Introduction
Sheep farmers produce meat, milk, wool/hair, and skins for national and international markets (Morris, Reference Morris Stephen2017). In the world, more than 1.2 billion sheep are being produced globally each year and 0.40 of the effective sheep population is found in semi-arid regions, with greater concentration in India, the Middle East, the highlands of East Africa and South America (Zygoyiannis, Reference Zygoyiannis2006; IWTO, Reference Dilworth2022).
In these regions, the systems used for raising sheep vary between and within different countries. There are three major sheep production management systems in the world, namely extensive, semi-extensive and intensive production. Generally, the different breeding systems can provide good results (Morris, Reference Morris Stephen2017). However, to be efficient in the sheep production system, a high survival rate of the lambs is necessary, which is influenced by the temperament of the ewe, the vigour of the newborn, and other factors related to the environment, e.g. nutrition, human-animal interaction, facilities and thermal environment (Costa et al., Reference Costa, Almeida, Pimenta Filho, Junior and Santos2008; Silva et al., Reference Silva, Santos, Kohek, Machado, Hein, Vidor and Corbellini2013; Araújo da Silva et al., Reference Araújo da Silva, Saraiva, Bispo, Fonsêca, Gonçalves dos Santos, Dos Santos and Almeida2020). The exit of the newborn from the uterine to the external environment requires profound behavioural, physiological, and neurological adjustments in the ewe and newborn (Lanfranchi et al., Reference Lanfranchi, Calabrò, De Pascale, Fazio and Giannetto2016). When compared to adult animals, newborn lambs are more susceptible to the fluctuations in the thermal environment, as they born wetted, and has a greater body surface area to volume ratio (Jones, Reference Jones1997; Laburn, Reference Laburn and Blatteis2001; Lezama-García et al., Reference Lezama-García, Mota-Rojas, Martínez-Burnes, Villanueva-García, Domínguez-Oliva, Gómez-Prado and Muns2022).
Semi-arid equatorial zones have marked daily variations in air temperature (e.g. 15 to 40°C), high levels of mean radiant temperature, and solar radiation (Fonsêca et al., Reference Fonsêca, Saraiva, Pimenta Filho, Furtado, Mariz, Silva and Pinheiro2014). In equatorial semi- arid areas, newborn lambs delivered at night can face with cold stress, where thermal radiant conditions favour the transference of sensible heat loss to be greater than the metabolic heat produced by metabolism (Fonsêca et al., Reference Fonsêca, Maia, Saraiva, de Melo Costa, da Silva, Abdoun and Fuller2019). Newborn lambs from hair coat sheep breeds were more lethargic (i.e. lambs took longer to perform the first suckling) when delivered at circumstances of air temperature between 18 and 23°C (Fonsêca et al., Reference Fonsêca, Saraiva, Pimenta Filho, Furtado, Mariz, Silva and Pinheiro2014). Indeed, the hypothermia and starvation are reported to account for 10% of neonatal lamb mortality in tropical semi-arid regions (Riet-Correa and Méndez, Reference Riet-Correa and Méndez2001; Nóbrega Jr. et al., Reference Nóbrega, Riet-Correa, Nóbrega, Medeiros, Vasconcelos, Simões and Tabosa2005).
Neonatal lamb mortality does not have one specific cause. Primary risk factors include low birth weight, dystocia, competition between siblings in multiple births, adverse weather conditions, and poor formation of mother-young bond (Riet-Correa and Méndez, Reference Riet-Correa and Méndez2001; Medeiros et al., Reference Medeiros, Tabosa, Simões, Nóbrega Júnior, Vasconcelos and Riet-Correa2005; Nóbrega Jr et al., Reference Nóbrega, Riet-Correa, Nóbrega, Medeiros, Vasconcelos, Simões and Tabosa2005; McCoard et al., Reference McCoard, Henderson, Knol, Dowling and Webster2014). For example, cleaning the lambs by the ewes after birth is crucial to prevent the rapid evaporation of fluids and heat transference from the skin surface of lambs to the surrounding environment. The thermoregulatory behaviour of ewes also plays an important role, as it directly influences lambs’ motivation for seeking more favourable microclimate (e.g. seeking shelter or shade) in order to avoid extreme cold or heat (Terrien et al., Reference Terrien, Perret and Aujard2011; Fonsêca et al., Reference Fonsêca, Saraiva, Pimenta Filho, Furtado, Mariz, Silva and Pinheiro2014; Ferner et al., Reference Ferner, Schultz and Zeller2017; Lezama-García et al., Reference Lezama-García, Mota-Rojas, Martínez-Burnes, Villanueva-García, Domínguez-Oliva, Gómez-Prado and Muns2022).
While several studies have evaluated the negative impact of extreme cold weather on vigour of newborn lambs from wool breeds in temperate regions (Steyn, Reference Steyn2022), the heat stress as risk factor for neonatal lamb mortality has not received attention. However, in equatorial semi-arid regions, where levels of solar radiation are high and nearly constant throughout the year, newborn lambs can absorb high amount of radiant heat and face with challenges to maintain their thermal equilibrium (Fonsêca et al., Reference Fonsêca2016). For instance, newborn lambs artificially exposed to radiant temperature of 40°C kept their body temperature within narrow range, but at a cost of excessive rise in respiratory rate (Mercer et al., Reference Mercer, Andrews and Szekely1979). Recently, field studies conducted in equatorial semi-arid regions reported that newborn lambs from hair coat sheep breeds were moderately hyperthermic when exposed to high levels (Up to 1000 W m−2) of direct solar radiation (Santos, Reference Santos2020). In this review, we aimed at investigating the following topics: (I) Characterization of sheep farming systems in semi-arid regions; (II) to discuss the main risk factors for neonatal lamb mortality; (III) and aspects of thermoregulation of newborn lambs.
