


INTRODUCTION
Historically the Mediterranean Basin is the most important area for production of durum wheat (Triticum turgidum L. var. durum), being the most significant durum import market, and the largest consumer of products derived from this cereal grain (Royo et al. Reference Royo, Elias, Manthey and Carena2009). Many traditional Mediterranean foods are manufactured from durum wheat. Pasta is the most common durum end product in southern Europe and North Africa as well as in non-Mediterranean regions such as North America and the former Soviet Union. Durum wheat is extensively consumed as couscous in North Africa and flat bread and bulgur are part of the staple diet in Eastern-Mediterranean countries. Durum wheat is usually cultivated under rainfed conditions in the Mediterranean Basin, which often imposes a number of environmental stresses on the crop. Terminal drought stress, a combination of water scarcity and warm temperatures during the grain-filling period, usually results in yield reductions, but in most cases results in high grain quality.
Archaeological evidence shows that the earliest domesticated wheats came from the Near East, in the region known as the Fertile Crescent. Dated to c. 10 000 years before the present time, they spread to the west of the Mediterranean Basin, reaching the Iberian Peninsula c. 7000 years BP (Feldman Reference Feldman, Bonjean and Angus2001). This migration process and both natural and human selections resulted in the establishment of local landraces along the Mediterranean Basin that were well adapted to diverse but specific agro-ecological zones. These unimproved landraces, which contain the largest genetic diversity within the species, practically disappeared from farmers’ fields as a consequence of the introduction of the more productive and homogeneous semi-dwarf varieties released since the Green Revolution. The extreme homogeneity in cultivar structures typical of modern agriculture caused a dramatic loss of natural variation from the middle of the 20th century in southern Europe and during the 1970s–80s in Northern Africa, which resulted in genetic erosion and possibly an increase in genetic vulnerability of wheat crops. Landraces, mostly conserved in germplasm repositories, can be considered as likely sources of putatively lost variability and may provide new favourable genes/alleles, which could be introgressed into modern cultivars.
Some studies have shown the existence of variability for quality traits in durum wheat landraces (Moragues et al. Reference Moragues, Zarco-Hernández, Moralejo and Royo2006; Aguiriano et al. Reference Aguiriano, Ruiz, Fité and Carrillo2008), as well as in related tetraploid species (Sissons & Batey Reference Sissons and Batey2003). The ability to tap into this diversity depends on the identification of accessions containing genes and alleles demonstrated to be useful in breeding programmes for quality improvement. Gluten strength is one of the main factors influencing durum wheat quality. It is commonly determined by the sodium dodecyl sulphate (SDS)-sedimentation test and depends on the composition of the storage proteins. They are comprised of gliadins (monomeric proteins) and glutenins (polymeric proteins), the major components of gluten with the latter reported to be the most influential on gluten strength. Based on their mobility in sodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS–PAGE), single polypeptides from the glutenin fraction are separated after di-sulphide bond reduction, into high molecular weight-glutenin subunits (HMW-GS) and low molecular weight-glutenin subunits (LMW-GS). The HMW-GS are encoded by the complex at the Glu-1 loci (Glu-A1 and Glu-B1), located on the long arm of group 1 homologous chromosomes (Shewry et al. Reference Shewry, Halford and Tatham1992), whereas the LMW-GS are encoded by genes at Glu-A3, Glu-B3 and Glu-B2 loci located on the short arm of group 1 homologous chromosomes (Vázquez et al. Reference Vázquez, Ruiz, Nieto-Taladriz and Albuquerque1996).
The allelic polymorphism controlling the HMW-GS and the LMW-GS has been used in diversity studies and cultivar identification. However, the difficulty in scoring alleles of the LMW-GS has limited their use in research (Lerner et al. Reference Lerner, Kolman and Rogers2009) and hindered the selection for specific LMW-GS alleles in wheat breeding programmes. Recent improvements in uni-dimensional SDS–PAGE separation protocols implemented at the International Maize and Wheat Improvement Center (CIMMYT) have allowed better discrimination between LMW-GS alleles and have opened up the possibility of using them effectively in diversity studies (Peña et al. Reference Peña, Gonzalez-Santoyo, Cervantes, Masci, Lafiandra and D'Ovidio2004).
The present study examined the SDS–PAGE glutenin protein banding patterns in a set of 155 Mediterranean durum wheat landraces from 21 countries and a set of 18 representative modern varieties, with the main purpose of determining the diversity of banding pat-terns related to known allelic variability for HMW-GS and LMW-GS. The relationship between glutenin composition and gluten strength and the possible existence of a geographic structure in the collection, based on both glutenin composition and gluten strength, were also investigated.
MATERIALS AND METHODS
Plant material and experimental set up
The study was conducted on a collection of 155 durum wheat landraces and old varieties from 21 Mediterranean countries including the major durum producers/users (Table 1) and 18 representative modern cultivars. Landraces were selected from a larger collection of 231 accessions, on the basis of their genetic variability determined by 33 simple sequence repeat (SSR) markers, in order to represent the genetic diversity of ancient local durum populations from the Mediterranean Basin (Nazco et al. Reference Nazco, Villegas, Ammar, Peña, Moragues and Royo2012). The modern set included the following varieties: Amilcar, Ancalei, Arment, Astigi, Boabdil, Bolo, Claudio, Gallareta, Hispasano, Jupare, Meridiano, Ocotillo, Senadur, Simeto, Sula, Svevo, Vitron and Vitronero. Several of them, particularly those from Spain, are derived from CIMMYT and have been released, under different names, as major cultivars also in North Africa and elsewhere around the Mediterranean Basin. The landraces received from the Germplasm Banks in 2005 were bulk purified selecting the dominant type (usually with a frequency above 80% of the bulk) and increased in 2006. Seed used for the field experiments from 2007 to 2009 came from purified bulks grown during each previous year. Grain samples were collected from field trials established under rainfed conditions during 3 years (2007–09) at Gimenells (41°40′N, 0°20′E, 200 m a.s.l.) in the Lleida province (north-eastern Spain). Field experiments consisted of non-replicated entries sown in plots of 6 m2 comprising 5 m rows, spaced 0·15 m apart, and arranged in a modified augmented design with three replicated controls (cultivars Claudio, Simeto and Vitron). Sowing density was adjusted to 250 viable seeds/m2 and plots were kept free of weeds and diseases according to standard cultural practices. A sample of grain, mechanically harvested at full maturity, was drawn randomly from each plot, cleaned and used for gluten-strength determination. Data of other quality attributes determined on grain samples from the same experiments used in this investigation may be found in Nazco et al. (Reference Nazco, Villegas, Ammar, Peña, Moragues and Royo2012).
