



Scientific Classification
Domain: Eukaryota
Kingdom: Plantae
Phylum: Spermatophyta
Subphylum: Angiospermae
Class: Dicotyledonae
Order: Solanales
Family: Solanaceae
Subfamily: Solanoideae
Tribe: Lycieae
Genus: Lycium
Species: ferocissimum Miers (Miers Reference Miers1854)
Synonyms: Lycium campanulatum E.Mey. ex C.H.Wright and Lycium macrocalyx Domin.,
Lycium afrum L. (misapplied), Lycium europaeum L. (misapplied), and Lycium horridum Thunb. (misapplied)
EPPO Code: LYUFE
Names and Taxonomy
The name of the genus Lycium originates from the Greek name lykion for a thorny shrub, derived from Lycia, the name of an ancient country in Asia Minor where a similar spiny shrub was found. The specific epithet ferocissimum comes from the Latin ferox, meaning “bold” or “fearless,” referring to the very spiny nature of the shrub (Green Reference Green, Orchard and Wilson1994; Parsons and Cuthbertson Reference Parsons and Cuthbertson2001).
Common names for Lycium ferocissimum are African boxthorn, boxthorn, snake-berry (slangbessie in Afrikaans). “Boxthorn” is believed to be derived from the Dutch boksdorn, the name Dutch settlers to South Africa gave the plant (Parsons and Cuthbertson Reference Parsons and Cuthbertson2001).

Lycium ferocissimum belongs to the genus Lycium, tribe Lycieae, within the subfamily Solanoideae of the family Solanaceae (Levin et al. Reference Levin, Bernardello, Whiting and Miller2011; Särkinen et al. Reference Särkinen, Bohs, Olmstead and Knapp2013; Stevens Reference Stevens2019). Members of Lycium tend to be woody shrubs with short lateral branches, usually ending in a spine, and are deciduous or evergreen (Webb et al. Reference Webb, Sykes and Garnock-Jones1988). Lycium species typically inhabit arid to semiarid environments (Adamson Reference Adamson1938; Levin et al. Reference Levin, Shak, Miller, Bernardello and Venter2007). The genus includes approximately 90 species, widely distributed, with centers of diversity in southern South America, southwestern North America, and southern Africa (Levin et al. Reference Levin, Bernardello, Whiting and Miller2011; Stevens Reference Stevens2019). It is considered likely that Lycium arose in the Americas (probably South America) and then dispersed to Africa, Eurasia, and elsewhere, including a single species (Lycium australe F. Muell.) to Australia (Fukuda et al. Reference Fukuda, Yokoyama and Ohashi2001; Levin et al. Reference Levin, Shak, Miller, Bernardello and Venter2007). The taxonomy and phylogeny of Lycium is highly complex, with evidence of hybridization and many undescribed species within numerous species complexes now emerging in South African populations (Levin et al. Reference Levin, Shak, Miller, Bernardello and Venter2007; McCulloch et al. Reference McCulloch, Mauda, Chari, Martin, Gurdasani, Morin, Walter and Raghu2020; Venter Reference Venter2000). Despite this, recent genetic analyses found no evidence of hybridization with any other Lycium species in Australia (McCulloch et al. Reference McCulloch, Mauda, Chari, Martin, Gurdasani, Morin, Walter and Raghu2020). The fleshy, bird-dispersed fruits of Lycium species most likely contributed to the cosmopolitan distribution of the genus (Fukuda et al. Reference Fukuda, Yokoyama and Ohashi2001; Levin et al. Reference Levin, Bernardello, Whiting and Miller2011). Lycium ferocissimum is a species native to southern Africa and part of the “Old World” clade within Lycium (Levin et al. Reference Levin, Shak, Miller, Bernardello and Venter2007). Close relatives of L. ferocissimum in this clade, especially the Eurasian goji berry species Lycium barbarum L. and Lycium chinense Mill., are also recognized as invasive plants (Randall Reference Randall2017).
Description
Lycium ferocissimum is a densely branched perennial shrub that can grow up to 5-m high (but more often 2 to 3 m) and up to 5-m across (though most commonly up to 3 m) (McCulloch et al. Reference McCulloch, Mauda, Chari, Martin, Gurdasani, Morin, Walter and Raghu2020; Parsons and Cuthbertson Reference Parsons and Cuthbertson2001) (Figure 1A). However, in wind-prone situations such as coastal sites, its habit is often quite different. There it is wind-pruned, very dense, and often relatively short, with its shape determined by the predominant wind direction (Noble and Rose Reference Noble and Rose2013; Taylor and Tennyson Reference Taylor and Tennyson1999) (Figure 1B).

Figure 1. Growth form of Lycium ferocissimum (A) in pastures and (B) on the South Australian coastline, where the species is wind-pruned by prevailing winds. (Photo credits: (A) New South Wales National Parks and Wildlife Service; (B) Matt Rose, Natural State.)
Stems are silver-gray when young, turning light brown to gray as they mature, and becoming fissured with age. The main stems grow large spines (to 15 cm), with smaller spines on branches. Leaves of L. ferocissimum (to 40-mm long and 20-mm wide) have very short petioles; are slightly fleshy, simple and entire, ovate, obovate to elliptic in shape, and glabrous; and cluster at nodes (Figure 2). Inflorescences are solitary or in pairs and form in the leaf axil (Figure 2). They are 8 to 12 mm in diameter and 10- to 12-mm long. Petals are white to lilac with purple markings toward the inside (Figure 2). The pedicel is 5 to 16 mm, there is a calyx with five unequal sepals, there are five petals, and there are 5 to 6 exerted stamens. Flowers (Figure 2) are present most of the year but are most prolific during summer. The fruit is a smooth round berry, initially green, but ripening to orange-red, up to 12 mm in diameter and with a prominent calyx, and typically containing between 20 and 70 dull yellow seeds (Blood Reference Blood2001; Green Reference Green, Orchard and Wilson1994; McCulloch et al. Reference McCulloch, Mauda, Chari, Martin, Gurdasani, Morin, Walter and Raghu2020; Muyt Reference Muyt2001; Parsons and Cuthbertson Reference Parsons and Cuthbertson2001; Purdie et al. Reference Purdie, Symon, Haegi and George1982) (Figure 2B).

Figure 2. Lycium ferocissimum flowers (A and B) and fruit (B). (Photo credit: Evans Mauda.)
Economic Importance and Environmental Impacts
Detrimental
Lycium ferocissimum is an invasive weed of environmental and agricultural ecosystems. It is considered a Weed of National Significance in Australia (Australian Weeds Committee 2013), and both environmental and agricultural stakeholders consider it to be difficult and costly to control (Cousens et al. Reference Cousens, Kennedy, Maguire and Williams2013; Ireland et al. Reference Ireland, Rafter, Kumaran, Raghu and Morin2019b) (Table 1). By forming impenetrable spiny thickets, the plant impedes the habitat needs of wildlife and movement of livestock, outcompetes native vegetation, and can harbor nonnative pest species such as rabbits (Oryctolagus cuniculus L.), rats, and fruit flies (Noble and Adair Reference Noble and Adair2014; Noble et al. Reference Noble, Adair and Yen2014; Noble and Rose Reference Noble and Rose2013).
Table 1. Major impacts of Lycium ferocissimum in Australia, as perceived by stakeholders (Ireland et al. Reference Ireland, Rafter, Kumaran, Raghu and Morin2019b).
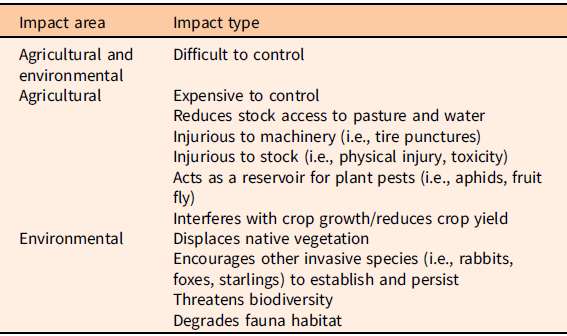
In Australia, displacement of native vegetation and the encouragement of establishment and persistence of other invasive pest species such as rabbits and rats have been identified as the main environmental impacts, while damage to machinery and prevention of stock access to water have been identified as the main agricultural impacts (Ireland et al. Reference Ireland, Rafter, Kumaran, Raghu and Morin2019b).