Sheep production system in semi-arid regions
The sheep was domesticated over 10 000 years ago during the Neolithic in Central Asia. Domesticated sheep underwent changes from wild shaggy sheep to Mesopotamian fleeced and furred domestic sheep. In Asia, Asian sheep, which originated from the mouflon (Ovis Orientalis), lead to the divergence of ovine genetic resources due to their recruitment from the wild several times for domestication (Hiendleder et al., Reference Hiendleder, Kaupe, Wassmuth and Janke2002; Bruford and Townsend, Reference Bruford and Townsend2006; Rezaei et al., Reference Rezaei, Naderi, Chintauan-Marquier, Taberlet, Virk, Naghash and Pompanon2010). Currently, there are 1.2 billion sheep in the world (IWTO, Reference Dilworth2022; Figure 1). Asia accounts for over 0.42 of all sheep in the world, followed by Africa, Europe, Oceania, and the Americas with 0.31, 0.11, 0.08 and 0.07. (Pulina et al., Reference Pulina, Milán, Lavín, Theodoridis, Morin, Capote and Caja2018). Among countries, China leads the ranking with 0.14, followed by Australia, India, Nigeria, and Sudan, with 0.057, 0.050, 0.035 and 0.033, respectively (Toldrá, Reference Toldrá2022).

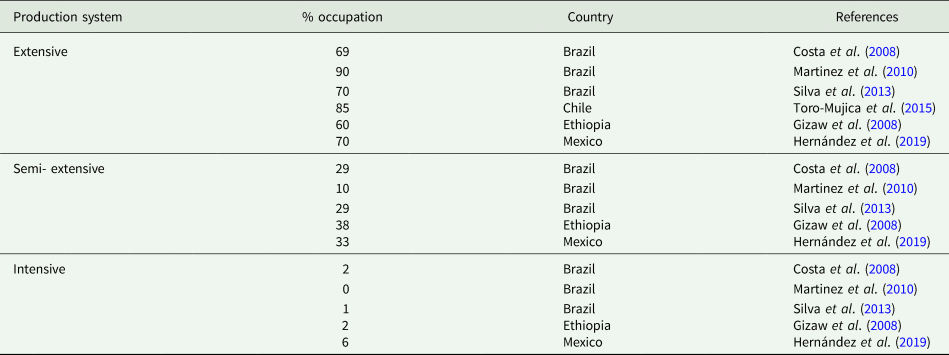
The utility of sheep farming in the world is multidimensional, providing a variety of products (e.g. meat, milk, and wool) in a wide spectrum of sociological and socioeconomic conditions. Since its emergence, most production systems were linked to subsistence, with creations in fragile lands. Over the years, this has become multifactorial, as climatic conditions, stocking rate, available area, forage availability, and quality will determine the variation of production systems, which can be characterized by the set of technologies and management practices, which lead to considering the type of animal, breed, the purpose of breeding and region where the activity is carried out (Holanda Júnior, Reference Holanda Júnior2006). Currently, sheep farming has commercial objectives, which are characterized by the production of food of high biological value (Moraes Neto et al., Reference Moraes Neto, Rodrigues, Albuquerque and Mayer2003; Braga and Rodriguês, Reference Braga and Rodriguês2005). Despite the expansion and evolution of systems, a large part of domestic sheep farming is still done extensively and semi-extensively, which is characterized by low reproductive performance, low level of organization, poor use of husbandry techniques and zootechnical bookkeeping, in addition to seasonality in the availability of water and food resources (Costa et al., Reference Costa, Palmeira, Silva, Furtado, Dantas and Dos Santos2014; Simioni et al., Reference Simioni, Gomes, Mousquer, Teixeira, de Castro, de Paula and Fernandes2014; Table 1).
Table 1. Percentage of total occupancy of the production system (% occupation) adopted in the different regions of the world as a function of the total number of sheep herds
It is estimated that 0.43 of the world's surface is arid, where 0.40 of the world's sheep population is found (Kassas, Reference Kassas and Dregne1975; Harrington, Reference Harrington1981). Arid and semi-arid regions can be defined as areas where precipitation, relative to the level of evapotranspiration, is inadequate to sustain reliable agricultural production (Meigs, Reference Meigs1953). Precipitation in these areas is generally low and/or erratic. The vegetation consists of grasslands, shrublands, savannahs, or woodlands, but may also be covered by desert (Hill and Guerschman, Reference Hill and Guerschman2020). Anatomical, behavioural, and physiological characteristics reflect some specificities in the energy balance of native sheep in arid and semi-arid areas (Fonsêca et al., Reference Fonsêca, Maia, Saraiva, de Melo Costa, da Silva, Abdoun and Fuller2019). These animals deal with frequent challenges such as low availability of water resources, supply and quality of food, high levels of solar radiation, and seasonal and daily temperature fluctuations.
Both in the extensive and semi-extensive systems, the animals usually remain in pasture areas during the day and are collected for installation at night, becoming more susceptible to environmental weather conditions during the daylight. In conditions of equatorial semi-arid environment, high solar radiation and mean radiant temperature are the principal factors that affect the thermoregulation of animals, especially when they do not have access to shade (Maia et al., Reference Maia, Silva and Loureiro2008; Baêta and Souza, Reference Baêta, Souza and Araújo2010; Turco and Araújo, Reference Turco, Araújo and Voltolini2011). Under such circumstances, during lambing seasons, the radiative properties are decisive in the thermoregulation processes. In new-born animals, the pigmentation of the fur, characteristics of the covering layer, and body size are decisive for radiant thermal changes. Studies with adult animals have observed that animals with a dark external surface are more subject to heat stress than those of light colour, presenting greater absorbance of different wavelengths. In contrast, lighter hairs show greater reflectance, absorbing smaller amounts of thermal energy (Gebremedhin and Hillman, Reference Gebremedhin and Hillman1997; Silva et al., Reference Silva, Lascala and Tonhati2003; Maia et al., Reference Maia, Silva and Bertipaglia2005). The small body size and a relatively large surface about its mass are striking characteristics in new-born animals, making them efficient heat sinks and more susceptible to variations in the thermal environment, when compared to adult animals (Cain et al., Reference Cain, Krausman, Rosenstock and Turner2006; McManus et al., Reference McManus, Paludo, Louvandini, Gugel, Sasaki and Paiva2009).
Due to the economic losses generated by the sheep production system, the survival of lambs has become a worldwide concern. However, attempts to reduce lamb mortality in recent years (1970–2014) have not changed significantly, remaining at an average of 15% in many countries (Fig. 2). For example, in New Zealand and Uruguay, neonatal deaths account for more than 30% (Sassi and McCosker, Reference Sassi and McCosker1975; Aspin, Reference Aspin1997); whereas in England and Australia, deaths vary from 2 to 21% (McFarlane, Reference MCFarlane1965; Stamp, Reference Stamp1967). In addition, in other semi-arid regions of the world, mortality rates exceed 30% (e.g. Ethiopia, 46 to 51%, Bekele et al., Reference Bekele, Kasali and Woldeab1992; Morocco, 18 to 31%, and Ghana 33%; Chaarani et al., Reference Chaarani, Robinson and Johnson1991).