Table 1. Country of origin of the 155 landraces included in the study and number of locus-specific banding patterns detected by SDS–PAGE at the HMW- and LMW-glutenin subunits loci

High molecular weight-glutenin subunits and LMW-GS allelic composition
Electrophoresis was run on a bulk of ten seeds from a purified head-row of the dominant type. The procedure was as follows: a spike of the dominant type was selected from the population and its grains were sown in a single row that was harvested at ripening and ten seeds were bulked for electrophoresis.
Electrophoretic analysis (1D SDS–PAGE) was conducted according to the protocols implemented at CIMMYT by Peña et al. (Reference Peña, Gonzalez-Santoyo, Cervantes, Masci, Lafiandra and D'Ovidio2004), following the nomenclature of Nieto-Taladriz et al. (Reference Nieto-Taladriz, Ruiz, Martínez, Vázquez and Carrillo1997) and Martinez et al. (Reference Martinez, Ruiz and Carrillo2004), to identify high- and low molecular weight glutenin subunit compositions at five loci (Glu-A1, Glu-B1, Glu-A3, Glu-B3 and Glu-B2). The banding patterns/alleles with a frequency below 0·05 were classified as rare.
Gluten-strength determination
Gluten-strength was determined for each experimental field plot on a sample of 1 g of whole wheat flour using the SDS-sedimentation test of Axford et al. (Reference Axford, Mcdermott and Redman1978), as modified by Peña et al. (Reference Peña, Amaya, Rajaram and Mujeeb-Kazi1990).
Statistical analysis
The SDS-sedimentation data were fitted to a linear mixed model with the control cultivars as fixed effects, and row number, column number and genotype as random effects (Littell et al. Reference Littell, Milliken, Stroup and Wolfinger1996). Restricted maximum likelihood (REML) was used to estimate the variance components and to produce the best linear unbiased predictors (BLUPs) for data from each cultivar/year combination. The calculations were done through the MIXED procedure of the SAS–STAT statistical package, which was used for all the analyses. A standard analysis of variance (ANOVA) was conducted with the BLUPs of gluten-strength data using the genotype × year interaction as an error term. Genotypes were classified according to the mean value of SDS-sedimentation across experiments as outstanding (SDS ⩾ 11), very high (10 < SDS < 11), high (9 ⩽ SDS ⩽ 10), medium (7 ⩽ SDS < 9) and low (SDS < 7) gluten-strength groups and the genotype effect was partitioned in the ANOVA according to this classification. Means were compared by the Student–Newman–Keuls (SNK) test at P = 0·05. The FREQ procedure of the SAS–STAT statistical package (SAS 2011) was used to conduct a 2 × 5 Fisher's exact test on each significant allele/banding pattern, to determine whether there were any significant differences between the gluten-strength groups with regard to banding pattern frequency. Genetic diversity was calculated with the D index, according to the following:
 $$D_j {\rm} = {\rm} 1 - \Sigma {\rm} p_{ij}^2 $$
$$D_j {\rm} = {\rm} 1 - \Sigma {\rm} p_{ij}^2 $$where p is the frequency of the i th allele at locus j th. The allelic effect on gluten strength was computed as the difference between the mean SDS-sedimentation values across years of genotypes carrying or not-carrying a given allele/banding pattern. The allelic frequency of the allele Glu-A1c (null) for each country was used to conduct hierarchical cluster analyses by the Ward method of the JMP V.8 software (SAS 2009).
Results
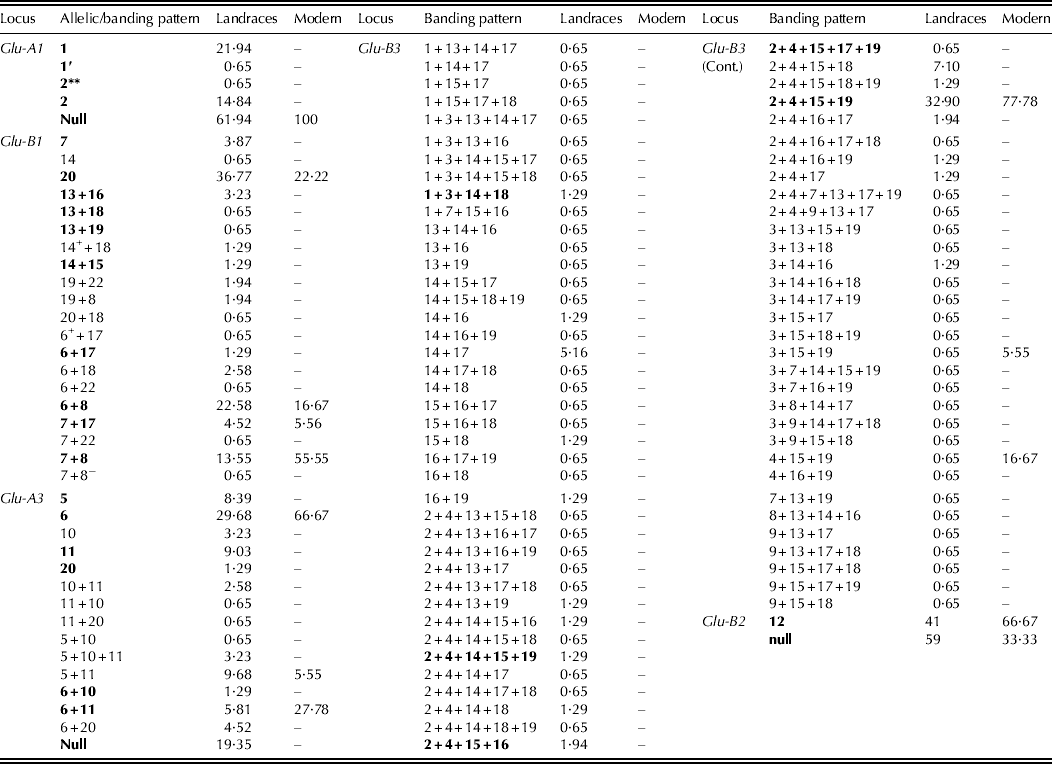
One hundred and fourteen banding patterns, potentially Glu-1/Glu-3 allele-specific, were identified by SDS–PAGE (bottom part of Table 1), 97 of which appeared only in the landraces and with frequencies below 0·05, thus being considered herein as rare patterns/alleles. All the patterns detected in modern cultivars, which are considered allele-specific according to the Glu-1/Glu-2/Glu-3 nomenclature used, were also present in the landraces. For the two loci encoding for HMW-GS, 25 alleles/banding patterns were identified in landraces and only five of them were detected in the modern varieties. For the three loci encoding the LMW-GS the difference in variability was even higher between the two groups, with landraces exhibiting some 89 potentially allele-specific banding patterns and only eight of them preserved in the modern cultivars. Glu-B3, encoding for LMW-GS, was the locus showing the largest number of patterns in landraces, while Glu-B1 (HMW-GS) was the most variable locus in the modern cultivars (bottom part of Table 1).