In an environmental context, L. ferocissimum is known to displace native vegetation and degrade faunal habitat, directly impact native fauna (e.g., by ensnaring birds), and likely facilitates the degradation of cultural heritage sites (Noble and Adair Reference Noble and Adair2014; Noble et al. Reference Noble, Adair and Yen2014; Noble and Rose Reference Noble and Rose2013). Lycium ferocissimum adversely impacts a significant range of vegetation species and communities. It is considered a threat to biodiversity in at least two rangeland biodiversity hot spots in Australia (Martin et al. Reference Martin, Campbell and Grounds2006) and is invasive in a number of global biodiversity hot spots, including southwest Australia, the Mediterranean Basin, and New Zealand (GBIF 2020; Mittermeier et al. Reference Mittermeier, Turner, Larsen, Brooks, Gascon, Zachos and Habel2011). In some coastal areas of Australia and New Zealand, L. ferocissimum has become the only woody plant present, changing the vegetation structure in some small island and coastal dune environments (Lavers Reference Lavers2014; Timmins and Mackenzie Reference Timmins and Mackenzie1995; Webb et al. Reference Webb, Sykes and Garnock-Jones1988; Ziegler and Hopkins Reference Ziegler and Hopkins2011). In salt marshes where L. ferocissimum has become established in Australia, bird species assemblages and behavior appear altered by the presence of the weed (Carlos et al. Reference Carlos, Weston and Gibson2017). This reduces habitat suitability for some native species and makes these areas more hospitable for pest animals such as starlings (Sturnus vulgaris L.) or more aggressive raptor birds, which may make use of the increased roosting sites afforded by L. ferocissimum to prey on other native and exotic fauna.
Lycium ferocissimum can significantly alter and interfere with native fauna habitats. Several native Australian animals have been recorded as being adversely impacted by the presence of L. ferocissimum. The fine, dense root mass of L. ferocissimum has been observed to impede the burrowing efforts of short-tailed shearwaters (Ardenna tenuirostris Temminck) (Lawley et al. Reference Lawley, Lawley and Page2005). Thorns of L. ferocissimum can ensnare, injure, and kill coastal native birds, including the short-tailed shearwater, New Zealand fairy prions (Pachypptilla turtur Kuhl), and white-faced storm petrels (Pelagodroma marina Latham) (Lohr and Keighery Reference Lohr and Keighery2016; Noble and Rose Reference Noble and Rose2013; Phillips Reference Phillips2014; Taylor Reference Taylor1968; Ziegler and Hopkins Reference Ziegler and Hopkins2011). On islands off South and Western Australia, L. ferocissimum disrupts seal (Arctocephalus spp.) and sea lion (Neophoca cinerea Péron) breeding, by displacing the native shrub Nitraria billardieri DC., which is used by seals for sheltering pups (Hussey et al. Reference Hussey, Keighery, Dodd, Lloyd and Cousens2007; Western Australian Herbarium 1998–). Lycium ferocissimum does not provide the equivalent quality of nursery habitat, leaving pups more vulnerable to predation (Humphries et al. Reference Humphries, Groves, Mitchell, Kikkawa and Moritz1994).
Cultural heritage site degradation may be facilitated by the presence of L. ferocissimum. Coastal areas in Australia, where L. ferocissimum is known to occur (Cousens et al. Reference Cousens, Kennedy, Maguire and Williams2013; Erkelenz Reference Erkelenz1994; Lohr and Keighery Reference Lohr and Keighery2016), are also likely locations for indigenous heritage such as middens, artifacts, and other evidence of occupation (Cann et al. Reference Cann, De Deckker and Murray-Wallace1991; Veth et al. Reference Veth, Ward, Manne, Ulm, Ditchfield, Dortch, Hook, Petchey, Hogg, Questiaux, Demuro, Arnold, Spooner, Levchenko and Skippington2017). These sites may be degraded through increased rabbit and fox (Vulpes vulpes L.) burrowing associated with L. ferocissimum, which may cause direct or indirect damage to artifacts and historical sites (Noble and Adair Reference Noble and Adair2014; Noble et al. Reference Noble, Adair and Yen2014; Noble and Rose Reference Noble and Rose2013).
In an agricultural context, L. ferocissimum has been associated with reducing access to pasture and water in grazing systems (Brown Reference Brown1969; Lee Reference Lee1978), harboring pest animals such as rabbits, foxes, and pest birds, and puncturing tires and injuring livestock and people with its thorns (Hoskin Reference Hoskin2006; Noble and Adair Reference Noble and Adair2014). It also hosts key insect pests and diseases of concern to agriculture, including the Queensland fruit fly Bactrocera tryoni Froggatt (Plant Health Australia 2017) and tomato-potato psyllid Bactericera cockerelli Sulc (Vereijssen et al. Reference Vereijssen, Smith and Weintraub2018). Bactericera cockerelli is a North American psyllid species that, in its place of origin, overwinters on several native and nonnative Lycium species (Cooper et al. Reference Cooper, Horton, Miliczky, Wohleb and Waters2019; Thinakaran et al. Reference Thinakaran, Horton, Rodney Cooper, Jensen, Wohleb, Dahan, Mustafa, Karasev and Munyaneza2017). It is a major pest of solanaceous crops, as it is the vector of the plant bacterium Candidatus Liberibacter solanacearum Jagoueix et al. (CLso), the causal agent of zebra chip disease in potatoes (Solanum tuberosum L.) (Liefting et al. Reference Liefting, Weir, Pennycook and Clover2009; Munyaneza Reference Munyaneza2012). Populations of B. cockerelli may build up or survive on overwintering hosts such as L. ferocissimum and then colonize commercial crops in spring (Vereijssen et al. Reference Vereijssen, Smith and Weintraub2018), so the presence of L. ferocissimum in agricultural areas is likely to be of significant concern when managing populations of the psyllid.
Beneficial
Lycium ferocissimum may also provide significant social amenity and refuge benefits to native flora and fauna, particularly where native habitat has been removed or degraded. It was considered a valued hedge and windbreak in southern Africa (Venter Reference Venter2000), Australia (Herbiguide 2015; Parsons and Cuthbertson Reference Parsons and Cuthbertson2001), and New Zealand (Lambert Reference Lambert2015) during the 1800s and early 1900s. It is also known to provide refuge for more palatable plant species in heavily grazed situations where those species might otherwise be lost (Rutherford et al. Reference Rutherford, Powrie and Husted2014). For example, on the Eyre Peninsula in South Australia, L. ferocissimum may protect threatened west-coast mint bush (Prostanthera calycina F.Muell. ex Benth.) juveniles from grazing (Pobke Reference Pobke2007). In Africa, L. ferocissimum is recognized for playing a similar role as a component of native vegetation (Todd Reference Todd2000). Within its native range in South Africa, L. ferocissimum is recognized for its potential in restoring degraded landscapes. Its hardiness, thorns, and limited palatability mean it has the capacity to withstand herbivore-driven degradation (Rutherford et al. Reference Rutherford, Powrie and Husted2014). There, it is considered also to have potential as a keystone restoration species, due to its regular and large crops of fruits, which are attractive to birds that might then disperse other native plant species (van der Vyver et al. Reference van der Vyver, Cowling, Campbell and Difford2012).
In regard to fauna, L. ferocissimum is considered to confer significant benefits to a range of vulnerable Australian species, though these interactions are complex. For example, the critically endangered orange-bellied parrot’s (Neophema chrysogaster Latham) habitat in coastal Victoria and South Australia now includes L. ferocissimum thickets. However, where the parrot’s habitat is intact, L. ferocissimum and other invasive species are a threat to the parrot’s foraging habitat (Orange-bellied Parrot Recovery Team 2006). In some coastal locations, little penguins (Eudyptula minor J.R. Forster) and yellow-eyed penguins (Megadyptes antipodes Hombron & Jacquinot) may utilize the shelter of thorny L. ferocissimum shrubs for nesting and protection from predators (Lalas et al. Reference Lalas, Jones and Jones1999; Noble and Rose Reference Noble and Rose2013). Positive correlations have been documented between wombat (Vombatus ursinus Shaw) burrows and L. ferocissimum in Australia (Roger et al. Reference Roger, Laffan and Ramp2007; Taylor Reference Taylor1993). However, this beneficial role of L. ferocissimum for wombats is likely related to the species replacing the structural role played by native shrubs, which may have been displaced by L. ferocissimum or other edaphic or anthropogenic causes. The nationally endangered southern brown bandicoot (Isoodon obesulus Shaw) and nationally vulnerable eastern barred bandicoot (Perameles gunnii Gray) in Australia are both known to consume L. ferocissimum fruit (Heinsohn Reference Heinsohn1964; Moloney Reference Moloney1982; Quin Reference Quin1985). These species may also gain predator shelter benefits from L. ferocissimum, particularly where native alternatives are not available. The nationally vulnerable Australian native greater stick-nest rat (Leporillus conditor Sturt) is reported to use L. ferocissimum on islands off South Australia for shelter from its primary predator, barn owls (Tyto delicatula Gould). The rats have been observed feeding on L. ferocissimum foliage, and when the rodent numbers are exceptionally high, consumption of bark has also been evident. The greater stick-nest rat has also been observed using L. ferocissimum segments as building material to construct its stick nests (J van Weenen, South Australian Government, personal communication, 2013). On islands in the Bass Strait of Australia, Tasmanian pademelons (Thylogale billardierii Desmarest) have been recorded as being common in L. ferocissimum bushes, having climbed to various heights up to 3 m (Figure 3). Pademelons were found to have established routes up through the bushes, allowing them access to browse leaves and fruit (Driessen Reference Driessen2011). Lycium ferocissimum has also been recorded as providing some cover for Tasmanian devils in Australia (Sarcophilus harrisii Boitard) (Guiler Reference Guiler1970). While a range of both common and endangered native birds and mammals make use of L. ferocissimum for supplemental habitat and food in its invaded range (Carlos et al. Reference Carlos, Weston and Gibson2017; Noble and Rose Reference Noble and Rose2013; Zann Reference Zann1994), these relationships are complex, and the overall nature of the impact of L. ferocissimum presence on biodiversity in these regions is unclear.