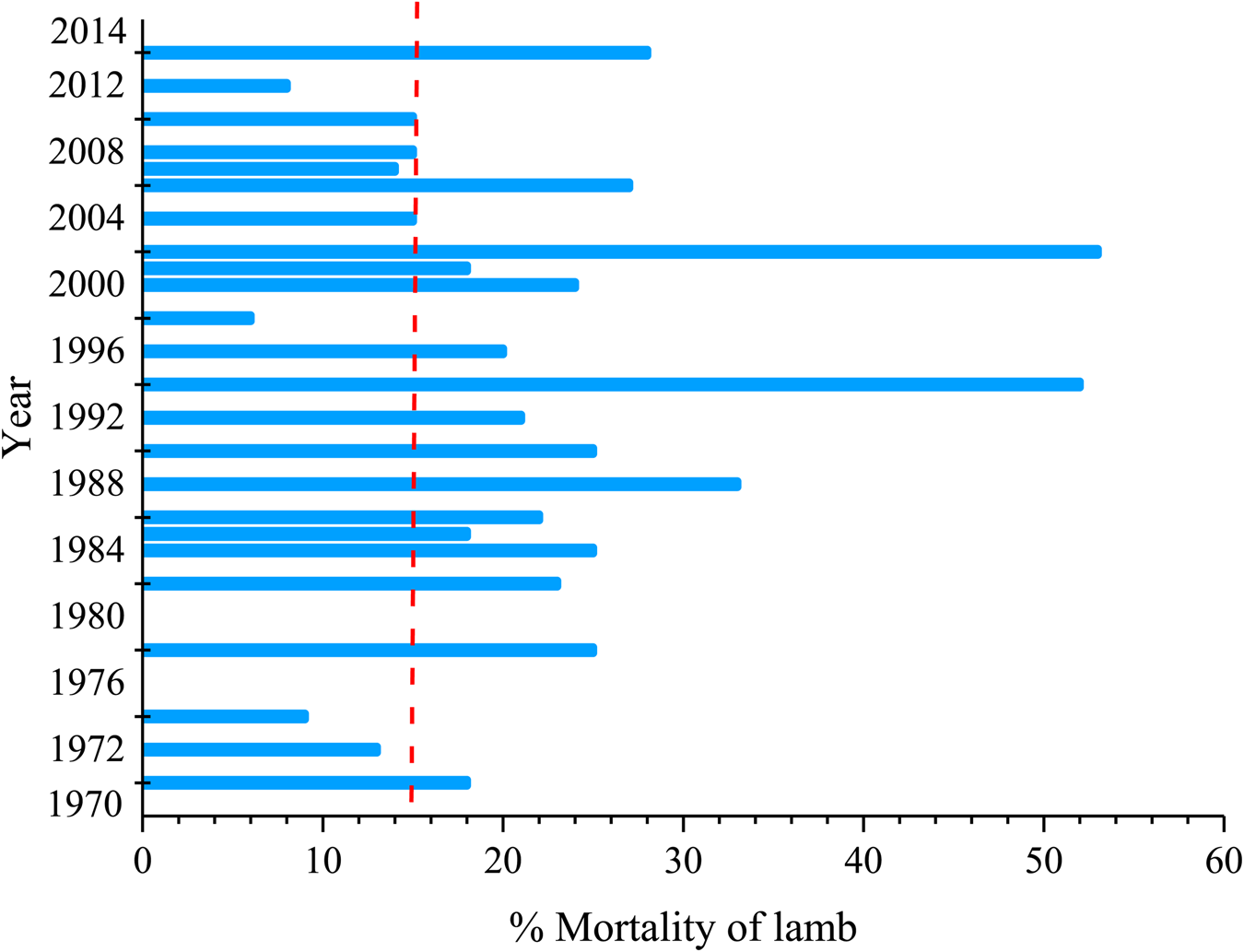
Figure 2. The average percentage of neonatal mortality in the world between 1970 and 2014. The dotted line refers to the average rate (Adapted from Dwyer et al., Reference Dwyer, Conington, Corbiere, Holmøy, Muri, Nowak and Gautier2016).
Considering neonatal mortality rates with animals reared in semi-arid zones, the scenario is practically the same. In studies with Santa Ines ewes with and without previous parturition experience, Simplício et al. (Reference Simplício, Riera, De Figueiredo and Nunes1982) reported mortality rates ranging from 5 to 19%, respectively. Lima (Reference Lima1983), when working with Santa Ines, Morada Nova, and Somalis sheep in the Northeastern semi-arid region, mentions rates of 13, 18, and 9%. Girão et al. (Reference Girão, Medeiros and Girão1998) cite rates of 15% for Santa Ines lambs. Nóbrega Jr. et al. (Reference Nóbrega, Riet-Correa, Nóbrega, Medeiros, Vasconcelos, Simões and Tabosa2005) observed that 4% of deaths would occur before birth, 30% in the first 24 h, 20% from the first to the third day of life, and 35% after 3 days of life. Fragkou et al. (Reference Fragkou, Mavrogianni and Fthenakis2010) state that the optimal neonatal mortality rate should not exceed 3%, with 5% being acceptable. However, such values are not obtained in most sheep production systems in the world. Thus, for the activity to be based on good management practices and maintenance of acceptable levels of animal welfare, it is extremely important to look at the aspects that interfere with neonatal mortality, especially in production systems with a low technological level.
Risk factors that interfere with lamb survival in the first hours of life
Neonatal mortality is a complex problem in sheep farming, not have a single specific cause (Oliveira and Barros, Reference Oliveira and Barros1982). The main causes of neonatal mortality are birth weight, dystocia, sibling competition in multiple births, adverse weather conditions, inadequate supply of milk or colostrum (e.g. starvation/hypothermia complex), infections, and poor ewe behaviour and/or lamb. In diagnostic necropsies performed in new-born lambs around the world, from 1970 to 2022, several authors described the main causes of perinatal mortality (Table 2).
