



Introduction
Grain yield in oats is predominantly determined by grain number (Peltonen-sainio et al., Reference Peltonen-Sainio, Kangas, Salo and Jauhiainen2007; Finnan and Spink, Reference Finnan and Spink2017) with grain weight of lower importance. Annual cereal crops respond to environmental variation through changes in grain number with grain weight a more stable characteristic (Sadras, Reference Sadras2007). The response of oats to environmental variation is therefore reflected in the plasticity of the key yield components responsible for the determination of total grain number (Mahadevan et al., Reference Mahadevan, Calderini, Zwer and Sadras2016) with the following oat-specific hierarchy suggested: tiller number > grains/panicle > panicle number > grain weight, indicating the relative flexibility of tiller number and the stability of grain weight. This hierarchy may also transfer to the capacity of an oat crop to respond to changes in agronomic practices. Sowing date (Wall and Cartwright, Reference Wall and Cartwright1976; Crampton et al., Reference Crampton, Moot and Martin1997), cultivar (Brinkman and Rho, Reference Brinkman and Rho1984; Buerstmayr et al., Reference Buerstmayr, Krenn, Stephan, Grausgruber and Zechner2007), seed rate (Peltonen-sainio et al., Reference Peltonen-Sainio, Forsman and Poutala1997; Browne et al., Reference Browne, White and Burke2006; Finnan et al., Reference Finnan, Hyland and Burke2018), applied nitrogen (N) rate (Ohm, Reference Ohm1976; Browne et al., Reference Browne, White and Burke2006; Pecio and Bichonski, Reference Pecio and Bichonski2010; Finnan et al., Reference Finnan, Burke and Spink2019a) and plant growth regulator (PGR) (Leitch and Hayes, Reference Leitch and Hayes1989; Browne et al., Reference Browne, White and Burke2006) are all key agronomic parameters which have been shown to influence yield formation in oat crops. Establishing the impact of agronomic practices on the components contributing to total grain number and grain weight is a key target of this study.
Marshall et al. (Reference Marshall, Cowan, Edwards, Griffiths, Howarth, Langdon and White2013) note the unique prevailing climatic conditions in Ireland which facilitate the autumn planting of spring oat genotypes and leads to the production of the highest average oat yield in the EU-27. Information on the differences in yield formation processes of oats sown in the winter or spring is lacking and requires elucidation. Modern cultivars are characterized by a high grain number (Peltonen-sainio et al., Reference Peltonen-Sainio, Kangas, Salo and Jauhiainen2007) and an ability to maintain grain weight in an acceptable range (Calderini et al., Reference Calderini, Dreccer and Slafer1997) whereas there has also been a decrease in plant height in modern cultivars (Lynch and Frey, Reference Lynch and Frey1993) reducing lodging susceptibility. There is also debate as to whether increasing the seed rate of crops is economical due to the ability of cereal crops to achieve a yield stability across a range of seed rates (Finnan et al., Reference Finnan, Burke and Spink2019a, Reference Finnan, Burke and Spinkb).
The rate of applied N has been widely shown to be a key yield determining factor in oat production, (Chalmers et al., Reference Chalmers, Dyer and Sylvester-Bradley1998; Burke et al., Reference Burke, Browne and White2001; Browne et al., Reference Browne, White and Burke2003). This is associated with increases in total grain number through increases in panicles/m2 and spikelets/ear in oats with negative effects on single grain weight and increases in crop lodging (Chalmers et al., Reference Chalmers, Dyer and Sylvester-Bradley1998; Browne et al., Reference Browne, White and Burke2006; Finnan and Spink, Reference Finnan and Spink2017). Modern cultivars may have a reduced susceptibility to lodging at higher levels of applied N, which is the main deterrent of high levels of fertilizer N use on oat crops (Burke et al., Reference Burke, Browne and White2001). On the topic of the susceptibility of oat crops to lodging, White et al. (Reference White, Mc Garel and Ruddle2003) states that the application of PGR to oat crops is regarded as a necessity under Irish conditions. However, there are rising concerns about pesticide residues in food, with chlormequat frequently detected at levels below the maximum residue limit in oat samples (Reynolds et al., Reference Reynolds, Hill, Thomas and Hamey2004). Investigating the effect of applied N rate and PGR application on modern varieties forms a key element of this study.