The allelic/banding pattern frequencies at each presumed locus are shown in Table 2. Only 25·4% of the banding patterns detected have been previously described. The most frequent allele in the collection was the null allele at Glu-A1, and was monomorphic in modern cultivars. A banding pattern tentatively designated as 2** (Branlard et al. Reference Branlard, Autran and Monneveux1989) was detected in the area corresponding to the size of the subunits expected at this locus. At the Glu-B1 locus, the second one coding for HMW-GS, modern cultivars exhibited the three most frequent banding patterns known to be present in germplasm groups worldwide, namely, 7 + 8, 6 + 8 and 20 (Branlard et al. Reference Branlard, Autran and Monneveux1989; Aguiriano et al. Reference Aguiriano, Ruiz, Fité and Carrillo2008), while as many as 20 banding patterns could be distinguished in the landrace group in the mobility area corresponding to these HMW-GS. Subunit 20 was the most common in the landraces, whereas 7 + 8 was the most common in modern varieties (Table 2). At Glu-A3, of the 15 patterns identified overall, the most frequent was allele Glu-A3a (band 6). The second most prevalent allele in the modern varieties was Glu-A3d (bands 6 + 11), and the null allele in the landraces, the latter being absent in modern cultivars. The two alleles reported and described elsewhere (Nieto-Taladriz et al. Reference Nieto-Taladriz, Ruiz, Martínez, Vázquez and Carrillo1997; Carrillo et al. Reference Carrillo, Martínez, Moita Brites, Nieto-Taladriz, Vázquez, Royo, Nachit, Di Fonzo and Araus2000) at Glu-B2 (Glu-B2b or null and Glu-B2a or band 12) were observed at higher frequencies in both sets of germplasm, with the null allele being the most frequent in the landraces, whereas Glu-B2a was found at twice the frequency of the alternative allele in modern varieties. At the most variable locus overall, Glu-B3, 72 banding pattern/putative alleles were found in landraces but only three of them were present in modern varieties and 55 of them were recorded only in a single landrace (Table 2). The most frequent banding pattern at this locus was 2 + 4+15 + 19 (Glu-B3a). Other patterns with relatively high frequency in the landraces were 2 + 4+15 + 18 and 14 + 17, and 4 + 15 + 19 in modern varieties (Table 2). The allele 3 + 15 + 19 was found only in the modern French cultivar Arment.
Table 2. Allelic/banding pattern frequencies (%) at the HMW- and LMW-glutenin subunits loci for 155 landraces and 18 representative modern varieties. Known banding patterns are indicated in bold type

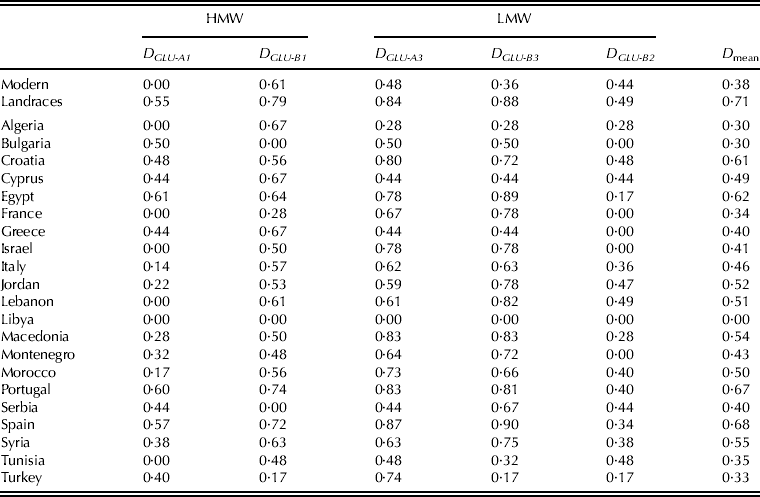
The overall genetic diversity index (D) for landraces was 0·71, substantially higher than the same index for modern varieties (0·38) (Table 3). It was highest in landraces from Spain (0·68), Portugal (0·67), Egypt (0·62) and Croatia (0·61), and lowest in accessions from Algeria and Bulgaria (0·30), Turkey (0·33), France (0·34) and Tunisia (0·35) (Table 3). For the HMW-G subunits locus Glu-A1, the highest diversity indices were observed in landraces from Egypt, Portugal and Spain, while landraces from Portugal, Spain, Algeria, Cyprus and Greece had the highest diversity indices for Glu-B1. The largest diversity was found in landraces from Spain, Portugal, Macedonia and Croatia at Glu-A3, from Spain, Egypt, Macedonia and Lebanon at Glu-B3 and from Lebanon, Croatia, Tunisia and Jordan for Glu-B2 (Table 3).
Table 3. Genetic diversity indices calculated for five glutenin loci for 155 landraces and 18 representative modern varieties and for the landraces per country of origin
The ANOVA for gluten strength showed that both year and genotype effect were statistically significant (P < 0·001), accounting for 9 and 73%, respectively, of the total variation (Table 4) (see also Nazco et al. Reference Nazco, Villegas, Ammar, Peña, Moragues and Royo2012). The partitioning of the genotype effect into its individual components revealed that differences between groups of accessions, based on their SDS-sedimentation values, accounted for 91·5% of the genotypic variability, which corresponded to 67% of the total variation, while differences within each sedimentation group were not statistically significant (Table 4).
Table 4. Analysis of variance for gluten strength of 173 entries (155 landraces + 18 modern varieties). Genotype effect was partitioned according to five levels of gluten strength (SDS-values): outstanding (SDS ⩾ 11), very high (10 < SDS < 11), high (9 ⩽ SDS ⩽ 10), medium (7 ⩽ SDS < 9) and low (SDS < 7)

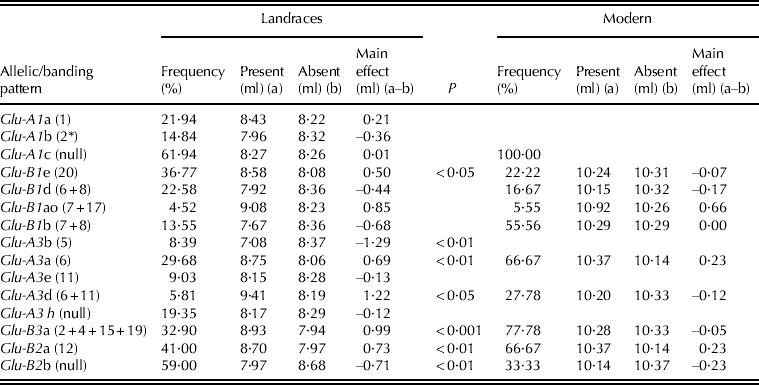
The effect on gluten strength of each allele/banding pattern was computed separately for landraces and modern varieties, as the difference between the mean SDS-sedimentation test value of the accessions carrying and not carrying the allele/pattern (Table 5). Allelic/banding pattern effect was not significant among the modern cultivars group, with all members showing high to outstanding SDS-values. On the other hand, significant main effects of some alleles/banding patterns were detected within the landraces. At the HMW-GS loci, the only allele with a significant effect was Glu-B1e (band 20). Four alleles/banding patterns at LMW-GS loci increased significantly the gluten strength of landraces (Table 5). Only three among the rare banding patterns found in the landraces had a significant effect on gluten strength (data not shown). One of them was putative allele 7 at Glu-B1, present in six landraces (three Montenegrin, one Croatian and two Spanish: 3·87% of the total landraces), which reduced the SDS-sedimentation value by 1·75 ml (P < 0·01) on average. The other two were banding patterns 2 + 4+14 + 15 + 18 (present in the Egyptian landrace PI-366109) and 2 + 4+15 + 18 + 19 (present in two landraces, Trigo Glutinoso from France and Lobeiro de Grao Escuro from Portugal) at Glu-B3, which increased SDS-values by 3·44 ml (P < 0·05) and 2·96 ml (P < 0·01), respectively.