Table 2. Main causes of perinatal mortality of necropsied lambs in the world

Birth weight is a major contributor to neonatal mortality (Yapi et al., Reference Yapi, Boylan and Robinson1990; Fogarty et al., Reference Fogarty, Hoplins and Van de Ven2000), e.g. lambs with low birth weight and from twin births are more susceptible to hypothermia and starvation (Scales et al., Reference Scales, Burton and Moss1986; Ribeiro et al., Reference Ribeiro, Gregory and Mattos2002). In contrast, heavy lambs from simple births may also be more susceptible to mortality, especially due to dystocia (Dalton et al., Reference Dalton, Knight and Johnson1980). Dalton et al. (Reference Dalton, Knight and Johnson1980) described a correlation between birth weight and mortality of new-born lambs, where the mortality rate of lambs weighing between 2.0 to 2.5 kg was 48%, while lambs weighing between 4.0 to 2.5 kg 5.5 kg was 14%, and 22% for the heaviest (i.e. >6.0 kg). Similarly, in studies with Barki sheep, Sallam (Reference Sallam2019) observed a higher neonatal mortality rate in the lighter animals (19%), followed by those of intermediate weight (6%) and heavier (3%). Dwyer et al. (Reference Dwyer, Conington, Corbiere, Holmøy, Muri, Nowak and Gautier2016) determined a ‘U’ relationship between birth weight and the neonatal survival rate, which shows the interval of 3 to 5 kg as being of optimum birth weight.
Worldwide, the overall proportion of lamb mortality attributable to dystocia (pooled proportional mortality rate) is 47% (95% CI: 38%; 55%; Figure 3; Bruce et al., Reference Bruce, Young, Masters, Refshauge, Thompson, Kenyon and Jacobson2021) (Hall et al. Reference Hall, Fogarty and Gilmour1995; Holst et al. Reference Holst, Fogarty, Hopkins and Stanley1997, Reference Holst, Fogarty and Stanley2002; Sargison et al. Reference Sargison, West and Clark1997, Reference Sargison, West and Clark1998; Kerslake et al. Reference Kerslake, Everett-Hincks and Campbell2005; Everett-Hincks et al. Reference Everett-Hincks, Mathias-Davis, Greer, Auvray and Dodds2014; Behrendt et al. Reference Behrendt, Edwards, Gordon, Hyder, Kelly, Cameron and Thompson2019; Lockwood et al. Reference Lockwood, Hancock, Kearney and Thompson2019a, Reference Lockwood, Hancock, Paganoni, Macleay, Kearney, Sohi and Thompson2019b). Dystocia can be caused by hereditary, nutritional, managerial, infectious, traumatic factors, or a combination of these (Roberts, Reference Roberts1971). For example, in a study with Morada Nova sheep, Fonsêca et al. (Reference Fonsêca, Saraiva, Pimenta Filho, Furtado, Mariz, Silva and Pinheiro2014) and Santos et al. (in preparation) observed that dystocia lambs were slower to get up and perform the first feeding; in addition, they received less intensity of maternal care.
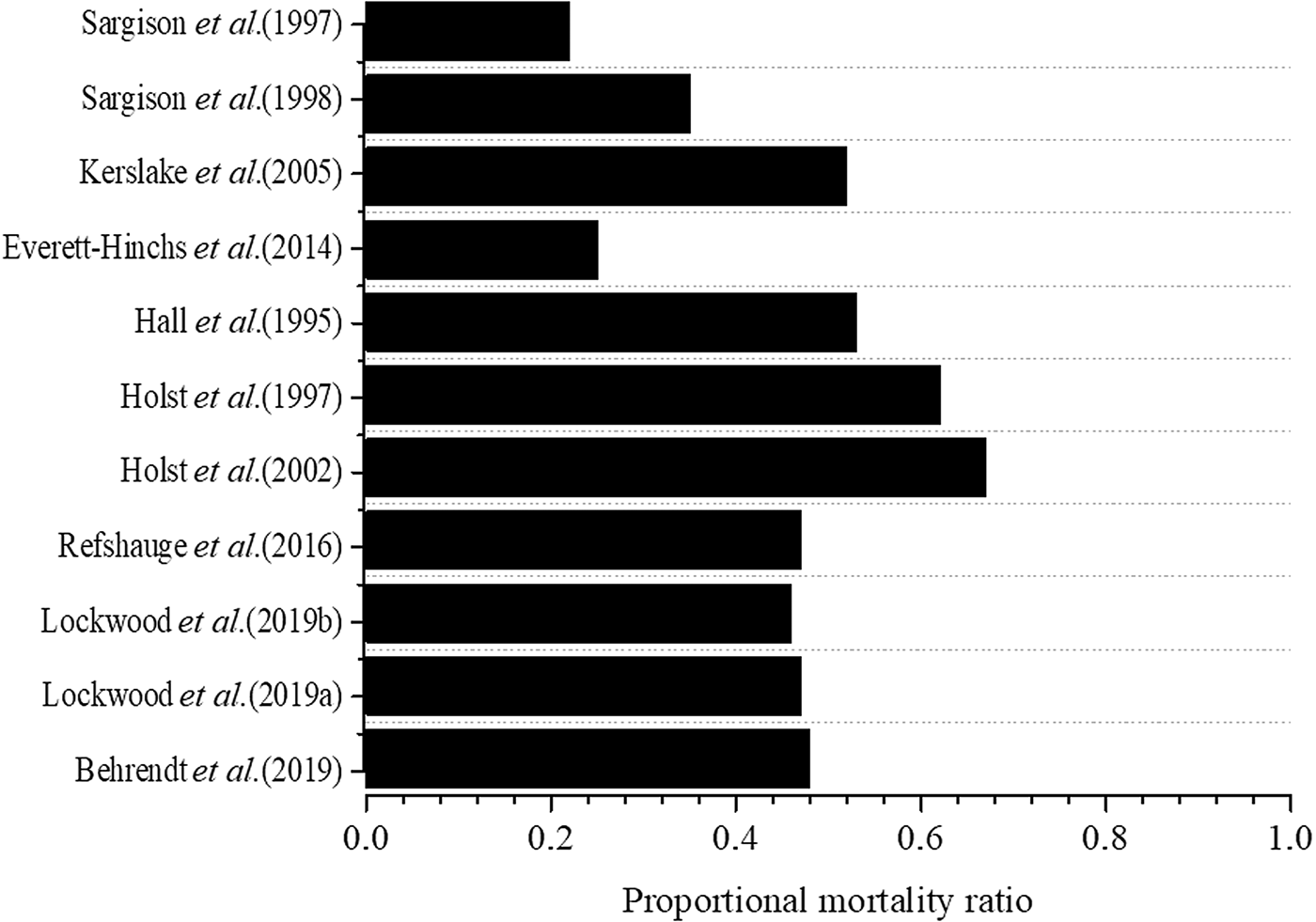
Figure 3. Proportional mortality ratio due to dystocia as a function of the number of animals born in studies with newborn lambs. Adapted from Bruce et al. (Reference Bruce, Young, Masters, Refshauge, Thompson, Kenyon and Jacobson2021).