Figure 3. Tasmanian pademelons on Inner (West) Sister Island climb to 3 m to feed on Lycium ferocissimum leaves and fruit. (Photo credit: Michael Driessen.)
Geographic Distribution
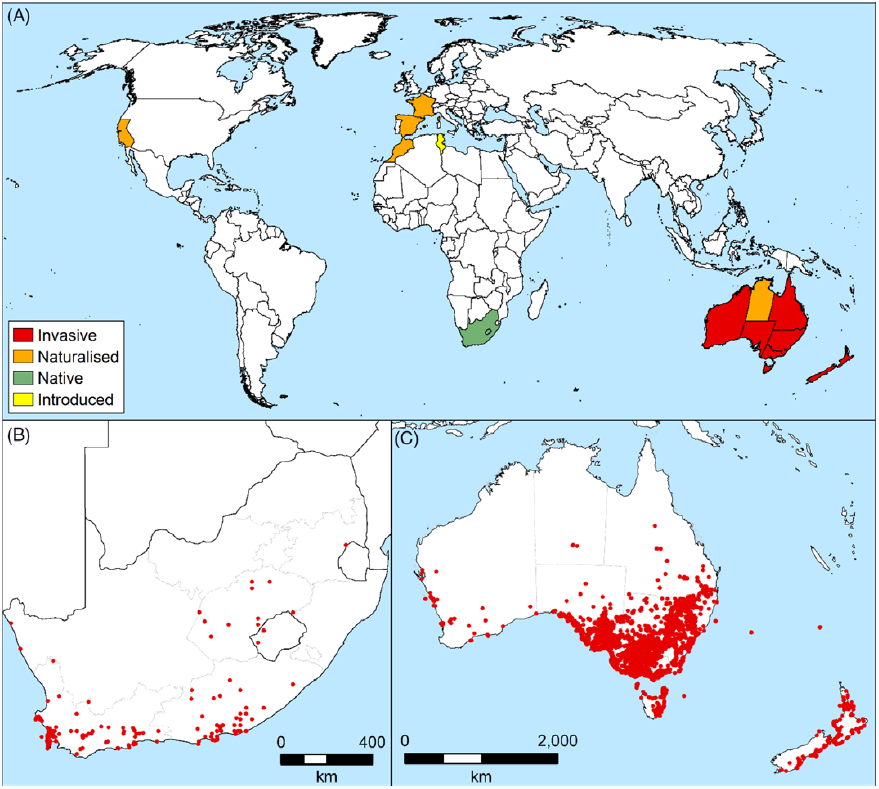
Lycium ferocissimum is native to South Africa, with a nonnative global distribution that is spatially limited, but intensive where it occurs, particularly in Australia and New Zealand. The plant is naturalized and invasive in parts of Europe and has limited naturalized presence in the United States. (Figure 4A).

In the plant’s native range of South Africa, it is widespread and native to the Eastern and Western Cape provinces (Venter Reference Venter2000). Further afield from its native range within South Africa there are distribution records for populations in the Free State, Kwazulu-Natal, Mpumalanga, and Northern Cape provinces of South Africa, and northern Lesotho, most likely occurring due to the species being planted as a hedge (GBIF 2020; Venter Reference Venter2000; Welman Reference Welman1993; Welman Reference Welman2003) (Figure 4B). While L. ferocissimum has been recorded as occurring in Namibia (GBIF 2018), the veracity of these records is questionable, especially given the age of the records (1963 and 1976) and the complexities of L. ferocissimum and Lycium spp. identification and taxonomy (Levin et al. Reference Levin, Shak, Miller, Bernardello and Venter2007; McCulloch et al. Reference McCulloch, Mauda, Chari, Martin, Gurdasani, Morin, Walter and Raghu2020; Venter Reference Venter2000).
Lycium ferocissimum is recorded as being present in a number of northern African and European nations within the Mediterranean Basin. It is recorded as introduced to Tunisia (Monastir, Nabeul, and Siliana governorates) (GBIF 2020; Venter Reference Venter2000) and introduced and naturalized in coastal Morocco (Gharb-Chrarda-Béni Hssen, Marrakech-Tensift-Al Haouz, and Rabat-Salé-Zemmour-Zaer regions) (GBIF 2020; Lambinon and Lewalle Reference Lambinon and Lewalle1986; Venter Reference Venter2000), coastal southwestern Spain (Málaga Province, plus a single introduced site in the city of Guadalajara in central Spain) (GBIF 2020; Perez Latorre et al. Reference Perez Latorre, Yus Ramos and Dana Sanchez2006), and Cyprus (recorded from shores of Lake Akrotiri in the south, to abandoned mine sites in the northeast) (Cetinkaya and Sozen Reference Cetinkaya and Sozen2011; Gallego Reference Gallego2012; Hand Reference Hand2000; İlseven and Baştaş Reference İlseven and Baştaş2018; Meikle Reference Meikle1985; Perez Latorre et al. Reference Perez Latorre, Yus Ramos and Dana Sanchez2006; Peyton and Mountford Reference Peyton and Mountford2015; Venter Reference Venter2000). The most recent European records of the species, in the southern provinces of Cagliari and Medio Campidano on the island of Sardinia, Italy (Lazzeri et al. Reference Lazzeri, Mascia, Sammartino, Campus, Caredda, Carlesi, Fois, Gestri, Mannocci, Mazzoncini, Cuena Lombraña and Santinelli2013), and in the coastal area of Aude in France (EPPO 2020; Fried Reference Fried2020), are considered to be invasive and naturalized in nature, respectively. Two unverified records of L. ferocissimum were also made on the Greek island of Crete in 2017 and 2018 (GBIF 2020).
In the United States, L. ferocissimum has been recorded as naturalized, but rare, in California (Calflora 2013; DiTomaso and Healy Reference DiTomaso and Healy2007; GBIF 2020; USDA-NRCS 2019).
Many unverified iNaturalist records of L. ferocissimum have also been made, most notably in the Philippines and Mexico (GBIF 2021). These are both single, unverified iNaturalist records in the City of Davao on the island of Mindanao in the Philippines and on the northern outskirts of Mexico City in Mexico. EPPO (2020) also lists Indonesia and Bolivia as countries with distributions of L. ferocissimum, but it is unclear where these records are derived from. Similar to the records of L. ferocissimum in Namibia, given the complexities of L. ferocissimum and Lycium species identification and taxonomy (Levin et al. Reference Levin, Shak, Miller, Bernardello and Venter2007; McCulloch et al. Reference McCulloch, Mauda, Chari, Martin, Gurdasani, Morin, Walter and Raghu2020; Venter Reference Venter2000), the veracity of all of the distribution records noted should be considered questionable and not necessarily representative of the true range of this species.
It is only widespread and troublesome on a national scale in Australia and New Zealand (Parsons and Cuthbertson Reference Parsons and Cuthbertson2001) (Figure 4C). In New Zealand, it is present on both the North and South Islands, where it is largely restricted to coastal areas; here it persists in large agroecosystem distributions as established hedges, with spread also recorded into forest and scrub reserves (Breitwieser et al. Reference Breitwieser, Brownsey, Ford, Glenny, Heenan and Wilton2019; Popay et al. Reference Popay, Champion and James2010; Timmins and Mackenzie Reference Timmins and Mackenzie1995; Timmins and Williams Reference Timmins and Williams1991) (Figure 4C). In Australia, L. ferocissimum is widespread in coastal to semiarid inland habitats and islands of southern Australia, with records from every jurisdiction (GBIF 2020; Hussey et al. Reference Hussey, Keighery, Dodd, Lloyd and Cousens2007; Noble and Rose Reference Noble and Rose2013; Parsons and Cuthbertson Reference Parsons and Cuthbertson2001) (Figure 4C). It is found predominantly in the southern part of the Australian continent in coastal and island situations (except Queensland). Lycium ferocissimum occurs on islands off the southern half of the Western Australian coastline, along with the islands of the Great Australian Bight and Bass Strait, and on Lord Howe Island and Norfolk Island (Erkelenz Reference Erkelenz1993; Green Reference Green, Orchard and Wilson1994; Keighery Reference Keighery2010; Keighery et al. Reference Keighery, Alford and Longman2002; Lawley et al. Reference Lawley, Lawley and Page2005; Western Australian Herbarium 1998–; Ziegler and Hopkins Reference Ziegler and Hopkins2011). Inland, L. ferocissimum is abundant in areas of New South Wales, Victoria, and South Australia, where it is a common weed of semiarid pastures and rangelands and is often found growing along dry streambeds (Parsons and Cuthbertson Reference Parsons and Cuthbertson2001). It has a lesser, but significant presence in southeast Queensland, southern Western Australia, and Tasmania. Lycium ferocissimum currently does not occur in higher-altitude areas of Australia, having no substantial presence in the Victorian, New South Wales, and Tasmanian alpine areas.