Table 5. Effect of known allelic/banding patterns and their effect on SDS-sedimentation volume (ml) in the collection of 155 landraces and 18 modern cultivars
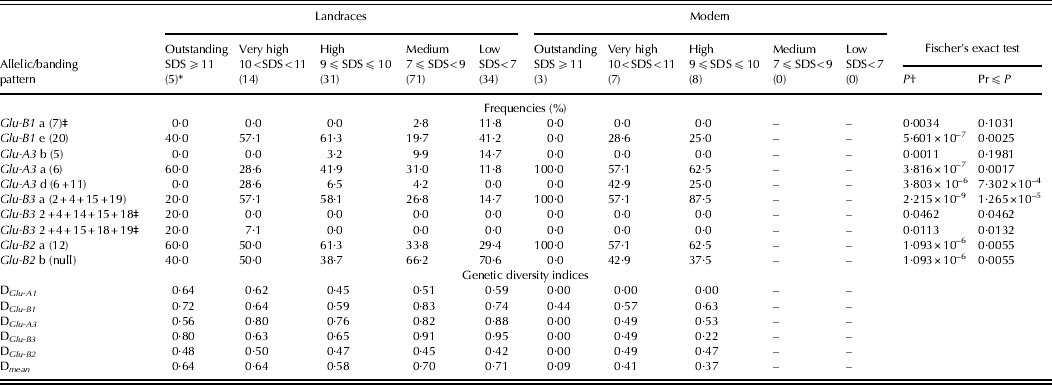
The frequency of alleles/banding patterns significantly affecting gluten strength was calculated for each of the gluten-strength groups considered in the ANOVA. The rare allele Glu-B1a (band 7) was not present in any of the 50 landraces with stronger gluten (SDS ⩾ 9 ml), but it was detected in 2·8% of the 71 landraces with medium gluten strength, and in 11·8% of the 34 landraces with low gluten strength (Table 6). For the landraces, allele Glu-B1e (band 20) was present with a high frequency in the five gluten-strength groups, particularly in c. 60% of the genotypes with high and very high gluten strength. It was also detected in modern varieties, but with lower frequencies and not in the outstanding gluten-strength group (Table 6). Band 5 (Glu-A3b) at locus Glu-A3 was absent in landraces with SDS-values >10, but its frequency increased in landraces as gluten strength decreased. In contrast, bands 6 (Glu-A3a) and 6 + 11 (Glu-A3d) at Glu-A3 and 2 + 4+15 + 19 (Glu-B3a) at Glu-B3 were detected in higher frequencies in accessions with SDS-values ⩾9, both in landraces and modern varieties. The rare patterns 2 + 4+14+ 15 + 18 and 2 + 4+15 + 18 + 19 were only present in the landraces with the highest SDS-values (Table 6). Alleles at Glu-B2 were uniformly distributed among all the gluten-strength groups for the landraces, but they were only present in modern varieties with SDS-values ⩾9. Band 12 (Glu-B2a) was present in more than 50% of genotypes with SDS-values ⩾9, while the alternative null (Glu-B2b) allele was highly frequent in landraces with medium and low SDS-values. The results of the Fisher's exact test revealed that, except for Glu-B1a (band 7) and Glu-A3b (band 5), differences between gluten-strength groups were associated with banding pattern frequencies (Table 6). When this test was conducted for landraces and modern varieties separately, no significant association appeared for modern varieties, while for the landraces the significant relationships coincided with the ones shown in Table 6 for the whole set of cultivars.
Table 6. Frequency of known and rare alleles/banding patterns with statistical significant effect on gluten-strength groups formed according to SDS-sedimentation values (ml), differences between gluten-strength groups with regard to banding pattern frequency according to the Fisher's exact test and genetic diversity indices for the same groups
* Number of genotypes in the strength group.
† Probability according to a hypergeometric distribution.
‡ Allelic/banding patterns with frequencies <0·05.
Only eight genotypes in the whole collection, five landraces and three modern varieties, had SDS-values ⩾11. None of them carried the alleles/banding patterns associated with negative main effects such as band 5 (GluA-3b) at Glu-A3 or the rare allele 7 at Glu-B1. However, alleles/patterns found to significantly increase gluten strength were detected in all of them. This was the case for band 6 (Glu-A3a) at Glu-A3 and band 12 (Glu-B2a) at Glu-B2, which were present in three of the five landraces and in the three modern varieties with the highest SDS-values, and the pattern 2 + 4+15 + 19 (Glu-B3a) at Glu-B3, which was also carried by the three modern varieties and one of the landraces with outstanding gluten strength.
The overall genetic diversity was much higher in landraces than in modern varieties, as noted previously. This tendency was also observed when considering diversity within the different gluten-strength groups (bottom part of Table 6). Modern varieties with SDS-sedimentation values ⩾11 were essentially monomorphic at Glu-A3, Glu-B3 and Glu-B2, all exhibiting the Glu-B3 pattern commonly known as LMW-2 type (Pogna et al. Reference Pogna, Lafiandra, Feillet and Autran1988), which is associated with strong gluten type and high pasta-cooking quality (Pogna et al. Reference Pogna, Lafiandra, Feillet and Autran1988; Peña & Pfeiffer Reference Peña, Pfeiffer, Royo, Nachit, Di Fonzo, Araus, Pfeiffer and Slafer2005). Modern varieties with high and very high gluten strength had intermediate-to-low diversity index values for all loci. The diversity indices for the five SDS-sedimentation groups of the landraces ranged from 0·58 to 0·71.