The oat is quite unique amongst cereal crops due to its panicle-type inflorescence and the ability to adjust grain number during grain fill through the processes of grain site abortion and tertiary grain production (Browne et al., Reference Browne, White and Burke2006). The oat spikelet is also unique as it usually contains one to three florets that are in a primary, secondary and tertiary ranking, based on their position within the spikelet (White, Reference White and Welch1995; Doehlert et al., Reference Doehlert, McMullen, Jannink, Panigrahi, Gu and Riveland2005). Primary grains are the largest with kernel size reduced as the order increases. Takeda and Frey (Reference Takeda and Frey1980) and Doehlert et al. (Reference Doehlert, McMullen and Riveland2002) state that most oat spikelets contain two grains, containing primary and secondary florets. It has been accepted that the distribution of grain size in oat lots fits a bimodal distribution as a result of this physiological feature (Symons and Fulcher, Reference Symons and Fulcher1988; Doehlert et al., Reference Doehlert, McMullen and Riveland2002; Doehlert et al., Reference Doehlert, McMullen, Jannick, Panigrahi, Gu and Riveland2004). This variation in oat grain size presents difficulties for the oat milling processes as grains of different sizes will dehull at different rotor speeds (Deane and Commers, Reference Deane, Commers and Webster1986; White and Watson, Reference White and Watson2010). Increases in spikelet and total grain number are widely linked with an increased grain yield in oats (Peltonen-sainio et al., Reference Peltonen-Sainio, Kangas, Salo and Jauhiainen2007), but the influence that these increases may have on grain fill procedures within the spikelet is unclear. Finnan and Spink (Reference Finnan and Spink2017) suggested that grain yield in oats is limited by assimilate availability during grain fill. They further commented that increases in grain number are linked with an increased spikelet number at high yield levels rather than an increased level of secondary and tertiary grain formation, as demonstrated by Browne et al. (Reference Browne, White and Burke2003). Further examination into the procedures of grain filling at high grain yield levels is required in order to clarify this relationship between high grain number and grain filling procedures.
Browne et al. (Reference Browne, White and Burke2006) reported on the unique ability of the oat panicle to alter the number of potential grain sites by controlled abortion and the modification of the number of available tertiary grain sites based on the supply of assimilates during the grain fill period. Doehlert et al. (Reference Doehlert, McMullen and Riveland2002) reported that competition for photosynthate between a large grain number resulted in the abortion of grain sites in North Dakota. Browne et al. (Reference Browne, White and Burke2006) also reported cultivar and applied N rate effects on grain site abortion under Irish conditions. Identifying the grain fill procedures and levels of grain site abortion on oat panicles as potential grain number increases was a key target in the current study. Clarification of the role grain site abortion can play in maintaining grain yield during the periods of assimilate shortage will also be discussed.
The objective of the reported study was to re-evaluate the current understanding of the yield formation of oats under cool mild conditions, as typically experienced in Ireland. All yield components and panicle conformation measurements were recorded, with the examination of tertiary grain formation and post-anthesis abortion a key research focus.
Materials and methods
This research was conducted in the east of Ireland with full site details provided in Table 1. ‘Spring-sown’ oat crops were sown at two sites in 2016 and at one site in 2017 and 2018, with ‘winter-sown’ crops at a single site in 2017, 2018 and 2019. Therefore, comparisons between sites also involves different growing seasons and although the term ‘site’ is used through this document, the growing season aspect is recognized as being an important element within this. All sites had been in long-term continuous arable cropping for 15–20 years with optimal soil types for producing high yielding, high-quality oats. Soils with a possibility of a high mineralization of N were avoided based on cropping history (Wall and Plunkett, Reference Wall and Plunkett2016) (Table 2). Meteorological data including monthly precipitation and mean daily temperatures were obtained from Met Eireann (www.met.ie). Rainfall and temperature data were sourced from locations within 5 km of the experiments.
Table 1. Experimental site description

