



No CrossRef data available.
Published online by Cambridge University Press: 17 October 2024
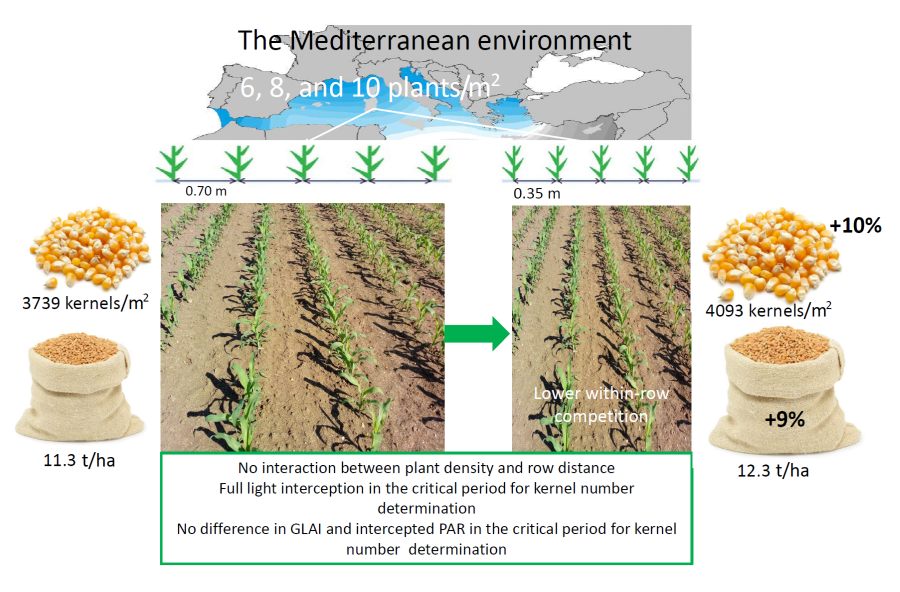
The spatial distribution of maize plants influences the number of kernels set per plant by modifying the radiation environment during the critical period for kernel number determination. A two-year field experiment was conducted in Sardinia to explore whether, in a Mediterranean environment, a reduction in row distance can have a positive effect on grain yield of full-season maize hybrids and, if it does, whether this effect is also affected by plant density. Treatments were a factorial combination of three plant densities (6, 8 and 10 plants/m2) and two row spacings (35 and 70 cm). Grain yield and grain number benefited from a reduction in row distance from the canonical 70 cm to 35 cm irrespective of plant density and yield level (14.7 t/ha on average in 2021, 9.9 t/ha in 2022 vs 13.7 and 8.8 t/ha with canonical row distance). Row distance did not affect either the coefficient of extinction of radiation or the intercepted photosynthetically active radiation (PAR) during the critical period for grain number determination. The higher grain number per unit area at the smaller row distance translated into a greater grain yield, which cannot solely be attributed to radiation interception and plant growth rate during the period of kernel number determination. Halving the common row distance is a valuable management option for full-season hybrids grown in Mediterranean environments without nitrogen and water limitations as it seems capable of increasing grain yield via an increase in kernel number per unit area, regardless of plant density and yield level.


To send this article to your Kindle, first ensure no-reply@cambridge.org is added to your Approved Personal Document E-mail List under your Personal Document Settings on the Manage Your Content and Devices page of your Amazon account. Then enter the ‘name’ part of your Kindle email address below. Find out more about sending to your Kindle. Find out more about saving to your Kindle.
Note you can select to save to either the @free.kindle.com or @kindle.com variations. ‘@free.kindle.com’ emails are free but can only be saved to your device when it is connected to wi-fi. ‘@kindle.com’ emails can be delivered even when you are not connected to wi-fi, but note that service fees apply.
Find out more about the Kindle Personal Document Service.
To save this article to your Dropbox account, please select one or more formats and confirm that you agree to abide by our usage policies. If this is the first time you used this feature, you will be asked to authorise Cambridge Core to connect with your Dropbox account. Find out more about saving content to Dropbox.
To save this article to your Google Drive account, please select one or more formats and confirm that you agree to abide by our usage policies. If this is the first time you used this feature, you will be asked to authorise Cambridge Core to connect with your Google Drive account. Find out more about saving content to Google Drive.