Another contributor to neonatal mortality is the number of lambs born per birth. Refshauge et al. (Reference Refshauge, Brien, Hinch and Van De Ven2015) and Bruce et al. (Reference Bruce, Young, Masters, Refshauge, Thompson, Kenyon and Jacobson2021) Proportional mortality attributed to dystocia was higher for singles (70%) than twins (54%) or triplets (59%). Girão et al. (Reference Girão, Medeiros and Girão1998) mention mortality rates of 14% for single lambs and 30% for twin lambs. This is due to the smaller uterine space in ewes with multiple births, causing lower birth weight, in addition to the limited capacity of ewes to provide colostrum and maternal care in adequate quantity and quality to the new-born lamb (Medeiros et al., Reference Medeiros, Tabosa, Simões, Nóbrega Júnior, Vasconcelos and Riet-Correa2005). Colostrum represents a failure of a passive immune transfer. Lambs are hypogammaglobulinemia animals (i.e. dependent on the successful transfer of colostral immunoglobulins from ewe to lamb). Thus, success in feeding depends on the ability of the lamb to lift and moves to the udder, and the ewe's role is to stimulate and guide her lamb (Alexander and Williams, Reference Alexander and Williams1962; Dwyer, Reference Dwyer2008a, Reference Dwyer2008b). According to Alexander and Peterson (Reference Alexander and Peterson1961), 15% of lamb mortalities are the sole responsibility of the s sheep, 33% of lambs and 52% are the results of their behavioural association. The onset of maternal-filial behaviour after delivery is induced and characterized by olfactory and sensory cues provided by the new-born as licking, low-intensity bleating, and postures that facilitate the access of the lambs to the udder (Dwyer and Lawrence, Reference Dwyer and Lawrence2005). Negative behaviours of the sheep moments after birth can be described as not looking for a safe place for delivery, not being close to the lamb after birth, low intensity of maternal care (e.g. grooming), and may also show indifference or hostility to the new-born lamb. Factors such as maternal malnutrition during pregnancy, the presence of predators and inadequate handling (human-animal interaction, space availability, stocking rate) may be responsible for the appearance of maternal behavioural failures shortly after delivery (Robinson et al., Reference Robinson, Sinclair and McEvoy1999; Greenwood and Bell, Reference Greenwood and Bell2003). For example, reduction in the food supply in the final third of gestation resulted in lighter new-born lambs, and poor maternal care, such as less time spent cleaning the young (i.e. grooming), in addition to the appearance of aggressive targeted behaviours to the neonate (Everett-Hincks et al., Reference Everett-Hincks, Blair, Stafford, Lopez-Villalobos, Kenyon and Morris2005). The lack of maternal care right after delivery makes it difficult to ingest colostrum and the new-born's thermoregulatory capacity (i.e. the main cause of the starvation/hypothermia complex).
Effect of the thermal environment on the thermoregulation of new-born lambs
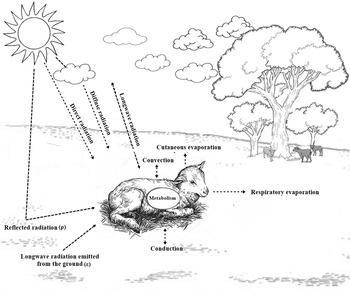
The external environment that surrounds the animal is composed of climatic elements such as air temperature, solar radiation, humidity and wind, which interact with each other, influencing its thermal balance (Silva, Reference Silva2000a). Sheep are animals that manage to maintain their body temperature in narrow levels of variation; this variation is a consequence of thermodynamic processes and the balance of generation, input and output of thermal energy. Figure 4 shows the representation of the heat balance in a new-born lamb, being described by Eqn (1):


Figure 4. Heat balance of new-born lambs. Source: personal archive.
where (α)RC is the short-wave irradiance, the amount absorbed on the surface of an animal depending on the absorption coefficient (i.e. variable depending on the radiative properties of the surface), e.g. new-born lambs in semiarid equatorial zones can be exposed to high levels of radiation and have their welfare compromised; M represents the amount of heat generated by the metabolism; e.g. in new-born lambs the main reserves in the body for heat production are brown adipose tissue (i.e. thermogenesis without tremor) and muscle glycogen (thermogenesis with tremor); Hest is the rate of storage of thermal energy; e.g. due to low body weight, new-born lambs have limited heat storage capacity; (α)RL is the amount of thermal energy exchanged between animal and environment through long wave radiation; e.g. due to the high levels of average radiant temperature found in semiarid equatorial regions, animals almost always gain heat via long wave radiation; CS is the rate of heat exchanged per convection on the animal's surface; CR is an index of heat eliminated per respiratory convection; ES represents a heat transfer by skin evaporation (can be represented by the rate of evaporation of amniotic fluid shortly after delivery and sweat); ER it is an elimination of heat through evaporation through the respiratory tract; Lastly, Δq” it is a variation of the internal thermal energy over time.
In newborn animals, the inefficiency of the thermoregulatory system after birth makes them ‘immature’ homoeotherms, losing and gaining heat more easily. This fact occurs because these animals have less thermal insulation and a higher body surface/volume ratio than adult animals. New-born lambs are born damp and have thin skin, scarce subcutaneous tissue and more superficial peripheral circulation (Nowak and Poindron, Reference Nowak and Poindron2006). In addition to the thermal gradient existing after delivery between the uterine environment and the external environment, e.g. the temperature inside the uterus with approximately 39°C for an external environment with temperatures that can reach negative values (Mercer et al., Reference Mercer, Andrews and Szekely1979).