Table 2. Site fertility and crop development
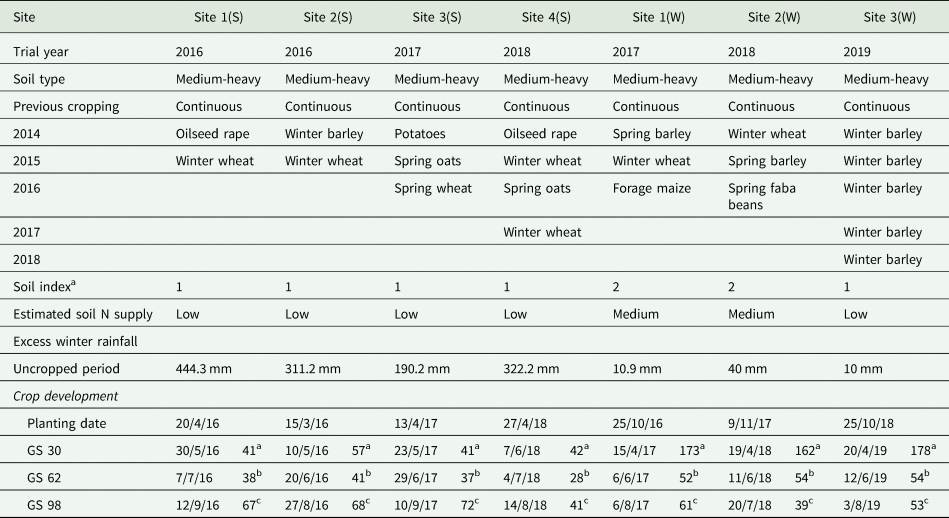
GS, growth stage; N, nitrogen.
a Time in days from planting to GS 30.
b Time in days from GS 30 to GS 62.
c Time in days from GS 62 to GS 98.
Experimental design
The experimental design was a split plot layout of three replications for the spring sowings in 2016 and winter sowings in 2017 at Backweston, with all other sowings having four replications. Main plots consisted of two cultivars (Husky and Keely) and three seed rates (250, 350 and 450 seeds/m2), with an applied N rate included as the split plot, and a +/− PGR application applied only to spring sowings. Plots were 15 m long with a 1 m discard to avoid fertilizer overlap.
Crop husbandry
Seedbeds were prepared using a reversible mouldboard plough to a depth of approximately 25 cm and a tilth was created using a power harrow. Seed was weighed out to target plant populations according to expected germination levels and thousand grain weight (TGW) values. All sowings were made by using a 1.5m Wintersteiger XL plot drill (Wintersteiger AG, Austria) equipped with Lemken double-disc seed coulters (Lemken GmbH, Alpen, Germany) with a row spacing of 150 mm. A single certified seed lot for each cultivar was used in each year. All spring sowings received insecticides for aphid control. Fungicides and herbicides were applied at recommended rates and timings.
In-field assessments and operations
All spring sowings received a pre-planting compound blend supplying 40 kg N/ha, 20 kg P/ha and 50 kg K/ha. Winter sowings received 50 kg N/ha, 20 kg P/ha and 60 kg K/ha in early spring when ground conditions facilitated fieldwork. The remaining applied N was applied at growth stage (GS) 21 (Zadoks et al., Reference Zadoks, Chang and Konzak1974) in the form of 27.5% calcium ammonium nitrate to bring the N allocation up to the prescribed total levels of 40, 70, 100, 130, 160 and 190 kg N/ha in spring-sown experiments and 80, 110, 140, 170 and 200 kg N/ha in winter-sown experiments. Plant growth regulation was applied as chlormequat chloride and Trinexapac-ethyl at 1.2 litres/ha + 0.1 litres/ha at GS 30 and GS 32/33 (Zadoks et al., Reference Zadoks, Chang and Konzak1974).
At 6 weeks after planting, all emerged shoots were counted along a 1 m stick in three locations/plot and average plants/m2 calculated by multiplying by a factor of 8 (eight rows/m). The number of panicles bearing stems was similarly recorded after anthesis. Prior to harvest, 15 panicles/plot were randomly selected for dissection of panicle components. After slow oven drying (40°C) to a constant dry weight, spikelet number per panicle and aborted spikelets per panicle were counted. Primary, secondary and tertiary grains were threshed, identified, weighed and counted. Total grain/panicle was calculated along with the numbers of aborted primary and secondary grains. Tertiary grain number is presented as the % of grains per panicle, with all grain site abortion data presented as the mean value per 15 panicles. Panicle conformation measurements were not recorded at site 2(S).
Plot yields were measured at full maturity using a Sampo Rosenlow 2025 plot combine (Pori, Finland), with an electronic weight cell. Where lodging occurred, harvesting was in the direction of lodging at slow speed to maximize grain recovery. TGW was calculated using a Contador seed counter (Pfeuffer Gmbh, Kitzingen, Germany). Grains/m2 data were calculated from panicle number and grains/panicle.
Statistical analysis
Bartletts test of homogeneity was conducted separately on the spring and winter data sets and it was found that the data were not homogenous for any variable due to drought conditions at site 4(S) and site 2(W). Each experiment was therefore analysed individually for all variables. Analysis was performed using the PROC MIXED procedure of SAS 9.4. Seed rate, applied N rate and PGR application were included as fixed effects with replication effects considered random. The model used was