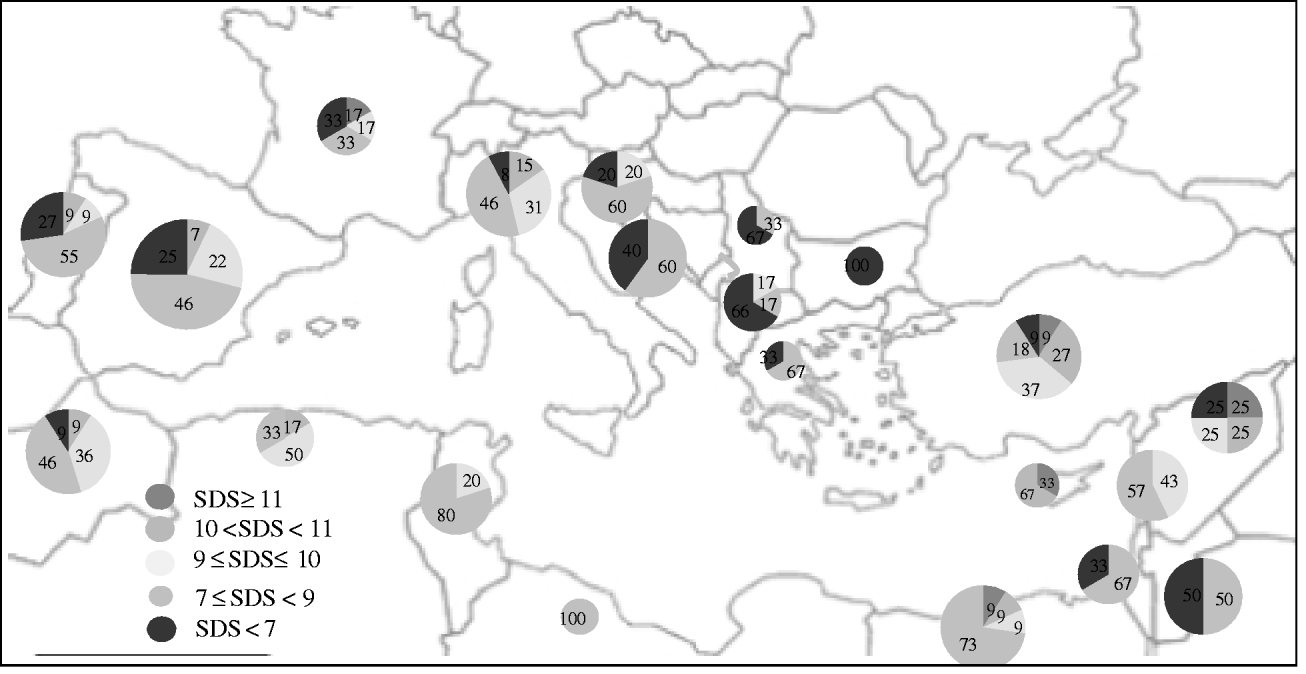
The geographic distribution of the five gluten-strength groups along the Mediterranean Basin is shown in Fig. 1. The highest percentages of accessions with strong to very strong gluten were found in Cyprus, Syria and Turkey, whereas 100% of the accessions from Jordan, Israel, Bulgaria, Serbia, Montenegro and Greece had medium to low gluten strength. All accessions from Algeria, Tunisia and Libya had SDS-values that were in the intermediate classes, with none belonging to either outstanding or low SDS-values groups. All the western Mediterranean countries had a high percentage of accessions with medium gluten-strength and, with the exception of France, none of the western Mediterranean accessions had SDS-values ⩾11.
Fig. 1. Geographic distribution of gluten-strength groups in landraces from the Mediterranean Basin. The colours inside the circles indicate the percentage of entries with outstanding (SDS ⩾ 11), very high (10 < SDS < 11), high (9 ⩽ SDS ⩽ 10), medium (7 ⩽ SDS < 9) and low (SDS < 7) SDS-values. The size of the circles is proportional to the number of entries from each country according to Table 1 (colour version is available online).
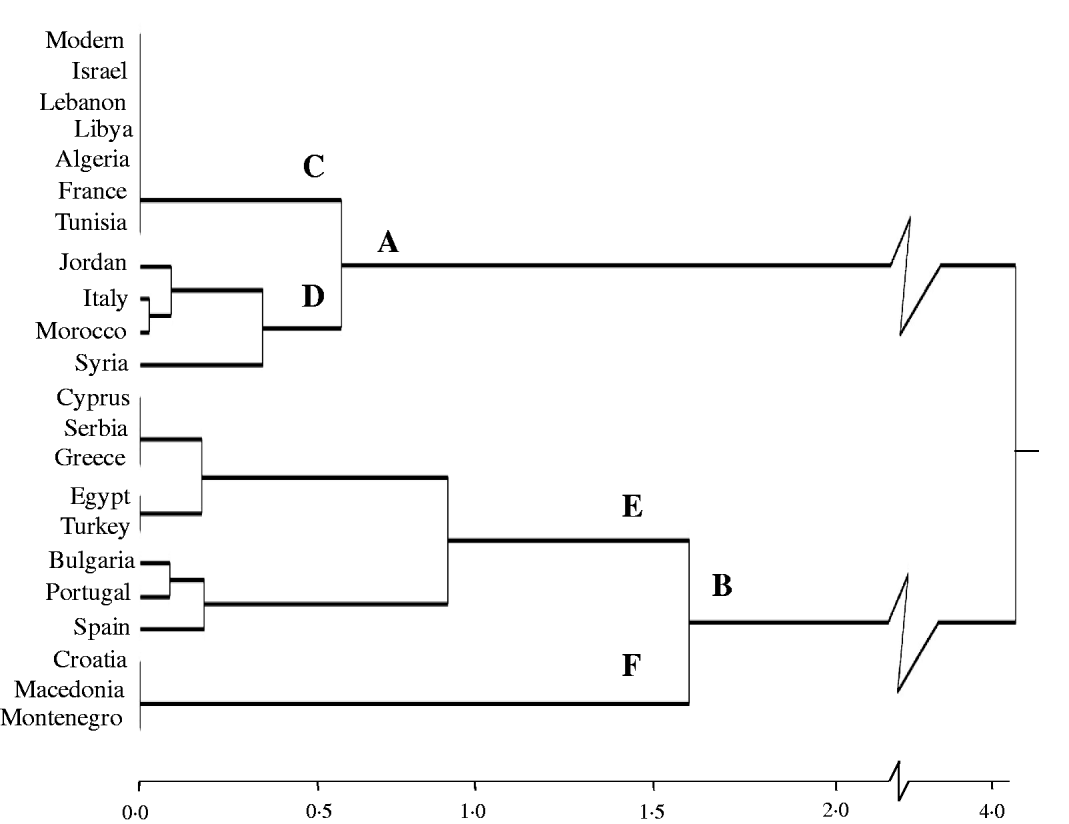
The clustering of countries based on the frequencies of all known alleles/banding patterns did not result in a clear geographical pattern except for the null allele (Glu-A1c) at Glu-A1, the most frequent in the present collection. Figure 2 shows that the allelic frequencies for the null allele clusters countries with high (75–100%, branch A) and low (0–60%, branch B) frequencies of this allele. Branch C clustered modern varieties and countries in which all the accessions were monomorphic for this locus, while branch D grouped countries in which this allele was present in very high frequency (75–92%). In the lower part of Fig. 2, branch E grouped countries with low to medium frequencies (27–60%), while Croatia, Macedonia and Montenegro, with accessions lacking this allele, were clustered in branch F.