Invasion History
Lycium ferocissimum has not been widely distributed throughout the world and is only considered invasive in Australia and New Zealand, where records indicate that from at least as early as 1845 it was deliberately imported for use as a hedge and windbreak (Parsons and Cuthbertson Reference Parsons and Cuthbertson2001). It was considered naturalized in Australia by the mid-1800s, and New Zealand by the late 1800s, and an invasive plant in Australia by the early 1900s, and in New Zealand by the 1950s (Fuller Reference Fuller1998; Lambert Reference Lambert2015; Parsons and Cuthbertson Reference Parsons and Cuthbertson2001; Webb et al. Reference Webb, Sykes and Garnock-Jones1988). In the mid-2010s it was reported as invasive in southern Sardinia, Italy (Lazzeri et al. Reference Lazzeri, Mascia, Sammartino, Campus, Caredda, Carlesi, Fois, Gestri, Mannocci, Mazzoncini, Cuena Lombraña and Santinelli2013), aside small-scale introduced and naturalized populations in the Mediterranean Basin nations of Spain, Morocco, Cyprus, and Tunisia (Gallego Reference Gallego2012; Hand Reference Hand2000; Meikle Reference Meikle1985; Perez Latorre et al. Reference Perez Latorre, Yus Ramos and Dana Sanchez2006; Venter Reference Venter2000). Most recently it was recognized as naturalized in the coastal area of Aude in France in 2019 (Fried Reference Fried2020), increasing concern for the invasive nature of L. ferocissimum in Europe, with an EPPO alert being published in July 2020 (EPPO 2020).
The first reports of L. ferocissimum in Australia consist of its listing on the Tasmanian nursery list in 1845. At that time, several names were used (e.g., Lycium europaeum L.), but there is little doubt reference was being made to L. ferocissimum (Parsons and Cuthbertson Reference Parsons and Cuthbertson2001). Lycium ferocissimum was advocated as an excellent hedgerow and windbreak plant and was deliberately introduced and distributed for this purpose. There remain situations in Australia where it continues to be maintained as a hedgerow. Australian L. ferocissimum populations likely originated in the Western Cape of South Africa (McCulloch et al. Reference McCulloch, Mauda, Chari, Martin, Gurdasani, Morin, Walter and Raghu2020), which is consistent with the most likely historical trade routes and plant production for trade occurring through the port of Cape Town. Occurrence of L. ferocissimum at Camden Park, NSW, was recorded by 1850, and by 1857 and 1858, it was recorded at Hobart and Adelaide botanic gardens, respectively (Parsons and Cuthbertson Reference Parsons and Cuthbertson2001). In Victoria during the late nineteenth century, the planting of L. ferocissimum hedges was a requirement of certain leases in the Western District. However, by 1904 the species was declared noxious in certain areas of Victoria (Parsons and Cuthbertson Reference Parsons and Cuthbertson2001). Lycium ferocissimum was first noted as a pest in Queensland in 1917 (Fuller Reference Fuller1998). In 1925, the South Australian Government enacted the Noxious Weeds (African boxthorn) Act of 1925 to identify L. ferocissimum as noxious (except where it had been grown as a hedge before the commencement of the Act) (Government of South Australia 2004). Various invasive plant and weed legislations have been imposed in regard to L. ferocissimum over the past century in Australia. Currently, L. ferocissimum is recognized as a Weed of National Significance (Australian Weeds Committee 2013) and is a declared or controlled weed in all jurisdictions (Agriculture Victoria 2017; Biosecurity SA Reference Biosecurity2017; Business Queensland 2016; Department of Agriculture, Water and the Environment 2021; Department of Environment and Natural Resources 2018; Department of Primary Industries Parks Water and the Environment 2019; Department of the Environment Climate Change Energy and Water 2009; NSW DPI 2019), except Western Australia (Department of Primary Industries and Regional Development 2019).
Lycium ferocissimum was introduced to New Zealand in the 1870s as a hedging plant (Hoskin Reference Hoskin2006; Wishart Reference Wishart2018). It was offered for sale through the Caledonia Nursery in New Plymouth from at least 1883, and there are records of it being propagated in the nursery industry in the 1920s (Hoskin Reference Hoskin2006). It proved particularly popular in the Taranaki region, with some historical hedges still in existence today (Lambert Reference Lambert2015). Boxthorn honey and jam were popular products produced from the species, and the wood was found to burn extremely hot and was used in domestic fires (Hoskin Reference Hoskin2006). Similar to the Australian experience, farmers soon found the species to be weedy and problematic to manage, especially before the advent of mechanical hedge trimming (Brown Reference Brown1979; Hoskin Reference Hoskin2006; Lambert Reference Lambert2015). Lycium ferocissimum hedges are still widespread in New Zealand, albeit becoming reduced in number (Hoskin Reference Hoskin2006). The presence of these hedges in proximity to cropping regions has proved problematic in the management and control of pests such as the tomato-potato psyllid, B. cockerelli (Vereijssen et al. Reference Vereijssen, Smith and Weintraub2018). Lycium ferocissimum is now classified as a plant pest in New Zealand, with restrictions on further propagation of the species across the country (Lambert Reference Lambert2015).
Elsewhere in the world L. ferocissimum may have been introduced in the mid- to late twentieth century, much later than in Australia and New Zealand. Lycium ferocissimum was first recorded in the southern Californian county of Los Angeles in 1978, and to this date has not been recorded outside Los Angeles (Calflora 2013; GBIF 2020), indicating the species may not be especially invasive in this environment. In the Mediterranean Basin, L. ferocissimum was first recorded as deliberately introduced into a plantation in Cyprus in the early 1900s and naturalized there by 1985 (Meikle Reference Meikle1985), in Tunisia in 1964 (Venter Reference Venter2000), Morocco by 1986 (Lambinon and Lewalle Reference Lambinon and Lewalle1986), and as a hedge in Guadalajara, Spain, by 1986 (Perez Latorre et al. Reference Perez Latorre, Yus Ramos and Dana Sanchez2006). The species continues to spread naturally via birds in Cyprus (İlseven and Baştaş Reference İlseven and Baştaş2018) and has been considered as a candidate species for abandoned mine site phytostabilization efforts (Cetinkaya and Sozen Reference Cetinkaya and Sozen2011). The species was well established as a single population in abandoned cropping fields at the mouth of the Vélez River in Malaga, Spain, in the Iberian Peninsula by the time it was first recorded in 2006 (Perez Latorre et al. Reference Perez Latorre, Yus Ramos and Dana Sanchez2006). These plants most likely formed part of an abandoned hedge on the property, and despite the production of large numbers of fruit and surrounding fertile soils, the species has not been observed to spread (Perez Latorre et al. Reference Perez Latorre, Yus Ramos and Dana Sanchez2006). Lycium ferocissimum was most recently recorded in southern Sardinia for the first time in 2012 (Lazzeri et al. Reference Lazzeri, Mascia, Sammartino, Campus, Caredda, Carlesi, Fois, Gestri, Mannocci, Mazzoncini, Cuena Lombraña and Santinelli2013), with at least seven established populations of between two to several hundred plants. While the nature of introduction of these populations is as unclear, Lazzeri et al. (Reference Lazzeri, Mascia, Sammartino, Campus, Caredda, Carlesi, Fois, Gestri, Mannocci, Mazzoncini, Cuena Lombraña and Santinelli2013) are suitably concerned about continued spread of the species via bird feeding, and research into the biology of L. ferocissimum in Sardinia in order to better manage the species is ongoing (Podda et al. Reference Podda, Santo, Puddu, Biagini and Bacchetta2015).
Dispersal and Establishment
Dispersal of Lycium ferocissimum occurs almost always by seed, though the species has been recorded to spread by vegetative means as well. Broken stem and root fragments can remain viable for months before taking root when in contact with moist soil (Muyt Reference Muyt2001). Seeds are dispersed when animals consume fruit and subsequently excrete viable seed. Seed and vegetative material can be dispersed by water, machinery, agricultural products, and soil (Muyt Reference Muyt2001; Noble and Rose Reference Noble and Rose2013; Parsons and Cuthbertson Reference Parsons and Cuthbertson2001).
