



No CrossRef data available.
Article contents
Swimming efficiently by wrapping
Published online by Cambridge University Press: 13 September 2024
Abstract
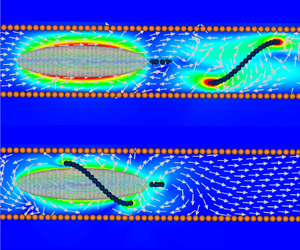
Single-flagellated bacteria are ubiquitous in nature. They exhibit various swimming modes using their flagella to explore complex surroundings such as soil and porous polymer networks. Some single-flagellated bacteria swim with two distinct modes, one with the flagellum extended away from its body and another with the flagellum wrapped around it. The wrapped mode has been observed when bacteria swim under tight confinements or in highly viscous polymeric melts. In this study we investigate the hydrodynamics of these two modes inside a circular pipe. We find that the wrapped mode is slower than the extended mode in bulk but more efficient under strong confinement due to a hydrodynamic increase of its flagellum translation–rotation coupling and an Archimedes’ screw-like configuration that helps to move the fluid along the pipe.
- Type
- JFM Papers
- Information
- Copyright
- © The Author(s), 2024. Published by Cambridge University Press
References
Balboa Usabiaga, F. & Delmotte, B. 2022 A numerical method for suspensions of articulated bodies in viscous flows. J. Comput. Phys. 464, 111365.CrossRefGoogle Scholar
Balboa Usabiaga, F., Kallemov, B., Delmotte, B., Bhalla, A.P.S., Griffith, B.E. & Donev, A. 2016 Hydrodynamics of suspensions of passive and active rigid particles: a rigid multiblob approach. Commun. Appl. Maths Comput. Sci. 11 (2), 217–296.CrossRefGoogle Scholar
Bansil, R., Constantino, M.A., Su-Arcaro, C., Liao, W., Shen, Z. & Fox, J.G. 2023 Motility of different gastric helicobacter spp. Microorganisms 11 (3), 634–648.CrossRefGoogle ScholarPubMed
Childress, S. 2012 A thermodynamic efficiency for Stokesian swimming. J. Fluid Mech. 705, 77–97.CrossRefGoogle Scholar
Constantino, M.A., Jabbarzadeh, M., Fu, H.C., Shen, Z., Fox, J.G., Haesebrouck, F., Linden, S.K. & Bansil, R. 2018 Bipolar lophotrichous helicobacter suis combine extended and wrapped flagella bundles to exhibit multiple modes of motility. Sci. Rep. 8 (1), 1–15.CrossRefGoogle ScholarPubMed
Das, D. & Lauga, E. 2018 Computing the motor torque of Escherichia coli. Soft Matt. 14, 5955–5967.CrossRefGoogle ScholarPubMed
Grognot, M. & Taute, K.M. 2021 More than propellers: how flagella shape bacterial motility behaviors. Curr. Opin. Microbiol. 61, 73–81.CrossRefGoogle ScholarPubMed
Higdon, J.J.L. 1979 The hydrodynamics of flagellar propulsion: helical waves. J. Fluid Mech. 94 (2), 331–351.CrossRefGoogle Scholar
Kinosita, Y., Kikuchi, Y., Mikami, N., Nakane, D. & Nishizaka, T. 2018 Unforeseen swimming and gliding mode of an insect gut symbiont, Burkholderia sp. RPE64, with wrapping of the flagella around its cell body. ISME J. 12 (3), 838–848.CrossRefGoogle ScholarPubMed
Kühn, M.J., Schmidt, F.K., Eckhardt, B. & Thormann, K.M. 2017 Bacteria exploit a polymorphic instability of the flagellar filament to escape from traps. Proc. Natl Acad. Sci. USA 114 (24), 6340–6345.CrossRefGoogle ScholarPubMed
Kühn, M.J., Schmidt, F.K., Farthing, N.E., Rossmann, F.M., Helm, B., Wilson, L.G., Eckhardt, B. & Thormann, K.M. 2018 Spatial arrangement of several flagellins within bacterial flagella improves motility in different environments. Nat. Commun. 9 (1), 5369.CrossRefGoogle ScholarPubMed
Lauga, E., DiLuzio, W.R., Whitesides, G.M. & Stone, H.A. 2006 Swimming in circles: motion of bacteria near solid boundaries. Biophys. J. 90 (2), 400–412.CrossRefGoogle ScholarPubMed
Lauga, E. & Eloy, C. 2013 Shape of optimal active flagella. J. Fluid Mech. 730, R1.CrossRefGoogle Scholar