Fig. 2. Cluster analysis based on Glu-A1c (null allele) frequency in modern varieties and by country for landraces.
DISCUSSION
Although the number of modern cultivars included for comparison in the present study was lower than that of landraces, it can be considered as reliably representative of the current modern Mediterranean germplasm in terms of its variability for glutenin loci, in particular for Glu-A1, Glu-B1 and Glu-B3. Because the genotypes are not only the major cultivars in the northern-Mediterranean countries where they were released, but also several are extensively grown under different names in North Africa and elsewhere, alleles or banding patterns that would be present in the overall modern Mediterranean germplasm are highly likely to be represented within the group of 18 cultivars included in this study. One exception is the LMW-1 pattern associated with low gluten strength, which was eliminated from most modern European varieties, but remains in some widely grown North-African cultivars. All others alleles present in the set of 18 modern cultivars were the overwhelmingly predominant ones identified in large collections of durum wheat by Branlard et al. (Reference Branlard, Autran and Monneveux1989), Sissons et al. (Reference Sissons, Ames, Hare and Clarke2005) and Igrejas et al. (Reference Igrejas, Guedes-Pinto, Carnide and Branlard1999).
In this context, the present collection of 155 landraces clearly showed much more variability in glutenin composition than modern varieties with only 13 of the 114 alleles/banding patterns identified in the landraces being also present in modern varieties. This indicates an overall loss of 88·6% in allelic variability going from landraces to modern cultivars. Eighty-five per cent of the alleles/banding patterns identified in the landraces had a frequency lower than 5%, therefore considered as rare forms, and 84·5% of them had not been described previously. This clearly confirms that local/native durum germplasm resources from the region indeed represent a rich reservoir of untapped diversity for glutenin composition and could be used towards widening the variability within modern germplasm, should some alleles or banding patterns prove to be useful for enhancing/diversifying gluten characteristics.
The largest diversity for HMW-GS was found at the Glu-B1 locus, both in landraces and modern varieties. However, at the Glu-A1 locus, modern cultivars were monomorphic, all exhibiting the null allele, whereas four alternative, non-null alleles were expressed in close to 40% of the landraces. The null allele has been found in very high frequencies in other durum collections (Branlard et al. Reference Branlard, Autran and Monneveux1989), and practically fixed in modern germplasm worldwide. The same order in allelic frequencies at Glu-A1, namely null >1 > 2*, has been reported previously in durum wheat (Kaan et al. Reference Kaan, Branlard, Chihab, Borries and Monneveux1993; Moragues et al. Reference Moragues, Zarco-Hernández, Moralejo and Royo2006; Aguiriano et al. Reference Aguiriano, Ruiz, Fité and Carrillo2008).
The genetic variability found at Glu-B1 in the present study was much greater than that reported by previous studies conducted with Mediterranean landraces (Moragues et al. Reference Moragues, Zarco-Hernández, Moralejo and Royo2006) or with durum world collections (Kaan et al. Reference Kaan, Branlard, Chihab, Borries and Monneveux1993). The four banding patterns identified at the Glu-B1 locus in modern varieties represented only 77·4% of the allelic frequency in the landraces group, in which 16 additional rare banding patterns were detected. However, the most frequent banding pattern in the landraces (HMWGS-20) did not coincide with that prevalent in modern varieties (HMWGS-7 + 8). Previous studies have reported a wide presence of band 20 in landraces from North Africa and South West Asia (Moragues et al. Reference Moragues, Zarco-Hernández, Moralejo and Royo2006). The high frequency of the 7 + 8 banding pattern in modern varieties may have resulted from selection in breeding programmes, since it has been widely associated with strong gluten and therefore good pasta-making quality (Du Cros Reference Du Cros1987; Sissons et al. Reference Sissons, Ames, Hare and Clarke2005), although its positive effect could not be evidenced in the present set of genotypes. The rare band 7 at Glu-B1 locus had been previously detected at very low frequencies in landraces from the Iberian Peninsula and South West Asia (Moragues et al. Reference Moragues, Zarco-Hernández, Moralejo and Royo2006).
The variability detected for LMW-GS was even wider than the one characterizing HMW-GS as found in other studies (Du Cros Reference Du Cros1987; Brites & Carrillo Reference Brites and Carrillo2001; Sissons et al. Reference Sissons, Ames, Hare and Clarke2005). Additionally, the number of banding patterns identified at LMW-GS loci in the present collection was much higher than that reported in previous studies, confirming the wide genetic diversity of the germplasm used in the present study, especially for the Glu-B3 locus. Nine of the 15 banding patterns detected at Glu-A3 locus were rare. The most frequent banding pattern at this locus, either in landraces and modern varieties, was Glu-A3a (band 6), in agreement with the results obtained by Nieto-Taladriz et al. (Reference Nieto-Taladriz, Ruiz, Martínez, Vázquez and Carrillo1997), Carrillo et al. (Reference Carrillo, Martínez, Moita Brites, Nieto-Taladriz, Vázquez, Royo, Nachit, Di Fonzo and Araus2000) and Moragues et al. (Reference Moragues, Zarco-Hernández, Moralejo and Royo2006). The 2 + 4+15 + 19 banding pattern (Glu-B3a allele) at Glu-B3 was the most prevalent in landraces as well as in modern varieties, in accordance with the results obtained by Nieto-Taladriz et al. (Reference Nieto-Taladriz, Ruiz, Martínez, Vázquez and Carrillo1997), Carrillo et al. (Reference Carrillo, Martínez, Moita Brites, Nieto-Taladriz, Vázquez, Royo, Nachit, Di Fonzo and Araus2000) and Moragues et al. (Reference Moragues, Zarco-Hernández, Moralejo and Royo2006).
