more material than MM, however, PS counted only 15 times more ova, suggesting that egg recovery using the PS method is less efficient than with MM, as described previously (Britton et al. Reference Britton, Ripley and Slusarewicz2024). Three possible sources of PS egg loss are entrapment of ova in the faecal sediment during centrifugation, adhesion of ova to the centrifuge tube under high centripetal forces, and obscurement of ova under faecal debris on the egg chamber mesh. Nevertheless, the much larger amount of faeces analysed by PS still leads to the detection of over an order of magnitude more ova, which has potential repercussions on other assay parameters such as precision and sensitivity (see below).
The multiplication factor (MF) of an FEC method is a function of the concentration of the slurry used and the volume of that slurry examined. These values can be used to calculate the number of grams examined, and that value can be used to normalise the data by converting the raw count into the number of ova expected in one gram of sample, i.e. EPG. The MF for MM in this study was 33.3 (0.1 g/mL slurry, 0.3 mL counted = 0.03 g counted, and 1/0.03 = 33.3) whereas the MF of PS was 1.25 (0.1 g/mL slurry, 8 mL counted = 0.8 g counted, and 1/0.8 = 1.25).
Multiplying the mean raw MM count (14.3) by 33.3 produced a mean EPG of 476 whereas multiplying PS raw count (215.4) by 1.25 produced an EPG of 269, reflecting the greater degree of egg loss with PS. When one FEC method produces a higher count, it is relatively more accurate than the method it is being compared with (Nielsen Reference Nielsen2021); however, accuracies can be corrected using empirical data such as those presented here (Britton et al. Reference Britton, Ripley and Slusarewicz2024).
In this case, the PS raw results were normalised to MM EPGs using an MF of 2.83 to produce linear correlations with slopes close to unity (Figure 4). Such normalisation to a widely used standard proffers the advantage of there being no need for veterinary practitioners or chicken producers to alter their clinical protocols to accommodate a new method.
Demonstration of a linear correlation between new and established methods is an important proof-of-concept step for novel FEC techniques (Nielsen Reference Nielsen2021). Numerous studies have used linear correlations in this respect (Noel et al. Reference Noel, Scare, Bellaw and Nielsen2017; Slusarewicz et al. Reference Slusarewicz, Slusarewicz and Nielsen2019; Nagamori et al. Reference Nagamori, Hall Sedlak, DeRosa, Pullins, Cree, Loenser, Larson, Smith and Goldstein2020; Cringoli et al. Reference Cringoli, Amadesi, Maurelli, Celano, Piantadosi, Bosco, Ciuca, Cesarelli, Bifulco, Montresor and Rinaldi2021; Nagamori et al. Reference Nagamori, Sedlak, DeRosa, Pullins, Cree, Loenser, Larson, Smith, Penn and Goldstein2021;), and many of these methods are now commercially available. It is important to note, however, that a linear correlation does not take into account precision, nor does it provide information on diagnostic performance of the test. Instead, it shows that the novel method produces counts that are consistent with currently used tests.
All FEC methods are susceptible to variability due to the relatively low concentration of ova present coupled with the random Poisson processes involved when subsampling specimens and faecal slurries (Torgerson et al. Reference Torgerson, Paul and Lewis2012). Precision, or repeatability, is the measure of a test’s ability to consistently produce FECs from the same sample that are clustered around the mean value. More precise tests (with smaller CVs) are therefore more likely to produce an accurate result from any single test. This is important in clinical or agricultural practice, where there is seldom time to conduct multiple tests to help average out sampling errors.
Variability in FEC tests can be separated into two categories: biological and technical (Cain et al. Reference Cain, Slusarewicz, Rutledge, McVey, Wielgus, Zynda, Wehling, Scare, Steuer and Nielsen2020; Nielsen Reference Nielsen2021). Biological variability refers to that inherently found between subsamples or the solid specimen due to uneven egg distribution within the faeces and can account for the majority of variability between FECs (Cain et al. Reference Cain, Slusarewicz, Rutledge, McVey, Wielgus, Zynda, Wehling, Scare, Steuer and Nielsen2020) Technical variability, on the other hand, refers to the variability inherent in any given test methodology; it can be measured by removing the factor of biological variability by performing repeated counts on the same faecal slurry, thereby only testing the ability of a given method to provide precise results. While repeated counts on the same slurry do not account for different homogenisation methods, which can affect results (Went et al. Reference Went, Scare, Steuer and Nielsen2018; Nielsen Reference Nielsen2021), it is the best approximation of method precision in terms of parameters such as the actual counting of parasite eggs, egg loss during filtration and/or centrifugation steps, and the ability of eggs to float (Nielsen Reference Nielsen2021).
In this study, PS detected more eggs than MM and was also more precise in terms of technical variability. This is unsurprising since Poisson processes are known to generate lower CVs when counts are high (Torgerson et al. Reference Torgerson, Paul and Lewis2012). Further work will be required, however, to determine whether these higher counts also produce higher precision with respect to biological variability.
The quantitative parameters of accuracy and precision are the most important test performance parameters when conducting FECs (Nielsen Reference Nielsen2021) and thus were the focus of this study. Clinical sensitivity and specificity are the parameters that describe the ability of a test to produce true positive or true negative results. In terms of FECs, sensitivity only becomes relevant when counts are very low, and specificity must be evaluated using ova-negative samples. Overall, 75% of all samples examined in this study were over 200 EPG, and none were negative. Thus, a comprehensive examination of clinical sensitivity and specificity was not conducted.
Nevertheless, PS did generate 10 false-negative results in one set of 10 replicates from a sample whose mean MM count was less than 10 EPG whereas MM generated only seven. It is unclear why this would be the case, since PS was able to count many more eggs than MM in all the other samples. One possibility is that the three MM results from this sample were in fact false positives caused by cross-contamination of the McMaster slides due to inadequate washing between tests, but in any case, these presumptive PS false-negative results would not have affected any treatment decisions. Low egg counts can, however, be important when performing an FECRT in order to detect a decrease in anthelmintic efficacy, particularly in the light of new guidelines emphasising the number of eggs counted for FECRT in other agriculturally relevant animals (Kaplan et al. Reference Kaplan, Denwood, Nielsen, Thamsborg, Torgerson, Gilleard, Dobson, Vercruysse and Levecke2023). As a result, assessment of PS performance at lower FECs is worthy of further investigation.
Finally, this study examined samples exclusively infected with A. galli. The chitin-binding protein used to stain A. galli eggs in this study also stains the ova all other nematodes we have tested to date, indicating that chitin is indeed a common component of the eggshells across this clade. As a result, it is highly likely that it will also stain the ova of H. gallinarum. However, it is unclear whether the features of these of these morphologically similar ova are sufficiently different to allow for their discrimination by an algorithm trained on the two different egg types; this is a subject worthy of further investigation.
In summary, the automated, fluorescence-based FEC detection and quantification system, PS, was shown to be a useful method for performing A. galli FEC with chicken faeces. Overall, PS was as accurate and more precise than the traditionally used manual MM method. This method provides farmers and veterinarians with a low cost, high volume, rapid method for implementation and analysis of anthelmintic control programs without the need for technical expertise to reliably perform the analyses. Future studies will focus on lower counts as well as assessing the unit’s utility for FECRT in chickens as well as turkeys.
Financial support
This work was supported by Parasight System Inc.
Competing interest
P. Slusarewicz is an employee of, and owns stock in, Parasight System Inc. At the time of this work, J. Cain was an employee of Parasight System Inc. D. Wilson has no conflicts to declare.