where Y is the observation, μ is the overall mean, O, P and Q represent all possible two-, three- and four-way interaction between the agronomic variables, respectively. The main effects of these factors are presented in a tabular form with the probability of interactions between these factors also presented. These interactions are not included in the tabular form due to inconsistency across seasons and experiments.
Results
Climatic conditions
Monthly and 10-year average rainfall and mean temperatures for the spring- and winter-sown experiments are presented in Figs 1 and 2. Rainfall levels were similar at spring-sown sites 1(S), 2(S) and 3(S) whereas site 4(S) (2018) experienced moisture stress due to the lack of summer precipitation. High levels of August rainfall led to significant lodging at sites 1(S), 2(S) and 3(S). Total rainfall levels were similar between site 1(W) (2017) and site 2(W) (2018), but the pattern of rainfall varied greatly. Site 2(W) had high levels of winter precipitation and a period of moisture stress during the summer. Site 3(W) was a wet experimental site with high rainfall levels recorded throughout the season. The influence of temperature on crop development was also of importance, particularly in winter-sown experiments. Site 2(W) experienced a period of extreme cold in February and March relative to sites 1(W) and 3(W) where good temperatures encouraged higher early season growth. Temperature was relatively consistent across spring-sown experimental sites.

Fig. 1. Monthly rainfall values (mm) from March to September for spring-sown oats and October to August for winter-sown oats

Fig. 2. Monthly rainfall values (mm) from March to September for spring-sown oats and October to August for winter-sown oats
Grain yield and yield components
The effects of agronomic factors on grain yield, grains/m2 and TGW are presented in Table 3 (spring-sown oats) and Table 4 (winter-sown oats). Grain yield increases of 0.22–0.45 t/ha in the spring sowings and 0.4–0.8 t/ha in the winter sowings were observed as the seed rate increased from 250 to 450 seeds/m2. Similar increases were observed in response to the increased levels of applied N in all experiments. For spring sowings, the major increases were observed between 40 and 100 kg N/ha (0.7–1.5 t/ha) with an increase of between 0.1 and 0.6 t/ha observed as the applied N rate increased from 100 to 160 kg N/ha. The response in winter-sown crops was 0.5–2.04 t/ha as the rate of applied N increased from 80 to 170 kg N/ha. The benefit of applying additional N above 130 kg N/ha on spring-sown crops and 170 kg N/ha on winter-sown crops was limited and site specific in nature. The application of PGR increased the grain yield at sites 2(S), 3(S) and 4(S).
Table 3. Effect of agronomic factors on grain yield, grain number and TGW in spring-sown oats

n.s., not significant; N, nitrogen; PGR, plant growth regulator; SR, seed rate.
a Whole plot combine data.
b Calculated from yield and TGW data.
Table 4. Effects of agronomic factors on grain yield, grains/m2 and TGW of winter-sown oat

n.s., not significant; N, nitrogen; SR, seed rate.
a Whole plot combine data.
b Calculated from yield and TGW data.
Grains/m2 increased in all crops as both seed rate and applied N rate were increased. Increases of between 1000 and 2000 grains/m2 as seed rate increased in spring-sowings, with increases of 2000–2500 grains/m2 observed for winter sown. Increases following spring sowing, from 40 to 130 kg N/ha, were in the range of 3000–7000 grains/m2 with the additional increases beyond this point site-specific and in the range of 0–1000 grains/m2. A similar range was observed for winter sown, with increases of 3000 to 5000 grains/m2 over the range of 80 to 200 kg N/ha of applied N.
Increasing the seed rate had no effect on TGW in spring- or winter-sown oats. Increasing the level of applied N had a negative effect on TGW in all spring-sown crops and at site 3(W). The application of PGR also had a negative effect on TGW in all spring-sown crops.
The effects of agronomic factors on spikelet number per panicle, grains/panicle and panicles/m2 are presented in Table 5 (spring-sown oats) and Table 6 (winter-sown oats). There was a small reduction observed in spikelets/panicle in response to the increased seed rate at sites 1(S), 4(S) and 2(W). Spikelet number per panicle increased in response to higher levels of the applied N at sites 1(S) and 3(W) but no consistent trends were observed across all the experimental sites. Spikelet number per panicle was higher in winter-sown oats.
Table 5. Effects of agronomic factors on spikelets/panicle, grains/panicle and panicles/m2 of spring-sown oats

n.s., not significant; N, nitrogen; SR, seed rate; PGR, plant growth regulator.
a Mean value per panicle from 15 panicles per plot.
b Field plot observation.
Table 6. Effect of agronomic factors on spikelets/panicle, grains/panicle and panicles/m2 of winter-sown oats

n.s., not significant; N, nitrogen; SR, seed rate.
a Mean value per panicle from 15 panicles per plot.
b Field plot observation.
Grains/panicle were negatively impacted by the increased seed rate at sites 3(S), 4(S), 1(W) and 2(W). Increased levels of applied N rate resulted in significant increases in grains/panicle at sites 1(S) and 3(W) with the application of PGR increasing grains/panicle at site 3(S). Winter-sown oats produced higher numbers of grains/panicle.
Panicles/m2 linearly increased with the increased seed rate in all spring- and winter-sown crops. Increases in spring sowings were in the range of 39–72 panicles/m2 with increases of 30–55 panicles/m2 observed in winter sowings. Significant increases were noted in response to the increased applied N rate observed at all experimental sites. Panicle number increases in spring sowings were most profound between 40 and 130 kg N/ha and were in the range of 60–150 panicles/m2 excluding the site which experienced prolonged drought. Increasing the rate of applied N from 130 to 190 kg N/ha resulted in increases of 21–24 panicles/m2. Increases in winter-sown oats were notably site specific. Increases of 16–35 panicles/m2 were observed as the rate of applied N increased from 80 to 140 kg N/ha at sites 1(W) and 2(W) with increases of 92 panicles/m2 observed as the applied N rate increased from 80 to 200 kg N/ha at site 3(W). The application of PGR had no effect on panicle number in any spring-sown crop.
Panicle conformation
Proportion of primary grain weight
The effects of agronomic factors on the proportion of primary grain weight and number and % of tertiary grains per panicle is presented in Table 7 (spring-sown oats) and Table 8 (winter-sown oats). Agronomic effects on the proportion of primary grain weight were not generally significant in spring- or winter-sown oats. However, increases in the proportion of primary grain weight were observed in response to the applied N rate at site 3(S) with an increase observed at site 3(W) as the seed rate increased.
Table 7. Effect of agronomic factors on the proportion of primary grain weight and number and tertiary grain number in spring-sown oats

n.s., not significant; N, nitrogen; SR, seed rate; PGR, plant growth regulator.
a Values represent mean of 15 panicles.
b Values represent % of total grains.
Table 8. Effects of agronomic factors on the proportion of primary grain weight and number and tertiary grain number in winter-sown oats