There was no clear sub-regional or geographic trend in the distribution of diversity within the set of landraces studied. Among the most diverse landrace groups were those of the Iberian Peninsula. It is worth noting that in spite of the Spanish landraces being about three times more represented than the Portuguese, the genetic diversity indices of both countries were similar, and also similar to that previously reported by Moragues et al. (Reference Moragues, Zarco-Hernández, Moralejo and Royo2006) who found an index of 0·62 in a set of 25 landraces from the Iberian Peninsula. The lowest genetic diversity indices were found for the Algerian, Tunisian, Bulgarian, French and Turkish sub-groups. This is in agreement with the results reported by Hamdi et al. (Reference Hamdi, Bellil, Branlard and Khelifi2010) in a study with 856 accessions of Algerian durum wheat, and by Moragues et al. (Reference Moragues, Zarco-Hernández, Moralejo and Royo2006) which reported little diversity in Turkish and Bulgarian landraces. As suggested by Ganeva et al. (Reference Ganeva, Korzun, Landjeva, Popova and Christov2010), this could be attributed either to agro-ecological factors or to the efficient and consistent selection done by farmers through time, with the aim of improving the uniformity and yield of local populations.
The clustering based on the frequency of the null allele at Glu-A1, the most frequent either in modern varieties and landraces, displayed a clear geographical pattern. A very high frequency of this allele was detected in the modern varieties and in landraces from the Middle East and North African countries, France and Italy. With the exception of Egypt, in which this allele was found at a medium frequency, this geographic distribution is consistent with the dispersal route of durum wheat from the Fertile Crescent to North Africa through the south side of the Mediterranean Basin, as suggested by Moragues et al. (Reference Moragues, Moralejo, Sorrells and Royo2007). The close relationships between France and its former colonies in North Africa (Morocco, Algeria and Tunisia) would explain the French landraces being clustered in this group. Moreover, the inclusion of Italian germplasm in this cluster may be a consequence of the extensive use of North African landraces by Italian scientists and breeders during the first half of the 20th century (Di Fonzo et al. Reference Di Fonzo, Ravaglia, Deambrogio, Blanco, Troccoli, Royo, Nachit, Di Fonzo, Araus, Pfeiffer and Slafer2005). In contrast, all Balkan countries, as well as the Iberian Peninsula countries, Turkey and Egypt were clustered in a branch associated with a low frequency of the null allele at Glu-A1 that is consistent with a northern dispersal through the Mediterranean Basin.
The ANOVA for gluten strength showed that the component of variation due to genotype was more than eight times greater than the component due to the year effect, suggesting that this trait in durum wheat is largely under genetic control. The classification of genotypes in five groups according to their gluten strength determined by the SDS-sedimentation test was appropriate, as demonstrated by the large amount of the total variance explained by differences between groups (91·5%) and the lack of significant differences in gluten-strength values within groups.
None of the Glu-A1 alleles had a significant main effect on gluten strength, as determined by the SDS-sedimentation test, which is in line with the findings of Du Cros (Reference Du Cros1987), and different to the findings of other authors who found a positive relationship between Glu-A1 alleles and gluten strength when dough rheological parameters were considered (Ciaffi et al. Reference Ciaffi, Lafiandra, Turchetta, Ravaglia, Bariana, Gupta and Macritchie1995; Brites & Carrillo Reference Brites and Carrillo2001). Also, increases in gluten and dough strength have been achieved in durum wheat by transferring HMW-GS coded by Glu-A1 from Triticum dicoccoides (Ciaffi et al. Reference Ciaffi, Lafiandra, Turchetta, Ravaglia, Bariana, Gupta and Macritchie1995). In bread wheat the alleles Glu-A1a (1) and Glu-A1b (2*) have been shown to improve dough quality and strength compared to the Glu-A1c (null) allele (He et al. Reference He, Liu, Xia, Liu and Peña2005). However, the Glu-A1 subunits in bread wheat interact with a relatively different set of additional HMW-GS and a drastically different set of LMW-GS than those present in durum wheat, which may explain the difference in importance of this locus between the two species. It is also important to note that the SDS-sedimentation test is an indirect measure of gluten strength, based on the extent of aggregation/precipitation of the gluten polymer, not a direct physical measurement of viscoelastic properties of the gluten complex or dough. Further studies using dough rheology parameters may be needed to definitively determine actual strength differences associated with allelic variations at Glu-A1 in the group of landraces evaluated.
The present results showed that at Glu-B1, only banding pattern 20 had a significant main effect on the gluten strength of the landraces, while its effect in modern varieties was negligible. Previous studies addressing the effect of banding pattern 20 at Glu-B1 have reported contrasting results, ranging from a reduction of gluten strength (Peña et al. Reference Peña, Zarco-Hernandez, Amaya-Celis and Mujeeb-Kazi1994; Ammar et al. Reference Ammar, Kronstad and Morris2000; Brites & Carrillo Reference Brites and Carrillo2001; Sissons et al. Reference Sissons, Ames, Hare and Clarke2005) to a positive effect on bread volume, a trait highly and positively correlated with gluten strength (Boggini & Pogna Reference Boggini and Pogna1989), to no significant effects (Vázquez et al. Reference Vázquez, Ruiz, Nieto-Taladriz and Albuquerque1996). Again, these divergent results may be due to interactions among HMW-GS and LMW-GS in the formation of the gluten complex, and therefore main effects of a given allele depend on the overall glutenin combination in which it is present. This may be the case particularly for durum wheat in which LMW-GS effect dominates over the effect of the HMW-GS, making any HMW-GS main effect highly dependent on LMW-GS background (Peña & Pfeiffer Reference Peña, Pfeiffer, Royo, Nachit, Di Fonzo, Araus, Pfeiffer and Slafer2005). In the present study, the presence of subunit 7 at Glu-B1 had the most detrimental effect on gluten strength, but differences between gluten-strength groups were not significantly associated to its frequency, probably due to its low occurrence in the present population.
Two of the three alleles found at Glu-A3 in modern varieties had a significant and positive effect on the gluten strength in the landraces. The main effect of Glu-A3a on gluten strength was not significant in modern varieties, possibly because alleles from other loci with stronger main effects were present and did not allow the detection of the individual effect of Glu-A3a. Previous studies have related durum quality with the presence of Glu-A3a (Nieto-Taladriz et al. Reference Nieto-Taladriz, Ruiz, Martínez, Vázquez and Carrillo1997; Carrillo et al. Reference Carrillo, Martínez, Moita Brites, Nieto-Taladriz, Vázquez, Royo, Nachit, Di Fonzo and Araus2000). Allele Glu-A3d was more frequent in modern varieties than in landraces, and it had the largest positive effect on the SDS-values of landraces, but its effect in modern varieties was not significant, probably due to additive or complementary effects of individual Glu-1/Glu2/Glu3 alleles. However, allele Glu-A3b (band 5) absent in modern varieties and present in 13 landraces had the worst effect on gluten strength, in agreement with the findings of Carrillo et al. (Reference Carrillo, Martínez, Moita Brites, Nieto-Taladriz, Vázquez, Royo, Nachit, Di Fonzo and Araus2000). The data from the present study could not determine whether differences between gluten strength groups were associated with the frequency of this allele within each group.