In regions with a semiarid equatorial environment, high levels of air temperature, solar radiation, and average radiant temperature can be challenging for new-borns in the first hours of life. These animals can absorb an excessive amount of heat via thermal radiation, especially when the farrowing occurs in a pasture environment and/or in management centres without providing shade (Neves et al., Reference Neves, de Azevedo, da Costa, Guim, Leite and Chagas2009). On the other hand, even though it is an inter-tropical region, cold nights (daily thermal amplitude of ± 15°C) can favour a rapid transfer of sensitive heat to the environment and may even be greater than the amount of metabolic heat produced by the animal (Fonsêca et al., Reference Fonsêca, Maia, Saraiva, de Melo Costa, da Silva, Abdoun and Fuller2019). In addition, wind and rain can make the heat balance more expensive, by recruiting optional thermogenesis to replace the excess heat lost, if the animal does not have the opportunity to find shelter (DeShazer et al., Reference DeShazer, Hahn and Xin2009).
To maintain thermal balance after birth, the new-born needs ‘fuel’ to increase its metabolic rate, in an attempt to match the inflows and outflows of thermal energy, especially when born at lower temperatures. Studies with several species of mammals have shown that new-born animals increase their metabolism to a maximum rate (i.e. summit metabolism), such as piglet (Mount and Rowell, Reference Mount and Rowell1960; Noblet and Le Dividich, Reference Noblet and Le Dividich1981); rats (Taylor, Reference Tayloy1970; Mortolla and Dotta, Reference Mortolla and Dotta1992); rabbits (Girard, Reference Girard1990); calves (Okamoto et al., Reference Okamoto, Robinson, Christopherson and Young1986; Steinhoff, Reference Steinhoff2010); goats (Celi et al., Reference Celi, Di Trana and Quaranta2008; Páleníková et al., Reference Páleníková, Hauptmanová, Pitropovská, Páleník, Husáková, Pechová and Pavlata2014); and lambs (Alexander, 1962, Reference Alexander1970; Eales and Small, Reference Eales and Small1980; Slee et al., Reference Slee, Simpson and Woolliams1987; Plush et al., Reference Plush, Brien, Hebart and Hynd2016). The main energetic substrates for the metabolism of new-borns are brown adipose tissue, muscle glycogen and colostrum intake (Mercer et al., Reference Mercer, Andrews and Szekely1979). Lambs with lower birth weights reach their maximum metabolic rate at higher temperatures when compared to lambs born with higher weights. This greater susceptibility in light animals occurs due to the lower supply of energy fuels.
Eales and Small (Reference Eales and Small1980) observed that new-born lambs increased their metabolic rate and rectal temperature when subjected to cold conditions, however, as soon as the metabolism reaches its maximum production, the rectal temperature drops slowly (Fig. 5). In addition, the metabolic rate peak occurred between 60 and 100 min; in this interval, if the new-born does not obtain energy from the ingestion of colostrum, the body reserves are depleted, which can lead to the death of the new-born by starvation/hypothermia complex (Hamadeh et al., Reference Hamadeh, Hatfield, Kott, Sowell, Robinson and Roth2000).
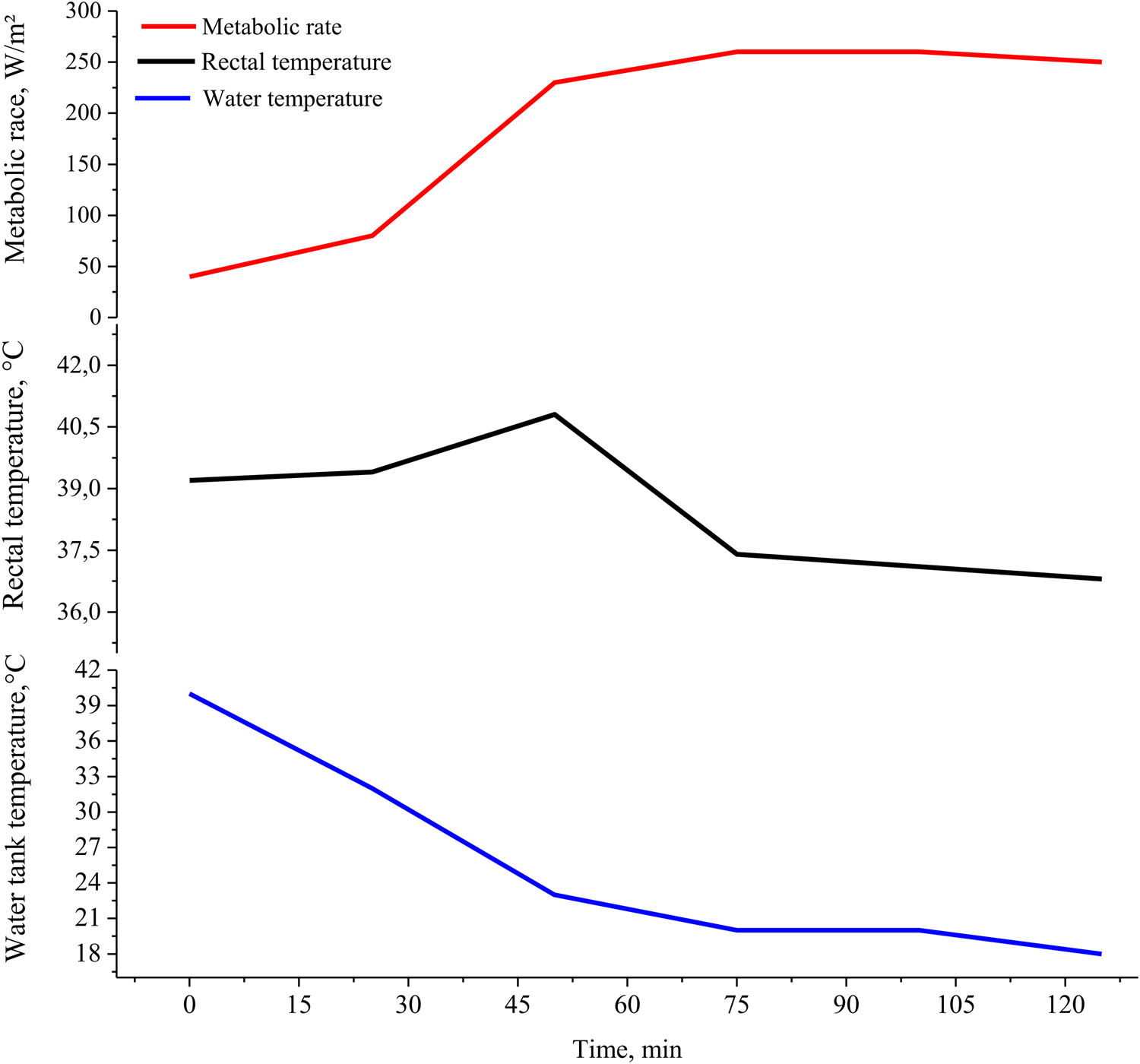
Figure 5. Metabolic rate, rectal temperature, and water tank temperature when measuring basal metabolic rate and maximum metabolic rate in a 4-hour-old lamb with 4.60 kg of body weight (Adapted de Eales and Small, Reference Eales and Small1980).