n.s., not significant; N, nitrogen; SR, seed rate.
a Values represent mean of 15 panicles.
b Value represent % of total grains.
Proportion of primary grain number
Overall, the seed rate had no consistent effect on the proportion of primary grain number in spring-sown oats. Significant contrasting responses were observed in winter-sown oats, with a reduction observed at site 2(W) and an increase at site 3(W). Increasing the rate of applied N resulted in an increase in the proportion of primary grain number at sites 1(S), 3(S) and 1(W). The application of PGR reduced the proportion of primary grains by number only at site 1(S), and not in any other crop.
Tertiary grains
The % grains accounted by tertiary grains reduced as the seed rate increased at site 4(S) with no seed rate effects observed in any other experiment. Increasing the rate of applied N resulted in an increase in tertiary grains at site 3(W) and not at any other site. The application of PGR had no effect on tertiary grain number at any spring-sown site.
Aborted grain sites
The effects of agronomic factors on the number of aborted spikelets, primary grain and secondary grain sites are presented (Table 9, spring-sown oats; Table 10, winter-sown oats). Grain site abortion was reduced as the seed rate increased at site 3(S) with the number of aborted secondary grain sites reduced at site 4(S). There were interesting although insignificant increases in aborted primary and secondary grain sites as the seed rate increased at site 3(W). Increasing the rate of applied N reduced spikelet abortion at site 3(S) with an increase observed at site 3(W). Increases in the applied N rate also reduced the levels of primary and secondary grain site abortion at site 3(W). The application of PGR increased the level of aborted secondary grain sites at site 3(S) but this was not repeated in any other crop.
Table 9. Effect of agronomic factors on post-anthesis grain site abortion in spring-sown oats

n.s., not significant; N, nitrogen; SR, seed rate; PGR, plant growth regulator.
Values represent total number aborted grain sites per 15 panicles.
Table 10. Effect of agronomic factors on post anthesis grain site abortion in winter-sown oats

n.s., not significant; N, nitrogen; SR, seed rate.
Values represent total number aborted grain sites per 15 panicles.
Key correlations
The correlation data presented in Tables 11 and 12 provide a unique, novel insight into the complex and site-specific nature of grain site abortion and its role in the maintenance of grain yield in oats. Secondary grain site abortion was significantly associated (P < 0.05) with the proportion of primary grain weight and number at sites 1(S) and 3(S) (r = 0.17–0.25) indicating that a high rate of secondary grain site abortion occurred within treatments where primary grain sites were dominant. Observations from winter-sown sites also suggest that grain fill within spikelets is strongly related to assimilate availability during grain fill. At site 1(W) which was very high yielding (9.5–11 t/ha), secondary grain site abortion was negatively correlated with grain yield (r = −0.41) with a similar correlation observed at site 3(W) (r = −0.27). These correlations indicate that a higher level of secondary grains was formed at these higher yield levels. At site 2(W) aborted primary grain number was significantly associated with grain yield (r = 0.22), primary grain weight ratio (r = 0.22) and TGW (r = 0.23). This experiment was characterized by a prolonged drought period during grain fill and primary grains were aborted by the plant to ensure a sustainable amount of grain fill within remaining grain sites. At site 1(S) the yield was significantly associated (P < 0.05) with secondary grain site abortion indicating that remobilization of assimilates occurred from secondary grain sites into primary grain sites in order to maintain grain yield. The implications of these observations are two-fold. Firstly, the relationship between aborted secondary grains and primary grain weight and number ratios at spring-sown experiments indicates that assimilate supply was insufficient during grain fill and suggests that yield was source limited in these experiments. The high level of secondary grains observed at high grain yield levels at sites 1(W) and 3(W) suggests that yield was sink limited at this site. There is a level of complexity to these responses, linked to the length and climatic suitability of the grain-fill period. Secondly, this points to a phenomenon within spikelets that primary grains are prioritized by oat plants during periods of assimilate shortages.
Table 11. Correlations between key yield components in spring-sown oats

Values indicate Pearson's coefficient of correlation (r). *P < 0.05, **P < 0.01, ***P < 0.0001.
Table 12. Correlations between key yield components in winter-sown oats