The common presence of Lycium ferocissimum around the base of trees, fences, and poles in Australia implicates birds as dispersal agents (Parsons and Cuthbertson Reference Parsons and Cuthbertson2001), and gulls (Laridae) are known to feed on and spread seed in coastal environments (Calvino-Cancela Reference Calvino-Cancela2011). In Australia alone, at least 21 species of birds, along with four mammal species and two reptile species, have been recorded as consuming L. ferocissimum fruit (Noble and Adair Reference Noble and Adair2014) (Table 2). Though many of these records, such as for the southern brown bandicoot (Quin Reference Quin1985), are limited in extent, the relationship between starlings and L. ferocissimum is significant in Australia. The presence of starlings appears to assist the invasive establishment of L. ferocissimum (Noble and Adair Reference Noble and Adair2014). Double endochory of Lycium intricatum Boiss seed has been recorded in the Canary Islands (Nogales et al. Reference Nogales, Delgado and Medina1998). This occurs where shrikes (Lanius excubitor L.) regurgitate pellets containing the remains of lizards (Galotia atlantica Peters & Doria) that have fed on L. intricatum fruit. The seeds from shrike pellets had higher germination rates than those from both uneaten fruits and lizard droppings. The treatment type and retention times in the guts of each species are thought to be the factors influencing germination (Nogales et al. Reference Nogales, Delgado and Medina1998). Whether spread of L. ferocissimum is likewise influenced by double endochory is unknown.
Table 2. Fauna recorded to consume Lycium ferocissimum fruit in Australia (Noble and Adair Reference Noble and Adair2014).
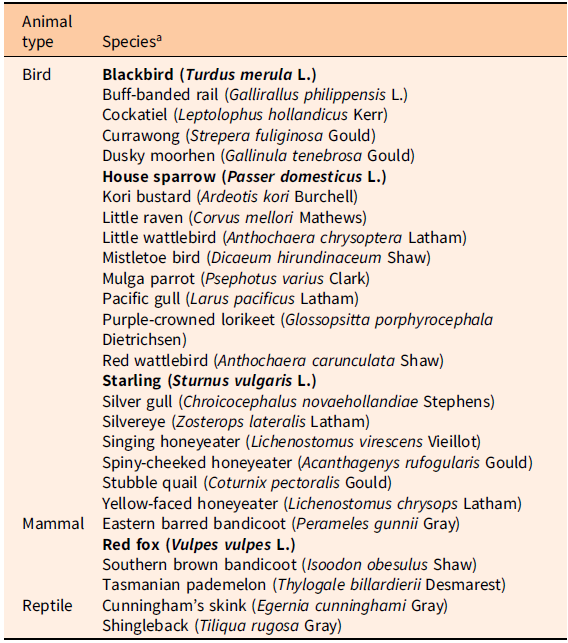
a Introduced species are listed in bold.
Invasion Risk and Pathways
Bioclimatic modeling indicates that L. ferocissimum has a limited global distribution, relative to the total area of projected climatically favorable regions in which the species may establish and persist (Kriticos et al. Reference Kriticos, Ireland, Morin, Kumaran, Rafter, Ota and Raghu2021) (Figure 5). These climatically favorable regions are dominated by coastal Mediterranean and humid subtropical climates on all continents. Given the opportunity, L. ferocissimum could establish in climatically favorable areas of the coastal western and inland southeastern U.S. states, the central highlands of Mexico, central-eastern Argentina, much of Uruguay, parts of southeastern Brazil, some southern coastal regions of Chile, and highlands throughout tropical South America, as well as provinces in southwestern China, including Yunnan (Kriticos et al. Reference Kriticos, Ireland, Morin, Kumaran, Rafter, Ota and Raghu2021) (Figure 5). Lycium ferocissimum is not yet widely distributed in climatically suitable invaded regions in Europe and the Mediterranean Basin (Kriticos et al. Reference Kriticos, Ireland, Morin, Kumaran, Rafter, Ota and Raghu2021), and further spread here may be of concern, especially given the relatively recent invasion and proliferation of the species in Sardinia, Italy (Lazzeri et al. Reference Lazzeri, Mascia, Sammartino, Campus, Caredda, Carlesi, Fois, Gestri, Mannocci, Mazzoncini, Cuena Lombraña and Santinelli2013), and naturalization in the coastal area of Aude in France (EPPO 2020; Fried Reference Fried2020).
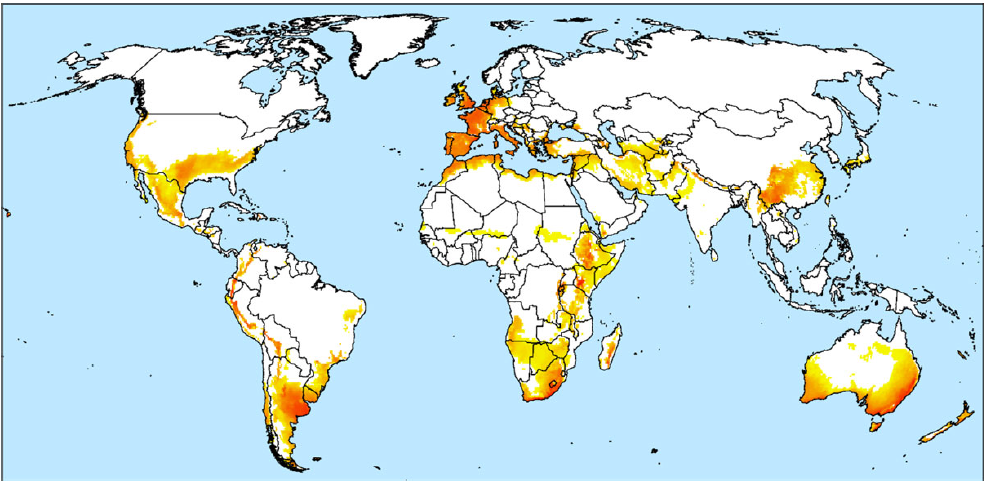
Figure 5. Global projected climatic suitability for Lycium ferocissimum. The projection is under the 1981–2010 climate normals, as modeled using CLIMEX by Kriticos et al. (Reference Kriticos, Ireland, Morin, Kumaran, Rafter, Ota and Raghu2021). Increased intensity of red color, starting from yellow, indicates higher climatic suitability.
While L. ferocissimum is already present and widely distributed through much of its projected highly suitable habitat range in Australia and New Zealand, one exception appears to be southwestern Western Australia (Kriticos et al. Reference Kriticos, Ireland, Morin, Kumaran, Rafter, Ota and Raghu2021) (Figures 4C and 5). There are several potential explanations for this, including the possibility that L. ferocissimum was introduced to Western Australia more recently than elsewhere, or that available distribution data are not as comprehensive as elsewhere, making distribution mapping misleading (Noble and Adair Reference Noble and Adair2014). Another possibility is that the absence thus far of an established population of starlings (Sturnus vulgaris L.) and other dispersers in southwestern Western Australia is restricting the distribution of L. ferocissimum (Harris and McKenny Reference Harris and McKenny1999; Taylor Reference Taylor1968). Nonetheless, indications are that the range of L. ferocissimum continues to extend in southern Western Australia (Abbott et al. Reference Abbott, Marchant and Cranfield2000; Keighery Reference Keighery2010; Keighery et al. Reference Keighery, Alford and Longman2002). Much of the North Island of New Zealand may also be climatically suitable for L. ferocissimum establishment and spread (Kriticos et al. Reference Kriticos, Ireland, Morin, Kumaran, Rafter, Ota and Raghu2021) (Figure 5), although intensive management here may be reducing its spread.
While it has been suggested that the potential distribution of L. ferocissimum may be limited to areas where the plant is intentionally planted (CABI 2018), no specific study of the pathways of introduction and establishment of the species has been conducted, and this hypothesis has not been tested. The recent incursion and spread of the species on the island of Sardinia (Lazzeri et al. Reference Lazzeri, Mascia, Sammartino, Campus, Caredda, Carlesi, Fois, Gestri, Mannocci, Mazzoncini, Cuena Lombraña and Santinelli2013), as well as continued spread throughout much of Australia outside regions where it was historically planted as a hedge, suggest that given the chance to flower and fruit, the species can become quite invasive and abundant in climatically and edaphically suitable regions of the world. Natural competition and enemies within the native range likely keep the species in check, but it would appear that the species may have the capacity to spread further east and northward and establish more abundant populations in the South African provinces of Kwazulu Natal and Mpumalunga should some of these limitations be removed (Kriticos et al. Reference Kriticos, Ireland, Morin, Kumaran, Rafter, Ota and Raghu2021). Where naturalized populations have been recorded in California, USA, close to the metropolis of Los Angeles (GBIF 2018), there seem to be few areas of suitable habitat for the species to spread into, which may explain its limited distribution in this region. In the Mediterranean Basin, the species may be able to spread farther into areas projected to be climatically suitable along the northern coasts of Morocco, Algeria, and Tunisia, much of Mediterranean Europe, and perhaps as far north as the south of the United Kingdom and into the Netherlands and northwest Germany. Numerous populations of L. ferocissimum in coastal regions of Sardinia in Italy and Aude in France are of concern, as spread of the species may accelerate as fruiting increases and birds feed and disperse the seed to neighboring islands and coastlines. The existence of L. ferocissimum on the Greek island of Crete reported in 2017 and 2018 (GBIF 2020) could prove problematic if verified, especially given the projected climatic suitability of this island and coastal mainland Greece (Kriticos et al. Reference Kriticos, Ireland, Morin, Kumaran, Rafter, Ota and Raghu2021).