Liron, N. & Shahar, R. 1978 Stokes flow due to a Stokeslet in a pipe. J. Fluid Mech. 86 (4), 727–744.CrossRefGoogle Scholar
Liu, B., Breuer, K.S. & Powers, T.R. 2014 Propulsion by a helical flagellum in a capillary tube. Phys. Fluids 26 (1), 011701.CrossRefGoogle Scholar
Murat, D., Hérisse, M., Espinosa, L., Bossa, A., Alberto, F. & Wu, L.-F. 2015 Opposite and coordinated rotation of amphitrichous flagella governs oriented swimming and reversals in a magnetotactic spirillum. J. Bacteriol. 197 (20), 3275–3282.CrossRefGoogle Scholar
Park, J., Kim, Y., Lee, W. & Lim, S. 2022 Modeling of lophotrichous bacteria reveals key factors for swimming reorientation. Sci. Rep. 12 (1), 6482.CrossRefGoogle ScholarPubMed
Pimponi, D., Chinappi, M., Gualtieri, P. & Casciola, C.M. 2016 Hydrodynamics of flagellated microswimmers near free-slip interfaces. J. Fluid Mech. 789, 514–533.CrossRefGoogle Scholar
Shum, H., Gaffney, E.A. & Smith, D.J. 2010 Modelling bacterial behaviour close to a no-slip plane boundary: the influence of bacterial geometry. Proc. R. Soc. Lond. A 466 (2118), 1725–1748.Google Scholar
Thormann, K.M., Beta, C. & Kühn, M.J. 2022 Wrapped up: the motility of polarly flagellated bacteria. Annu. Rev. Microbiol. 76 (1), 349–367.CrossRefGoogle ScholarPubMed
Tian, M., Wu, Z., Zhang, R. & Yuan, J. 2022 A new mode of swimming in singly flagellated pseudomonas aeruginosa. Proc. Natl Acad. Sci. USA 119 (14), e2120508119.CrossRefGoogle ScholarPubMed
Vizsnyiczai, G., Frangipane, G., Bianchi, S., Saglimbeni, F., Dell'Arciprete, D. & Di Leonardo, R. 2020 A transition to stable one-dimensional swimming enhances E. coli motility through narrow channels. Nat. Commun. 11 (1), 2340.CrossRefGoogle ScholarPubMed
Wajnryb, E., Mizerski, K.A., Zuk, P.J. & Szymczak, P. 2013 Generalization of the Rotne–Prager–Yamakawa mobility and shear disturbance tensors. J. Fluid Mech. 731, R3.CrossRefGoogle Scholar
Xing, J., Bai, F., Berry, R. & Oster, G. 2006 Torque–speed relationship of the bacterial flagellar motor. Proc. Natl Acad. Sci. USA 103 (5), 1260–1265.CrossRefGoogle ScholarPubMed
Yan, W. & Shelley, M. 2018 Universal image system for non-periodic and periodic Stokes flows above a no-slip wall. J. Comput. Phys. 375, 263–270.CrossRefGoogle Scholar

Gidituri et al. supplementary movie 1
In plane velocity field around a bacterium swimming in the extended and the wrapped mode. In these simulations only the flagellum orientation is updated to ease the comparison between both swimming modes. In the wrapped mode the flagellum works as an Archimedes' screw helping to push fluid to the back of the bacterium.

File
1.8 MB
Gidituri et al. supplementary movie 2
Cross-section view of a bacterium swimming in the extended and the wrapped mode inside a wide pipe. The bacterium in the extended mode is hydrodynamically attracted to the pipe's walls and it reaches a steady state distance to them. The bacterium in the wrapped mode is repelled by the walls and it does not reach a steady state distance.
File
6.2 MB
Gidituri et al. supplementary movie 3
A bacterium in both swimming modes swimming inside a narrow pipe. The bacterium in the extended mode is constrained to swim parallel to the pipe axis while the bacterium in the wrapped mode shows a precession motion around its main axis and a tilt towards the pipe's walls.
File
6.3 MB
Gidituri et al. supplementary movie 4
A bacterium with a long flagellum swimming inside a narrow pipe. If the flagellum is long enough it constraints the bacterium in both swimming modes to swim parallel to the pipe axis.
File
6.3 MB