Values indicate Pearson's coefficient of correlation (r). *P < 0.05, **P < 0.01, ***P < 0.0001.
Discussion
The results of this study has shown that seed rate, PGR application and applied N fertilizer rate influence yield formation in spring- and winter-sown oat crops, although the responses were not always consistent across all crops or sowing times, which creates complexity in interpreting the observations. Overall, crop structure and yield formation processes were predominantly determined by applied N rate with seed rate and PGR application displaying minor effects.
Yield components
Winter-sown oat crops yielded higher and more consistently than spring-sown crops with maximum yields of 9–10 and 5–9 t/ha, respectively. Winter-sown crops were characterized by higher spikelet and grain numbers per panicle than in the spring sowings. Although a direct comparison is not possible due to separate experimental sites/years the increase in yield potential in winter-sown oats is notable. The key determinant of yield potential in oats is grain number/m2 (Peltonen-Sainio et al., Reference Peltonen-Sainio, Kangas, Salo and Jauhiainen2007) with spikelet number per panicle in winter-sown oats increased due to the shortening of daylength and reduction in temperature during the winter period (Wall and Cartwright, Reference Wall and Cartwright1976). This slows down crop development and leads to an increase in spikelet primordia development and in yield potential (Kirby and Appleyard, Reference Kirby, Appleyard and Gallagher1984). In contrast, spring-sown crops develop rapidly as daylength increases in the spring, reducing the duration and rate of apex development leading to a lower spikelet number per panicle. This reduction in preformed yield potential was a key contributor to the observed lower yield response to applied N in the spring-sown crops.
The increases observed in panicles/m2 were in line with those reported by Peltonen-Sainio and Jarvinen (Reference Peltonen-Sainio and Jarvinen1995) and Finnan et al. (Reference Finnan, Burke and Spink2019a, Reference Finnan, Burke and Spinkb) with associated increases also observed in grains/m2. Winter sowing of spring cultivars can be expected to result in some level of plant death resulting in lower plants/m2. The wider range of observed increases in winter-sown experiments is therefore attributable to plant populations which are closer to or within the optimum range. Interestingly, the TGW of oats was relatively stable in response to seed rate when compared with the other key yield determining components, which is in line with Finnan et al. (Reference Finnan, Burke and Spink2019a, Reference Finnan, Burke and Spinkb). It appears that increasing the seed rate within a narrow range has a slight positive effect on total sink capacity, depending on the amount of tiller production, but has a major effect on the components of sink capacity, namely panicles/m2 and grains/panicle. As panicles/m2 increase in response to increased seed rate, grains/panicle decrease. However, individual grain weight is not as elastic and remains consistent in comparison within a normal plant population range as suggested by Sadras (Reference Sadras2007) and Finnan et al. (Reference Finnan, Burke and Spink2019a, Reference Finnan, Burke and Spinkb). Despite this, the study shows that yield increases of 0.22–0.45 t/ha were achievable in the spring sowings and 0.4–0.8 t/ha in the winter sowings. The benefits of using a high seed rate were linked to increases in both panicles/m2 and a reduction in the proportion of tiller ears in the crop. This is beneficial as panicles from tillers have been shown to have smaller grains and lower grains/panicle than mainstem ears (Peltonen-sainio and Jarvinen, Reference Peltonen-Sainio and Jarvinen1995; Finnan et al., Reference Finnan, Burke and Spink2019a, Reference Finnan, Burke and Spinkb). In addition, apical dominance will lead to the mainstem benefiting from the remobilization of assimilates contained in tiller ears which are aborted prior to anthesis (Palta et al., Reference Palta, Kobata, Turner and Fillery1994).
Increasing the applied N rate resulted in increases in grains/m2 in all crops. Interestingly, these increases were always attributable to increased panicles/m2 rather than grains/panicle, which only increased at two sites in response to higher N fertilizer. A number of previous studies have also reported increases in ears/m2 and grains/panicle in response to applied N rate, in winter wheat (Ellen and Spiertz, Reference Ellen and Spiertz1980, Efretui et al., Reference Efretui, Gooding, White, Spink and Hackett2016), spring barley (Easson, Reference Easson1984; Baethgen et al., Reference Baethgen, Christianson and Lamothe1995) and oats (Browne et al., Reference Browne, White and Burke2006; Finnan et al., Reference Finnan, Burke and Spink2019a). However, increases in grains/panicle and grain yield in response to N fertilizer application have also been reported previously in oats (Brinkman and Rho, Reference Brinkman and Rho1984; Pecio and Bichonski, Reference Pecio and Bichonski2010; Finnan et al., Reference Finnan, Burke and Spink2019a), indicating an underlying complexity in this yield to sink relationship in oats. It is suggested by Hay and Walker (Reference Hay and Walker1989) that grain weight is a factor of a source–sink relationship, particularly regarding total grain density and environmental stresses during grain fill. Grain sites will then compete for assimilates, resulting in lower individual grain weights. There is also a non-competitive theory (Miralles and Slafer, Reference Miralles and Slafer2007) that at higher grain numbers potential grain size is reduced as smaller grain sites are produced within spikelets. In oats, this would be linked to an increased detection of secondary and tertiary grain sites. The modern cultivars used in this study had high grain numbers but still produced TGW values in a consistent range suggesting that competition for assimilates is unlikely, particularly as crop growth rates have not improved in modern cultivars (Lynch and Frey, Reference Lynch and Frey1993). It has been hypothesized by Calderini et al. (Reference Calderini, Dreccer and Slafer1997) that modern cultivars have a higher ability to allocate assimilates to grain sites during grain fill. Grain weight reductions at high grain numbers in oat are therefore linked to both reductions in potential grain size and competition for assimilates at high grain numbers. Therefore, applied N fertilizer has a crucial role in increasing the grain yield of oats highlighted by the positive effect it had on grain number. However, the effect of applied N fertilizer on grain weight is an interesting phenomenon and demonstrates a biological trade-off from a yield formation perspective. Finnan and Spink (Reference Finnan and Spink2017) note that a yield maximization approach focused on the production of high grain numbers in oat will result in a high number of small grains with limited end-use quality. This is confirmed by the results of the current study.