Habitat
Climate
Lycium ferocissimum occurs across a diverse range of temperate (Group C) and dry (Group B) Köppen-Geiger climate types (Hijmans et al. Reference Hijmans, Cameron, Parra, Jones and Jarvis2005; Köppen Reference Köppen1936; Kriticos et al. Reference Kriticos, Webber, Leriche, Ota, Macadam, Bathols and Scott2012). In South Africa, L. ferocissimum predominantly occurs in temperate oceanic (Cfb), cold semiarid (BSk), and warm-summer Mediterranean (Csb) climatic areas, though the recorded species’ distribution does extend into areas characterized as hot semiarid (BSh), hot (BWh) and cold (BWk) desert, subtropical highland (Cwb; also in Lesotho), and humid subtropical (Cfa) climates. In Australia, the species predominantly occurs in warm-summer Mediterranean (Csb), as well as cold semiarid (BSk) climatic areas, with some extensions into temperate oceanic (Cfb), hot-summer Mediterranean (Csa), and humid subtropical (Cfa) climates. In New Zealand, the species only occurs in the temperate oceanic (Cfb) climatic zone, which encompasses the whole country. In California, L. ferocissimum has been recorded in predominantly cold semiarid (BSk) regions, along with singular sightings in hot (Csa) and warm (Csb) summer climates. Most climatic areas where L. ferocissimum occurs in the Mediterranean Basin are hot-summer Mediterranean (CSa) climates, including the southern provinces of Sardinia, where the species is considered invasive.
Plant Community
Lycium ferocissimum is associated with a range of other flora, including coastal fynbos native vegetation in South Africa (Cowling et al. Reference Cowling, Richardson and Paterson-Jones1995), meadows along riverbeds in Spain (Perez Latorre et al. Reference Perez Latorre, Yus Ramos and Dana Sanchez2006), and salt-tolerant shrubs and grasses on Middle Island, off the coast of southern Australia (King et al. Reference King, Wallis, Wallis, Peucker and Williams2015). In Cyprus it is often recorded in association with Eucalyptus spp. (İlseven and Baştaş Reference İlseven and Baştaş2018; Peyton and Mountford Reference Peyton and Mountford2015), an observation also made in Australia, which may relate to roosting sites for birds that have fed on L. ferocissimum berries. In Australia, Lycium ferocissimum is found in a breadth of plant communities from semiarid rangelands through to coastal vegetation and offshore islands (Noble and Rose Reference Noble and Rose2013).
Substrate and Soil Types
In its native range in southern Africa, L. ferocissimum is found on sandy soils and dry stony flats (Coates Palgrave et al. Reference Coates Palgrave, Moll, Drummond and Coates Palgrave2002; Cowling et al. Reference Cowling, Richardson and Paterson-Jones1995; Webb et al. Reference Webb, Sykes and Garnock-Jones1988). Outside its native range, it has been recorded growing on all soil types, including saline, sandy, volcanic, undisturbed and disturbed, and low- and high-fertility soils (Dodson Reference Dodson1974; Haegi Reference Haegi1976; Lee Reference Lee1978; Parsons and Cuthbertson Reference Parsons and Cuthbertson2001; Perez Latorre et al. Reference Perez Latorre, Yus Ramos and Dana Sanchez2006; Webb et al. Reference Webb, Sykes and Garnock-Jones1988). It can occur as a halophyte in the presence of sandy or volcanic soils in Australia, New Zealand, Cyprus, and Italy (Dodson Reference Dodson1974; Haegi Reference Haegi1976; Lazzeri et al. Reference Lazzeri, Mascia, Sammartino, Campus, Caredda, Carlesi, Fois, Gestri, Mannocci, Mazzoncini, Cuena Lombraña and Santinelli2013; Peyton and Mountford Reference Peyton and Mountford2015; Webb et al. Reference Webb, Sykes and Garnock-Jones1988). Interestingly, the distribution of L. ferocissimum in southern Queensland is centered on the fertile soils of the Darling Downs and on fertile dry scrub soils west of Ipswich, indicating a preference for more arable soils in that state (DAF 2020; Lee Reference Lee1978). It is often found close to permanent or seasonal water supplies and dry creek beds in arid and semiarid environs (Haegi Reference Haegi1976; Parsons and Cuthbertson Reference Parsons and Cuthbertson2001), indicating that water availability may be more restricting than soil type for this species. In New Zealand, it is widespread in coastal areas and is notably capable of colonizing unstable dunes (Webb et al. Reference Webb, Sykes and Garnock-Jones1988).
Lycium ferocissimum thrives on sites where soil disturbance has occurred (Cetinkaya and Sozen Reference Cetinkaya and Sozen2011; Erkelenz Reference Erkelenz1993; Peyton and Mountford Reference Peyton and Mountford2015; Rutherford et al. Reference Rutherford, Powrie and Husted2014), including low-fertility roadsides, pastures, and within abandoned cropping areas (Lee Reference Lee1978; Matthews Reference Matthews, Holzner and Numata1982; Perez Latorre et al. Reference Perez Latorre, Yus Ramos and Dana Sanchez2006). In a South African study, L. ferocissimum was the one shrub species that persisted in the face of high-intensity grazing and increases in most soil nutrient levels (Rutherford et al. Reference Rutherford, Powrie and Husted2014). Where it infests coastal and island environments containing dense bird colonies, evidence suggests that L. ferocissimum may be tolerant of highly nutrient-rich soils (and perhaps soils with high pH) (Gillham Reference Gillham1963).
In relation to soil types, L. ferocissimum is predominantly associated with low-fertility sandy soils (xersosols, desert soils with low levels of organic matter; and arenosols, low-fertility sandy soils), and to a lesser extent with clay-based soils, which may lose fertility quickly under cultivation (luvisols), and soils known to limit root growth (leptosols) within its native range of South Africa (FAO 2007; GBIF 2018). By contrast, in Australia, L. ferocissimum is associated with largely undifferentiated soils (regosols), infertile acidic soils (podzols), and low-fertility sandy soils (xersosols), and to a lesser extent with soils associated with inland water bodies or proximity to the ocean and clay-based soils, which may lose fertility quickly under cultivation (luvisols). In New Zealand, L. ferocissimum is largely associated with tropical “brown soils” (cambisols), which are also associated with rapid losses in fertility and degradation when under cultivation (similar to luvisols), and to a lesser extent with undifferentiated soils (regosols), nutrient-rich volcanic soils (andosols), and clay-based soils (luvisols).
Lycium ferocissimum was also found to most commonly be associated with medium- to coarse-textured soils (FAO 2007; GBIF 2018). Medium-textured soils were dominant in their associations with L. ferocissimum in South Africa and New Zealand, while coarse-textured soils were dominant in this association in Australia. In regard to slope, L. ferocissimum was found to most commonly be associated with level to hilly landscapes (slopes ∼0% to 30% gradients) in South Africa and Australia, but more commonly with steep slopes (>30% gradient) in New Zealand (FAO 2007; GBIF 2018).
While it is well understood that L. ferocissimum can tolerate a wide range of soil types and conditions, the most suitable soils for the species to thrive have not been identified.
Land Use Associations
In relation to land use, L. ferocissimum is predominantly associated with shrubs, both in unmanaged systems and with high livestock density, as well as with crops and high livestock density, both globally and in Australia (FAO 2010; GBIF 2018). It is associated to a lesser extent with unmanaged open water, urban land, rainfed crops, forests with agricultural activities or moderate to high livestock densities, and sparsely vegetated areas with moderate or high livestock densities. In New Zealand, L. ferocissimum was more commonly associated with urban land, which was also the predominant land use category associated with the species in its native range of South Africa.
Life History
Predominantly a stress tolerator with competitive tendencies (sensu Grime Reference Grime1977), L. ferocissimum can survive and thrive across a range of climatic and edaphic conditions, as discussed earlier. In response to stress (i.e., reduced soil moisture, herbivory, or mechanical damage), plants will shed leaves, presumably to conserve energy reserves, and effectively become dormant until conditions become more conducive to growth. While no field studies of the life cycle of the species have been conducted, seeds can germinate quickly within 1 to 3 wk under optimal conditions, growing into seedlings greater than 10-cm high within the first few months before producing multiple stems and growing into a woody shrub/tree habit over the following years (as observed by the authors, in laboratory and field studies) (Figure 6). Mature plants are typically sprawling, approximately 1 to 3 m in diameter, sprawling at the base, with open to closed canopies (especially when actively hedged). While individual plants are often no taller than 2-m high within the native range, plants can form dense, high (3- to 5-m) thickets, especially in the invaded ranges of southeastern Australia. Leaves, while occasionally larger in seedlings (<6 mo), are generally small and often leathery in the field. Flowering and subsequent fruiting may occur year-round, though they have most commonly been recorded as occurring in summer (Parsons and Cuthbertson Reference Parsons and Cuthbertson2001). Seed production has been recorded in plants from 2 yr of age (Parsons and Cuthbertson Reference Parsons and Cuthbertson2001) and may become prolific from the third year (Blood Reference Blood2001) (Figure 6).