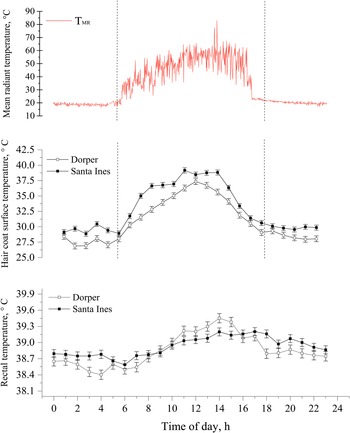
Many studies have been conducted to verify the effects of the thermal environment on the thermoregulation of new-born lambs (McCoard et al., Reference McCoard, Henderson, Knol, Dowling and Webster2014; El Hadi et al., Reference El Hadi, Frascati, Granzotto, Silvestrin, Ferlini, Vettor and Rossato2016; Labeur et al., Reference Labeur, Villiers, Small, Hinch and Schmoelzl2017; Santos, Reference Santos2020). Most of these studies were carried out in a temperate environment. McCoard et al. (Reference McCoard, Henderson, Knol, Dowling and Webster2014) state that in conditions of a thermoneutral environment or mild cold stress, an adequate balance of internal heat generation, through the production of thermal energy from brown adipose tissue and heat loss can prevent hypothermia after birth. On the other hand, in more extreme cold conditions, thermogenesis with tremors using muscle glycogen reserves can be essential for maintaining the thermal balance of new-born lambs (Plush et al., Reference Plush, Brien, Hebart and Hynd2016). Hypothermic lambs have greater lethargy, tend not to follow their mother, and may show signs of distress, compromising their survival (Defra, 2004). In our studies with Dorper, Santa Ines and Morada Nova lambs in the semi-arid region, we observed that newborn lambs in colder seasons (Ta < 25) took longer to carry out the first feeding (Fig. 6; Fonsêca et al., Reference Fonsêca, Saraiva, Pimenta Filho, Furtado, Mariz, Silva and Pinheiro2014; Santos et al., in preparation). Dwyer and Morgan (Reference Dwyer and Morgan2006) observed that lambs with a longer time to perform the first suckle had a lower rectal temperature at 1 h and 24 h after birth. Santos et al. (in preparation) observed that coat surface temperature and rectal temperature correlated positively with the mean radiant temperature pattern (Fig. 7). In addition, dark-coated lambs tend to have higher coat surface temperatures, which can generate greater absorption of radiant thermal load.

Figure 6. Mean of latency to suckle (min) both in studies in the semi-arid region with lambs in the semi-arid region as a function of air temperature classes. (Adapted Fonsêca et al., Reference Fonsêca, Saraiva, Pimenta Filho, Furtado, Mariz, Silva and Pinheiro2014 and personal archive).
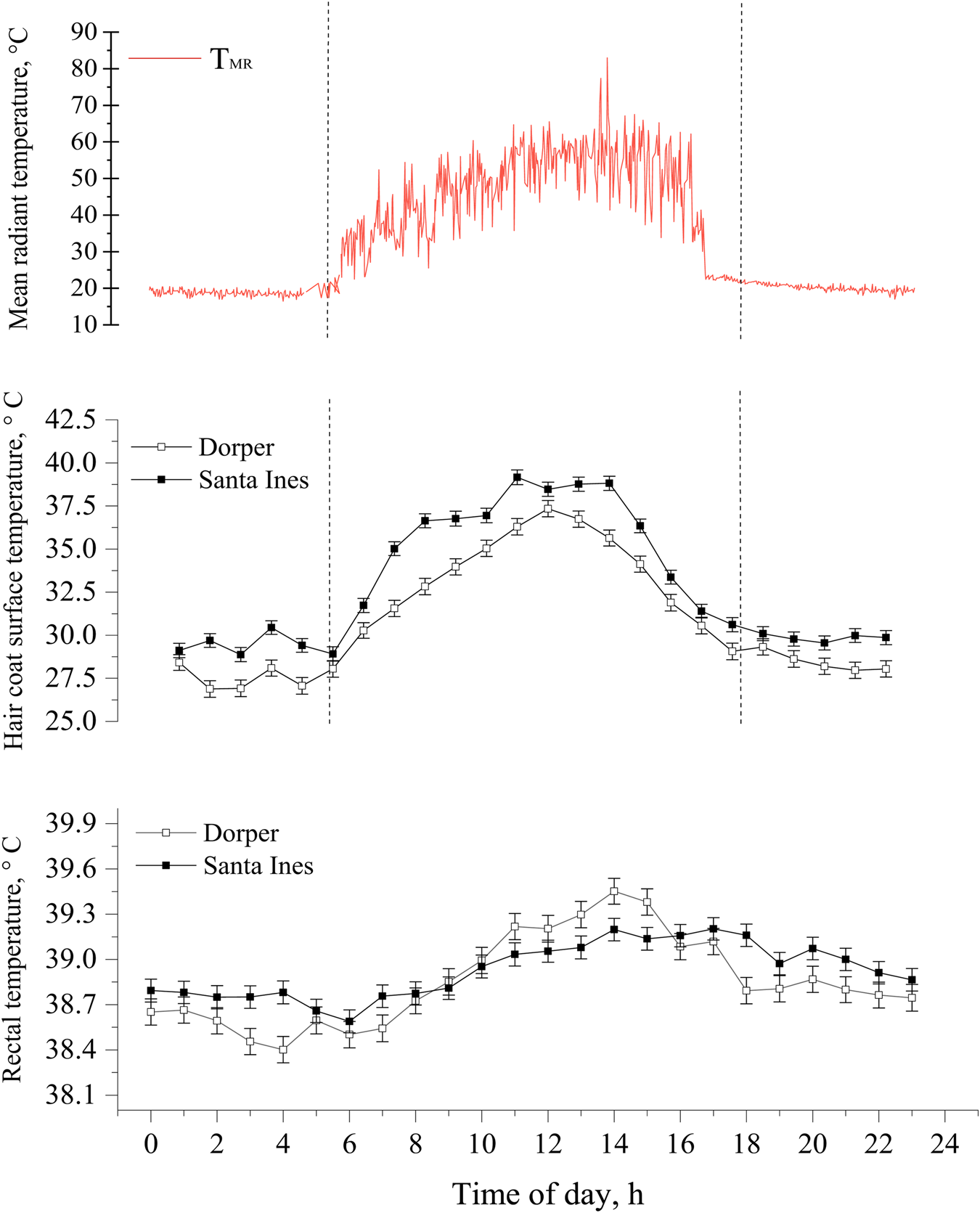
Figure 7. Least square means of mean radiant temperature (°C), hair coat surface temperature (°C) and rectal temperature (°C) both for Dorper and Santa Inês lambs across the 24 h. The data are presented as mean ± SEM. Source: personal archive.