The PGR application responses of neutral and positive effects on grain yield and components of total grain number but negative effects on grain weight are consistent with previous studies where lodging was not problematic (Green, Reference Green1986). At some sites (years) the increased yield was due to increased standing power and grain recovery or increased water use efficiency. This is corroborated by the studies of Leitch and Hayes (Reference Leitch and Hayes1990) and Chalmers et al. (Reference Chalmers, Dyer and Sylvester-Bradley1998) on standing power in oat and of Robertson and Greenway (Reference Robertson and Greenway1973) on water use efficiency in wheat. Additive effects of PGR application on grain number are generally associated with a reduction in grain weight to have an ultimately neutral effect on grain yield (Green, Reference Green1986). Positive yield responses to PGR in the absence of lodging are uncommon but not unusual. In the current study, panicles/m2 only increased in response to PGR application in an extremely dry season, whereas grains/panicle increased in response to PGR application in a crop with high panicle numbers. Reductions in grain weight in oats in response to PGR application were reported by Leitch and Hayes (Reference Leitch and Hayes1989) and Browne et al. (Reference Browne, White and Burke2006). This decrease has been linked to an increased sink capacity and an increase in competition between potential grain sites in wheat crops by Lowe and Carter (Reference Lowe and Carter1972) and Green et al. (Reference Green, Hawkins and McDonald1985) noted that chlormequat application reduced the rate of grain growth in the post anthesis period and instead caused redistribution of post anthesis assimilate into non-reproductive organs. It is probable that the reductions in grain weight caused by PGR application were a combination of both responses. Where additional grain sites were not stimulated by the application of PGRs it is probable that grain weight was reduced due to assimilate being redirected from grain sites. These results fit in with the theory suggested by Rajala (Reference Rajala2004) who, after a wide-ranging review, concluded that varying responses in oat crops to PGR application were case specific. Although the application of PGR can exert positive effects on yield components, these effects are not as reliable or consistent as other agronomic factors. Therefore, the application of PGR, although required to reduce the risk of crop lodging and increase grain recovery under Irish conditions, should not be claimed as a means of increasing yield per se.
Panicle conformation
Increasing seed rates resulted in significantly lower spikelet number/panicle in all spring sowings and at sites 1(W) and 2(W) in line with Peltonen-sainio and Jarvinen (Reference Peltonen-Sainio and Jarvinen1995). Spikelet numbers were unresponsive to increasing N rate while panicle number was generally higher, indicating that increased N fertilizer increased panicles/m2 rather than grains or spikelets per panicle. Increasing the level of applied N increased the primary grain weight and number ratios in all spring sowings but only increased the primary grain number in one winter sowing. This demonstrated that increases in grains/m2 in response to increased N rate were associated with an increase in the proportion of primary grain types. Notably, the results from Browne et al. (Reference Browne, White and Burke2006) where spikelet number and grain number increased while the primary grain number ratio was unaffected, demonstrates that increases in grain number in response to applied N are not linked to the increased levels of secondary and tertiary grain fills. An interesting response was observed at site 3(S) where the proportion of primary grain by number was <0.5. This response initially seems strange, but spikelets and grains/panicle were low at this site relative to other spring-sown sites. A more consistent distribution of assimilates between primary, secondary and tertiary grains is therefore observed at this site, as intra-panicle competition for assimilates is low. This explains the proportion of primary grain number falling below 0.5, as secondary and tertiary grains outnumbered primary grains.
Grain site abortion
The influence of agronomic factors on grain site abortion was inconsistent but detectable in all crops and was strongly associated with climatic conditions during grain fill. Oats are unique in that aborted grain sites are easily detectable on the harvested panicle due to the structure of the spikelet, but little published research is available to corroborate this. An important note here is that the presence of grain site abortion in tandem with tertiary grain sites at sites is linked to short stress periods of low assimilate availability during grain fill, rather than overall sink or source limitation. Short, late periods of moisture availability during the period of moisture stress at site 4(S) may have coincided with flowering in tertiary grains, leading to the formation of a high number of tertiary grains when no other sinks were available. A high spikelet number/panicle was formed at site 3(W) with a high level of spikelet and grain site abortion observed, as grain fill was limited due to a high potential grain number. Although spikelet abortion increased with the applied N rate, the negative relationship observed between the applied N rate and primary/secondary grain abortion demonstrates that an increase in assimilate availability has reduced grain abortion as grain fill progressed. Interestingly, the abortion of spikelets insufficiently readjusted the assimilate balance on the panicle as grain site abortion was still required in a lower number of spikelets.
As the seed rate increased grain site abortion remained similar or decreased at spring-sown sites. This was linked to a lower number of grains/panicle and a higher proportion of mainstems in high seed rate plots which can be adequately filled by the plant. Grain sites on tillers may also be aborted prior to mainstems as competition for assimilates increases during grain fill. Tillers are used as a source of assimilates during pre-anthesis (Peltonen-sainio and Jarvinen, Reference Peltonen-Sainio and Jarvinen1995) and may be used in a similar fashion during grain fill due to apical dominance of the mainstem. The application of PGR does not appear to provide any abortion protection and was actually associated with an increase in secondary grain site abortion in one spring crop.