Figure 6. Life cycle of Lycium ferocissimum, with putative timeline from seed germination (under optimal conditions) to seed production.
Growth and Development
Root growth in young plants is rapid. Small, fleshy leaves are protected from browsing by woody stems and branches that end in sturdy thorns that are 20- to 150-mm long (Noble and Rose Reference Noble and Rose2013). The rounded thorny L. ferocissimum shrubs are resistant to browsing and largely impenetrable to larger fauna. The woody, branched taproot is an important contributor to competitiveness with other species, resprouting vigorously if cut or broken (CRC Weed Management 2007; Muyt Reference Muyt2001; Parsons and Cuthbertson Reference Parsons and Cuthbertson2001). Damaged plants will reshoot from the base and sucker from roots to regrow and form dense thickets (Muyt Reference Muyt2001; Parsons and Cuthbertson Reference Parsons and Cuthbertson2001).
Physiology and Phenology
Available information on the physiology of Lycium ferocissimum is very limited. In Australia, it is considered tolerant of drought (Muyt Reference Muyt2001) and is found naturalized in semiarid subtropical areas through to areas that experience annual average rainfall of up to 1,200 mm (such as the northern coast of Tasmania). Growth experiments conducted under controlled conditions indicate that the optimal growth temperature range is likely in the range of approximately 15 to 21 C, with likely slowing of growth at below 10 C and above 29 C (Kriticos et al. Reference Kriticos, Ireland, Morin, Kumaran, Rafter, Ota and Raghu2021). While tolerant of salinity (Webb et al. Reference Webb, Sykes and Garnock-Jones1988), L. ferocissimum is considered to be intolerant of full shade (Muyt Reference Muyt2001; Timmins and Mackenzie Reference Timmins and Mackenzie1995). Rutherford et al. (Reference Rutherford, Powrie and Husted2014) assessed resilience of South Africa’s subtropical thicket in the face of herbivore-driven degradation, undertaking monitoring of plant species responses in high and low herbivore browsing situations. Lycium ferocissimum was found to recover in the high browsing pressure situations, but not under low browsing pressure. Shade intolerance is considered the factor that may have excluded it from the low browsing pressure (closed thicket) situation (Rutherford et al. Reference Rutherford, Powrie and Husted2014).
Lycium ferocissimum is drought deciduous. In South Africa’s Western Cape, it occurs in a winter rainfall (Mediterranean) climate, and plants shed their leaves at the start of summer (Cowling et al. Reference Cowling, Richardson and Paterson-Jones1995). However, where winter rainfall does not dominate (i.e., rainfall is nonseasonal), such as on the southern coast of South Africa, plants can remain evergreen in similar coastal environments (Cowling et al. Reference Cowling, Richardson and Paterson-Jones1995). Lycium ferocissimum has been recorded as being sometimes winter deciduous in Australia, depending upon location and seasonal conditions (Parsons and Cuthbertson Reference Parsons and Cuthbertson2001).
Flowering and fruiting occur primarily in summer but can occur at any time of year if sufficient moisture is available (Parsons and Cuthbertson Reference Parsons and Cuthbertson2001).
Reproduction
Floral Biology and Pollination
While no specific studies of natural L. ferocissimum pollination have been conducted, species in the genus Lycium are predominantly insect pollinated (Galetto et al. Reference Galetto, Bernardello and Sosa1998; Miller et al. Reference Miller, Levin and Feliciano2008). Lycium ferocissimum is strongly self-incompatible. In controlled pollination experiments, 75% of cross-pollinated flowers produced fruit, compared with only 8% producing fruit following self-pollination (Miller et al. Reference Miller, Levin and Feliciano2008).
Seed Production
Lycium ferocissimum is hermaphroditic (Minne et al. Reference Minne, Spies, Venter and Venter1994). Seed production occurs in plants from 2 yr of age (Parsons and Cuthbertson Reference Parsons and Cuthbertson2001) but only becomes prolific from the third year (Blood Reference Blood2001). Each fruit contains up to 70 seeds, with larger fruit containing more seeds (Erkelenz Reference Erkelenz1993). Large L. ferocissimum plants can produce thousands of fleshy fruits annually (Muyt Reference Muyt2001; Parsons and Cuthbertson Reference Parsons and Cuthbertson2001).
Seed Viability, Germination, and Seedbanks
The very limited information available on L. ferocissimum seedbank dynamics suggests that seed is not long-lived in the soil (CRC Weed Management 2007; Herbiguide 2015; Western Australian Herbarium 1998–). This being said, in some areas seeds can germinate year-round (Muyt Reference Muyt2001; Parsons and Cuthbertson Reference Parsons and Cuthbertson2001), and mass seed germination has been recorded following disturbance of soil containing seeds (Western Australian Herbarium 1998–). Under controlled conditions, seed germination for L. ferocissimum appears to be highest at 15 to 20 C with 12- to 14-h photoperiods. A study of Sardinian populations showed that L. ferocissimum is capable of germinating within the temperature ranges of 5 to 25 C, with peak germination (˜85% to 90%) recorded when seeds were exposed to temperatures in the range of 10 to 20 C with a 12-h photoperiod (Podda et al. Reference Podda, Santo, Puddu, Biagini and Bacchetta2015). Germination was higher under a 12-h photoperiod across all experiments (though not significant at 5 C) when compared with completely dark treatment regimes (Podda et al. Reference Podda, Santo, Puddu, Biagini and Bacchetta2015). Podda et al. (Reference Podda, Santo, Puddu, Biagini and Bacchetta2015) also found that L. ferocissimum seeds were quite tolerant of saline conditions; germination was recorded even at the highest salt concentration of 500 mM NaCl, though there were some issues of seedling viability following this exposure. Experience growing L. ferocissimum at the Commonwealth Scientific and Industrial Research Organisation (CSIRO) in Canberra has shown germination to be reliable when seeds are soaked for at least 24 h in a 500-ppm solution of gibberellic acid (Sigma-Aldrich, St Louis, MO, USA), sown directly into moist seed-raising mix (Plugger 111 Seed Raising Mix, Australian Growing Solutions, Tyabb, VIC, Australia) and kept at temperatures of 15 or 21 C with 12- to 14-h photoperiods under LED plant growth lights.
Vegetative Reproduction
Lycium ferocissimum can reproduce vegetatively in the field. As noted earlier, broken stem and root fragments can remain viable for months before taking root when brought in contact with moist soil (Muyt Reference Muyt2001). Propagation from woody or green cuttings has not been successful however (<1% strike rate), despite success with striking cuttings of L. australe and L. chinense (see Ireland et al. Reference Ireland, Hunter, Wood, Delaisse and Morin2019a). Lycium ferocissimum recovers readily from fire by resprouting from an extensive and resilient rootstock (Figure 7) and was found to suffer less than 30% mortality in 100% leaf-scorch fire conditions (Choate Reference Choate1997).

Figure 7. Lycium ferocissimum resprouting from rootstock following wildfire, Tasmania. (Photo credit: Karen Ziegler.)
Management Options
Long-term effective control of L. ferocissimum requires a combination of treatments over many years due to the capacity of the species to regenerate from rootstock, stems, and seed. Lycium ferocissimum seed is dispersed predominantly by birds and other animals, and potential for reinfestation of sites from outside sources should be considered in management planning. Considerations and a planning approach are provided in the African Boxthorn National Best Practice Manual: Managing African Boxthorn (Lycium ferocissimum) in Australia (Noble and Rose Reference Noble and Rose2013).
Physical Control Techniques
Physical control of L. ferocissimum includes winching, pulling, bulldozing, stick raking, blade plowing, and cultivation (Noble and Rose Reference Noble and Rose2013). These techniques are best used when L. ferocissimum plants are not carrying seed (or are carrying minimal seed). Otherwise, fresh seed is likely to be deposited into freshly disturbed soil.
Winching and pulling are the lowest-impact physical control techniques for situations where disturbance is a concern, such as where L. ferocissimum is growing within native vegetation. Winching involves connecting a chain or cable to the base of large individual plants and using consistent force to remove the plant and as much root matter as possible (Figure 8A). Pulling or plucking involves using hydraulic arms based on a front-end loader, articulated loader, or excavator to remove individual plants and as much root matter as possible. A hydraulically operated puller has been commercially developed specifically for use on L. ferocissimum (Figure 8B). Bulldozing, stick raking, and blade plowing are suitable in less sensitive landscapes (e.g. pasture) and provide a rapid control method for moderate to heavy infestations.