The Glu-B3a allele had a positive effect on gluten strength in the landraces, while its effect on modern varieties was negligible. The relationship between good quality and the presence of Glu-B3a has been observed by other authors (Nieto-Taladriz et al. Reference Nieto-Taladriz, Ruiz, Martínez, Vázquez and Carrillo1997; Carrillo et al. Reference Carrillo, Martínez, Moita Brites, Nieto-Taladriz, Vázquez, Royo, Nachit, Di Fonzo and Araus2000).
Two rare banding patterns at Glu-B3 increased significantly the gluten strength in the landraces, 2 + 4+14 + 15 + 18 and 2 + 4+15 + 18 + 19. The first one was present in only one landrace (PI-366109 from Egypt), which SDS-value was 3·44 ml higher than the mean SDS-value of the whole population of landraces. A previous study (Nazco et al. Reference Nazco, Villegas, Ammar, Peña, Moragues and Royo2012) identified this landrace as having the highest EU quality index, due mostly to its very high gluten strength and protein content. Thus, it could be considered as a valuable gluten-strength enhancer in durum breeding programmes. The banding pattern 2 + 4+15 + 18 + 19 was detected in two landraces: Trigo Glutinoso and Lobeiro de Grao Escuro from France and Portugal, respectively, and its effect on SDS-value was estimated at 2·96 ml, greater than the main effect of any known allele evaluated in the present study. Both landraces have a high EU quality index (Nazco et al. Reference Nazco, Villegas, Ammar, Peña, Moragues and Royo2012), and the high quality of their protein, associated with the presence of the banding pattern 2 + 4+15 + 18 + 19 at Glu-B3, supports their use as parents in breeding programmes.
The two alleles detected at Glu-B2 locus had significant and opposite effects on the landraces gluten strength; Glu-B2a was found to exert a positive, but moderate effect on gluten strength. It was more frequent in modern varieties than in landraces, the opposite being true for the null allele Glu-B2b. Very few studies have been conducted on the relationship between Glu-B2 alleles and gluten strength, and generally the presence or absence of Glu-B2a did not result in any significant difference in gluten strength (Martinez et al. Reference Martinez, Ruiz and Carrillo2004). This was the case of the set of modern varieties used in the present study, in which the variation at this locus was not large enough to be statistically significant. However, the results obtained in the landraces, with much wider genetic background, generally weaker gluten overall and more opportunities to measure moderate effects, indicate that these alleles caused moderate and opposite effects on gluten strength.
The country-specific classification of the frequencies within the five gluten strength-groups offered a picture of the ‘native’ or ‘pre-green revolution’ gluten quality distribution around the Mediterranean Basin. Although a clear pattern was not evident, four of the five landraces with outstanding gluten strength, and seven of the 14 landraces with very high gluten strength came from eastern Mediterranean countries (Syria, Turkey, Cyprus and Egypt), providing an indication that the best gluten quality may have been concentrated originally in the Fertile Crescent, where tetraploid wheat originated (Feldman Reference Feldman, Bonjean and Angus2001). The Balkan Peninsula was characterized by a large number of landraces with poor gluten strength, probably associated to a different geographic origin (Nazco et al. Reference Nazco, Villegas, Ammar, Peña, Moragues and Royo2012). Western Mediterranean countries showed a wide diversity of gluten types, possibly due in part to their broad representation in the collection, but members with outstanding gluten strength were lacking in this sub-region. These results suggest that during the dispersal of wheat from the east to the west of the Mediterranean Basin, the alleles conferring outstanding gluten strength were probably not linked to any variability resulting in increased fitness or adaptation to the new environments and not associated with any local preference.
CONCLUDING REMARKS
Mediterranean landraces retain a wide genetic diversity for glutenin composition that has been mostly lost in modern varieties. They are a natural reservoir of alleles potentially useful to enhance and diversify gluten characteristics in durum wheat breeding programmes. Landraces from the Iberian Peninsula and Egypt were among the most diverse. The null allele was fixed at the Glu-A1 locus in the modern varieties studied, while four alternative banding patterns were recognized in the landraces. Only four of the 20 banding patterns identified in the landraces at the Glu-B1 locus were present in modern varieties. The present study confirms previous findings that the variability detected in durum wheat landraces for LMW-GS was wider than for HMW-GS, especially for the Glu-B3 locus. Only eight of the 89 banding patterns found in the landraces at Glu-2/Glu3 loci appeared in modern varieties. The clustering of the countries based on the frequency of the null allele at Glu-A1 was consistent with the proposed routes of durum wheat dispersal from the Fertile Crescent through the West of the Mediterranean Basin.
The medium-to-high values of gluten strength found in modern varieties reflects the efforts made by breeding programmes to improve grain quality. The low variation for gluten strength in modern varieties when compared with landraces could probably explain the lack of significant relationships between the 13 banding patterns identified in this group and gluten strength. Significant increases in the SDS-values of the landraces resulted from the presence of alleles Glu-B1e (band 20), Glu-A3a (band 6), Glu-A3d (bands 6 + 11), Glu-B3a (bands 2 + 4+15 + 19) and Glu-B2a (band 12) and the two hitherto unpublished banding patterns: 2 + 4+14 + 15 + 18 and 2 + 4+15 + 18 + 19 at Glu-B3 locus. However, landraces carrying the Glu-B2b (null allele), Glu-A3b (band 5) and the rare allele Glu-B1a (band 7) showed lower gluten-strength values, but only for Glu-B2b (null allele) could differences between the gluten-strength groups considered in the present study be associated with the allelic frequency.
Geographic distribution of gluten strength suggests that the best gluten quality probably originated in the Fertile Crescent and that, during the dispersal of wheat from the east to the west of the Mediterranean Basin, the alleles conferring outstanding gluten strength were frequently lost, likely due to their lack of association with other desirable traits such as yield or local adaptation.
R. Nazco was recipient of a Ph.D. grant from the Comissionat per Universitats i Investigació del Departament d'Innovació, Universitats i Empresa of the Generalitat of Catalonia and the Fondo Social Europeo. This study was partially funded by CICYT under projects AGL2006-09226-C02-01, AGL2009-11187 and AGL2012-37217 and was developed within the framework of the agreement between INIA Spain and CIMMYT. The Centre UdL-IRTA is part of the Centre CONSOLIDER INGENIO 2010 on Agrigenomics funded by the Spanish Ministry of Education and Science. CRF, ICARDA and USDA Germplasm Bank are acknowledged for providing accessions for the present study.



 Open access
Open access