Browne et al. (Reference Browne, White and Burke2006) reported increases in primary and secondary grain site abortion in response to the increased levels of applied N fertilizer under Irish conditions. The current study provided a more complex pattern of raised and lowered primary and secondary grain site abortion rates and spikelet abortions rates across the different sites/years and sowing dates. Oats produce a higher number of spikelets than they can adequately fill (Peltonen-sainio and Peltonen, Reference Peltonen-Sainio and Peltonen1995) and so the differences in responses can be explained by the differences in the length of the pre-anthesis period between spring- and winter-sown oats, differences in crop growing conditions and the potential of applied N to increase spikelet number per panicle. For example, the increase in spikelet abortion at site 3(W) can be attributed to a higher initial spikelet number resulting from a longer pre-anthesis phase in this winter-sown crop and an associated increase in competition from potential grain sites for assimilate immediately following anthesis. The assimilates remobilized from these spikelets in tandem with higher assimilate availability may have contributed to a reduction in primary and secondary grain site abortion as grain fill progressed. This would be less likely to occur in spring-sown oats, as they grow rapidly as daylength increases with a lower number of spikelets formed as a result. Therefore, this explains the observed differential responses between different crops. Increasing the level of N application would increase assimilate availability and reduce the need for remobilization of nutrients just prior to anthesis, with the extension of the grain fill period lowering abortion rates amongst a lower number of spikelets in spring-sown crops. Similarly, the early abortion of surplus spikelets in the winter-sown crops reduced the level of demand from grain sites with the longer period of grain fill better equipped to increase grain fill amongst a smaller pool of potential grain sites. The differential responses in the spring-sown crops is also understandable, as Browne et al. (Reference Browne, White and Burke2006) have shown that the grain fill period in spring crops is shorter with abortion of grain sites required to ensure consistent grain fill within the remaining spikelets on the panicle. Therefore, it can be concluded that the effect of applied N rate on grain site abortion is highly linked to climatic conditions and the physiological time at which assimilate becomes limiting.
There is a hierarchal ranking for abortion of grain sites during the development of the oat panicle, which proceeds from the uppermost terminal spikelet to the base of the panicle with pollination proceeding in the same order across a period of 10–11 days (Rajala and Peltonen-Sainio, Reference Rajala and Peltonen-Sainio2011). Grains at the uppermost whorl are also typically bigger than grains from whorls closer to the base (White and Finnan, Reference White and Finnan2017). This can lead to a situation where grains at the uppermost panicle have moved through anthesis with grains at the lowest whorl only commencing anthesis. In crops characterized by a high spikelet number the demand for assimilates from grains at the uppermost whorl may trigger the abortion of spikelets at an early development stage. Once the plant has determined its total fertile spikelet number it begins the allocation of assimilates into primary and secondary florets within each spikelet. Finnan and Spink (Reference Finnan and Spink2017) note that increases in grain number due to a higher spikelet number per panicle is not predominantly linked with an increase in the number of secondary grains per spikelet, noting the potential for a high number of single-grain spikelets. The underlying multifaceted nature of this fluid relationship between abortion and maintenance of grain yield is clearly associated with the pattern of correlations observed between the key yield components, and so provides a unique insight into these processes.
Overall, the current study firmly reinforces the theory of elasticity of grain fill proposed by Browne et al. (Reference Browne, White and Burke2006). The associations between the proportion of primary grain weight and aborted secondary grains indicate that yield was source limited. The winter-sown sites are of high importance in this respect. Where grain yield was at its highest secondary grain abortion was low and all available grain sites were then filled, leading to the high yield achieved. During the period of moisture stress at site 2(W), when primary grain site abortion was significantly associated with grain yield, this indicates that remobilization of assimilates had occurred to maintain total grain yield. These results also support a hierarchy of grain fill during the periods of assimilate shortages with primary grain florets preferred. In order to achieve maximum yield levels, a consistent level of secondary grain fill is required, demonstrated by the results from site 1(W). Griffiths (Reference Griffiths2010) reported that the primary grain weight ratio was negatively correlated with grain yield (r = −0.65) noting that a high degree of secondary grain fill was required to achieve high yields. This is applicable when climatic and agronomic conditions are at their optimum during grain fill as all grain sites can be adequately filled, such as site 1(W). In cases where the plant lacks the photosynthetic capacity to fill all its grains, grain sites were aborted to ensure consistent grain fill within a smaller pool of potential grain sites.
Conclusion
Increases in panicle number accounted for yield increases in response to seed rate and N rate regardless of sowing date. This study demonstrated that oat grain yield under cool, humid conditions is associated with high levels of secondary grain fill. During seasons characterized by moisture stress, grain site abortion played a key role in the maintenance of crop yield by ensuring a base level of primary grain fill within each spikelet. Once the number of fertile spikelets is determined at anthesis, assimilate allocation is prioritized for primary grains to ensure at least one per spikelet survives. This study has also demonstrated an important relationship between spikelet number at anthesis and the suitability of grain fill conditions on the processes of grain site formation and the post-anthesis abortion of grain sites. A continued move to winter sowing will improve the yield potential and consistency of crop performance under Irish conditions.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S0021859621000320.
Acknowledgements
The authors thank Mr Eugene Brennan.
Financial support
This study was supported by Irish Research Council and Origin Enterprises PLC.
Conflict of interest
The authors declare there are no conflicts of interest.

















 Open access
Open access