Figure 8. Physical control techniques (A) and machinery (B) for Lycium ferocissimum. (A) Winching out an individual Lycium ferocissimum plant. (B) Lycium ferocissimum puller. (Photo credits: (A) Jon Fallaw; (B) Rob Higgins.)
Successful management of L. ferocissimum using the techniques noted is dependent on follow-up application of herbicide. This includes the cut-stump technique with immediate application of herbicide for any remaining base/roots after winching and pulling. For all physical control techniques, there is a need to return periodically and carry out foliar spray application and/or machine-based cut stump treatments until there is no regrowth or seedling presence (Noble and Rose Reference Noble and Rose2013).
Chemical Control Techniques
Chemical control of L. ferocissimum uses techniques including foliar spraying, cut stump application (including mechanical cut stump), stem injection, stem scrap or frilling (e.g., using chisel cuts), basal bark application, and soil–root zone application.
Foliar spraying of triclopyr–picloram herbicide mixes and of triclopyr, picloram, and aminopyralid are commonly undertaken for management of L. ferocissimum. Glyphosate, glyphosate–metsulfuron-methyl mix, and picloram and 2, 4-D amine-based herbicides can also be foliar sprayed on L. ferocissimum. Adjuvants improve herbicide uptake by the plant. Mature L. ferocissimum plants demonstrate strong resilience to foliar spraying, with new foliage readily appearing on plants that had seemingly “died off” after spraying. Follow-up applications are usually essential (Noble and Rose Reference Noble and Rose2013).
In situations where vehicular access is impractical or undesirable (such as coastal and island situations), the cut stump technique is often used. Triclopyr–picloram mix (in diesel), triclopyr (mixed with diesel), and picloram are suitable for this technique. Glyphosate (mixed 1:1 with water) has been used successfully with the cut stump technique over many years by Friends of Bass Strait Islands (Ziegler and Hopkins Reference Ziegler and Hopkins2011). A significant advantage of the cut stump technique is that it allows for the removal and destruction of L. ferocissimum debris. Where dead L. ferocissimum debris is left in place, this continues to provide habitat for pest animals such as starlings and rabbits. Also, the thorns continue to present a hazard to people, wildlife (fatally ensnaring birds), and livestock and can potentially puncture car tires.
Picloram-based herbicides can be used for stem-injection application, while glyphosate-based herbicides can be used for frilling or stem-scrape techniques (Noble and Rose Reference Noble and Rose2013). Triclopyr, along with triclopyr–picloram-based herbicides (mixed with diesel) can be applied to L. ferocissimum using the basal bark application technique.
Soil–root zone herbicide application is not suitable in environmentally sensitive areas, as there is significant potential for off-target damage. Hexazinone- or tebuthiuron-based herbicide is applied to the soil near the drip line of the target plant.
Natural Enemies/Biological Control
Lycium ferocissimum was first evaluated comprehensively for potential biological control in Australia in 2013 (Adair Reference Adair2013), and since 2016 CSIRO in Australia has undertaken further research into candidate biological control agents. A variety of generalist plant pests and pathogens have also been recorded on L. ferocissimum, both within the native and nonnative ranges. Native range surveys for candidate agents in South Africa have revealed four promising agents to date: one rust fungus, Puccinia rapipes Berndt & E. Uhlmann Reference Berndt and Uhlmann2006 (Ireland et al. Reference Ireland, Hunter, Wood, Delaisse and Morin2019a), and three leaf-feeding beetles, Cassida distingeuenda Spaeth (Chrysomelidae), Cleta eckloni Mulsant (Coccinellidae), and Neoplatygaster serietubertculata Gyllenhal (Curculionidae) (Chari et al. Reference Chari, Mauda, Martin and Raghu2020).
The abundance of natural enemies is low in Australia, with herbivory levels on leaves, stems, and fruits of L. ferocissimum at <1% (Adair Reference Adair2013). Fruits are often damaged by birds and either wholly or partially eaten. Fourteen insect and five pathogen associations with L. ferocissimum have been recorded in Australia (see Adair Reference Adair2013). All are generalist species, with many known as pests from commercially utilized crops or ornamental plants, including pests of biosecurity importance such the Mediterranean and Queensland fruit flies (Ceratitis capitata Wiedemann and B. tryoni, respectively). The tomato-potato psyllid, B. cockerelli, a serious pest of Solanaceae species, has been found feeding on L. ferocissimum in New Zealand and North America and may be a putative host of the destructive bacterium CLso, which is vectored by the psyllid (Liefting et al. Reference Liefting, Weir, Pennycook and Clover2009; Munyaneza Reference Munyaneza2012; Vereijssen et al. Reference Vereijssen, Smith and Weintraub2018). Bactericera cockerelli is now established in Western Australia (Commonwealth of Australia 2018), and while no associations with L. ferocissimum have been reported thus far, it would be likely that the pest would also feed and reproduce on this host in Australia. A new species of psyllid (Acizzia sp.) was recorded from L. ferocissimum in Australia in 2013 (Adair Reference Adair2013). This is the first record of a psyllid feeding on Australian Lycium and the first record of Acizzia from Lycium. Three other Acizzia species have recently been recorded from other Solanaceae in Australia (Kent and Taylor Reference Kent and Taylor2010; Taylor and Kent Reference Taylor and Kent2013). Acizzia are predominantly associated with Acacia (Taylor and Kent Reference Taylor and Kent2013). The Lycium Acizzia is widespread in the Australian state of Victoria, and densities on L. ferocissimum are often high, causing considerable damage by stunting growth and though chlorosis, distortion of foliage, and contraction of internodes. Stem death may also occur. The psyllid may also be widespread in other states and territories, but further surveys are required to determine its full distribution. It is not known at this stage whether Australian populations of the insect are vectors of any Candidatus bacteria. The mealy bug Pseudococcus sp. commonly shelters and feeds among the compacted growth caused by Acizzia, further contributing to stem weakening.
Puccinia rapipes (rust fungus)
The rust fungus P. rapipes was identified early as a candidate biological control agent (Figure 9), given that it had only been recorded as occurring on L. ferocissimum (Berndt and Uhlmann Reference Berndt and Uhlmann2006) and that rust fungi have historically performed well as biological control agents (Cullen et al. Reference Cullen, Kable and Catt1973; Hayes et al. Reference Hayes, Fowler, Paynter, Groenteman, Peterson, Dodd and Bellgard2013; Morin and Scott Reference Morin, Scott, Julien, McFadyen and Cullen2012; Morris Reference Morris1997). The fungus infects predominantly young, fleshy leaves, impacting host health and growth through direct loss of photosynthetic tissues where the fungus occurs, and draining host resources to maintain fungal growth. Host-specificity testing in Australia revealed that P. rapipes can also infect the Eurasian origin goji berries L. barbarum, L. chinense, and L. ruthenicum (Ireland et al. Reference Ireland, Hunter, Wood, Delaisse and Morin2019a). Importantly, P. rapipes does not infect the Australian native boxthorn, L. australe. These results indicate that P. rapipes may be sufficiently host specific to pursue as a biological control agent in an Australian context, should regulators be willing to accept damage to the Eurasian goji berries that are grown, albeit to a limited extent, in Australia.

Figure 9. Puccinia rapipes pathogen on Lycium ferocissimum. (Photographed in South Africa by Alan Wood.)
General Outlook
Lycium ferocissimum is a widespread and significant weed in southern Australia and New Zealand that could spread further into suitable coastal Mediterranean and subtropical humid climates around the world. The climatically suitable regions at risk include coastal western and inland southeastern states of the United States, the central highlands of Mexico, central-eastern Argentina, much of Uruguay, parts of southeastern Brazil, some southern coastal regions of Chile, and highlands throughout tropical South America, as well as provinces in southwestern China, including Yunnan (Kriticos et al. Reference Kriticos, Ireland, Morin, Kumaran, Rafter, Ota and Raghu2021). The newest populations in Sardinia are considered the most recently invasive, and further risk of spread in the Mediterranean Basin should not be overlooked. The invasive nature of the shrub may be reliant on it gaining a foothold as a hedge species, though extensive spread may be aided by birds, and so further distribution of the species in the nursery trade should be managed with care. Therefore, research on topics such as seed dispersal, as well as survival and longevity of seed following dispersal, would significantly aid weed management planning. Where the species is established, it may be possible to control and manage spread through carefully targeted removal and chemical control follow-up or potentially via biological control, should candidate agents be deemed safe for release into these environments.
Acknowledgments
KBI was supported by AgriFutures Australia (Rural Industries Research and Development Corporation), through funding from the Australian Government Department of Agriculture, Water and the Environment, as part of its Rural Research and Development for Profit program (PRJ-010527). Thanks are extended to the Biology of Invasive Plants editorial team, including Darren Kriticos and Sarah Ward, as well as Michelle Rafter, David Clements, and one other anonymous reviewer, for constructive feedback on previous versions of the article. No conflicts of interest have been declared.











 Open access
Open access