Research has also shown the ability of new-born lambs to maintain body temperature within narrow ranges under conditions of maximum fluctuation in air temperature. For example, the body temperature levels of new-borns up to 24 h old of the Dorper and Santa Ines breeds fluctuated from 36°C to 41°C under temperature conditions ranging from 20.7 to 36°C (Santos et al., in preparation). Likewise, Riesenfeld et al. (Reference Riesenfeld, Hammarlund, Jonzon and Sedin1988) in a study with Merino lambs observed that rectal temperature fluctuated from 38 to 40°C under temperature conditions ranging from 25 to 37°C. However, these authors mentioned that they allow normothermia to be achieved at the cost of excessively increasing the rate of respiratory and cutaneous evaporation, refuting a theory of ‘immature thermoregulators’ for new-born lambs. Faurie et al. (Reference Faurie, Mitchell and Laburn2004) found that the neonatal ability to maintain a low body temperature, even during the hottest hours of the day, is available with the ability to sweat and pant a few hours after birth.
Under conditions of high radiant thermal load, a large part of the studies was carried out on adult animals, and there is still a lack of data to understand the thermoregulation of lambs and the factors that affect the heat balance of animals raised in these conditions. Silva et al. (Reference Silva, Guilhermino and Morais2010) tried to quantify the thermal radiation absorbed by the surface in dairy cows raised on pasture in semiarid. Maia et al. (Reference Maia, da Silva, Nascimento, Nascimento, Pedroza and Domingos2015) investigated the effect of coat colour on goats exposed or not to solar radiation; and noted that black or white goats need protection from solar radiation to maintain thermal balance. In addition, when managed without this protection from the sun's rays, their physiological responses are altered, especially in black goats, when the solar irradiance is greater than 800 W/m2. Likewise, Fonsêca et al. (Reference Fonsêca, Maia, Saraiva, de Melo Costa, da Silva, Abdoun and Fuller2019) evaluated the effect of the radiant thermal environment on the bio-thermal responses of Morada Nova ewes and observed that the minimum body temperature for 24 h was correlated with the minimum temperature of the environment, indicating that heat conservation strategies were probably important for Morada Nova ewes when the rates of sensitive heat loss exceed the heat generated by metabolism. Santos et al. (in preparation) in studies with lambs in a semi-arid region, it was observed that the thermal radiation environment in an equatorial semi-arid region is a challenge for the thermoregulation of new-born lambs. The authors observed that the vigour and surface temperature of the coat were directly influenced by the radiant thermal load, which could even generate hyperthermia when exposed to solar radiation.
Conclusion and prospects
Information in the literature on the main survival risks of new-born lambs is abundant; studies show that the world average of neonatal-perinatal mortality reaches 15%. Mortality rates are often attributed to various causes such as birth weight, dystocia, adverse weather conditions, inadequate milk or colostrum supply, sibling competition in multiple births, hunger/hypothermia complex, and poor ewe or lamb behaviour. Most of the studies that investigate the factors that influence the thermal balance of new-born lambs were developed in laboratory conditions and temperate climate regions, which almost always emphasized the effect of cold as the greatest challenge to homoeothermy of new-borns in the first hours after birth. Research on thermoregulation and factors that affect the heat balance of new-born lambs in equatorial semi-arid regions is scarce. However, some studies already seek to understand how these animals deal with conditions of high mean radiant temperatures.
Acknowledgements
We are grateful to Empresa Paraibana de Pesquisa, Extensão Rural e Regularização Fundiária (EMPAER) for the infrastructure of the facilities for experimenting. The authors are grateful for the support of the Study Group on Bioclimatology, Behaviour and Animal Welfare (BIOET) at the Federal University of Paraíba, Brazil. In addition, we are grateful for the financial support (grants) granted by the National Council for Scientific and Technological Development (CNPq) and the Coordination for the Improvement of Higher Education Personnel (CAPES), Brazil.
Author contributions
José Danrley Cavalcante dos Santos: Contributed substantially to the conception, and design of the article and to the interpretation of the pertinent literature. She wrote the article.
Edilson Paes Saraiva: Contributed substantially to the conception and design of the article. Oversaw the writing process. Critically reviewed the article for important intellectual content.
Edgard Cavalcanti Pimenta Filho: Critically reviewed the article for important intellectual content.
Geni Caetano Xavier Neta: Contributed substantially to data collection and writing.
Larissa Kellen da Cunha Morais: Contributed substantially to data collection.
Humberto da Silva Teti: Contributed substantially to data collection.
Sérgio da Silva Fidelis: Contributed substantially to data collection.
Financial support
The authors are grateful to the National Council for Scientific and Technological Development (CNPq).
Conflict of interest
The authors declare there are no conflicts of interest.