


Outline
In this paper, we will show that planets of Red Dwarf stars (RDPs) are more hospitable to the evolution of biotic life (‘habitable’) than thought before, and demonstrate the implications of the habitability of RDPs on the expected abundance of biotic planets.
In the first (‘astronomy’) sections of this paper, we show how the results of the Kepler space telescope have revolutionized our knowledge of the statistics of extra-solar system planets (exoplanets). We focus especially on red dwarfs (RDs), by far the most numerous stellar population in the Milky Way. We review the recent work showing how conditions on RDPs may be more hospitable to life than thought earlier. In particular, we show that conditions favourable to life may exist on tidally locked RDPs as well as in multiple star systems, and that planetary water and oxygen can survive the relatively strong ultraviolet and X-ray outbursts during the early flare-phase in the evolution of RDs.
In the subsequent (‘biology’) sections, we consider the suitability of RDPs to oxygenic photosynthesis (OP), the process used by plants and other organisms to convert light into chemical energy. On Earth, OP uses radiation in the 400–700 nm waveband, which is considered to have been a prime requirement for the evolution of complex, multi-cellular life. On RDPs, near-infrared radiation (NIR, >700 nm) is far more copious than on planets orbiting Solar-type stars. Many authors have thus described mechanisms whereby photosynthesis could evolve to utilize NIR. We argue that, based on analogy to the evolution of photosynthesis and to the distribution of vegetation on Earth, the visible radiation (<700 nm) incident on RDPs, may be more than adequate to support OP (Gale & Wandel Reference Gale and Wandel2015). Finally, we argue that the product of the huge number of RDPs and their ability to support OP significantly enhances the statistical chance for life supporting planets in our Milky Way galaxy.
In the section ‘Prevalence of Earth-like planets outside the Solar System’, we review the most recent Kepler and other exoplanet data, in particular the implications for the existence of habitable Earth-sized planets of RDs. In the section ‘Potential clement conditions for life on RD planets’, we review the general conditions on RDPs that may allow Earth-like life. In the section ‘Habitability of planets in a multi-star system’, we discuss the habitability of multi-star (in particular RD) systems. The implications on habitability of the energetic flux in the early evolutionary stages of RD stars are given in the section ‘Energetic radiative flux at early evolutionary stages of RDs’. The section ‘Liquid water and tidal locking within Habitable Zone (HZs) of RD stars’, examines the implications of tidal locking to the presence of liquid water and to habitability of RDPs. In the section ‘The importance of OP for the evolution of complex life’, we survey how OP has been vital for the evolution of complex life on Earth. In the section ‘The environment on RD planets in relation to OP and plant growth patterns on Earth’, we discuss the environment on RDPs in relation to OP and plant growth, and make the analogy to certain regions on Earth. Finally, in the section ‘RDs and the abundance of biotic life’, we demonstrate by a quantitative calculation, how including RDPs enhances the abundance of putative biotic planets, potentially suitable for Earth-like life.
Prevalence of Earth-like planets outside the Solar System
In the past decade, planets have been discovered around hundreds of nearby stars, yet most of them were Jupiter-like gas giants, and too close to their host star to permit liquid water on their surface (e.g. Fridlund et al. Reference Fridlund2010). In recent years (2010–2015), the Kepler mission yielded over 4000 exoplanet candidates, most of them with sizes smaller than Neptune and down to Earth-sized planets. Such small planets (super-Earths) have been shown to be abundant and even to constitute the majority of exoplanets (Buchhave et al. Reference Buchhave2012; Batalha et al. Reference Batalha2013). Further work has demonstrated that such planets are often found within the habitable zone (HZ) of their host star. Recent analyses of the Kepler data showed (Petigura et al. Reference Petigura, Howard and Marcy2013) that about 20% of all Solar-type stars have small, approximately Earth-sized planets orbiting within their HZ. Observational uncertainties and false-positive detections (Farr et al. Reference Farr, Mandel and Stroud2014; Foreman-Mackey et al. Reference Foreman-Mackey, Hogg and Morton2014) may significantly reduce this figure (down to 2–4%, however, with a large uncertainty), yet it still implies a significant fraction and a huge number of stars with Earth-sized habitable planets. Similar results have been obtained by different methods. The High Accuracy Radial velocity Planet Searcher (HARPS) team (using the Doppler method) estimated that more than 50% of Solar-type stars harbour at least one planet, with the mass distribution increasing towards the lower mass end (<15 Earth masses) (Mayor et al. Reference Mayor, Marmier, Lovis, Udry, Ségransan, Pepe, Benz, Bertaux, Bouchy and Dumusque2011). HARPS also detected super-Earths in the HZ (Lo Curto et al. Reference Lo Curto, Mayor, Marmier and Lovis2013). These findings demonstrate that ‘Earth-like’ planets (in the sense of Earth-sized planets in the HZ) are probably quite common, enhancing the probability of finding planets with conditions appropriate for the evolution of biological life as we know it.
Another conceptual break-through has been the recognition that life need not be limited to planets orbiting Solar-type stars, which are only a small fraction of all stars in the Galaxy. Especially RD or M-type stars (the lowest-mass stars, 0.08–0.6 MSun, less luminous and cooler than the Sun), are of great interest, as RDs are the most common in the Milky Way galaxy, constituting about 80% of the stars in our neighbourhood. In this review, we focus our discussion on RDPs.
The Kepler results show that a significant fraction of all RDs may have Earth-sized planets (Dressing & Charbonneau Reference Dressing and Charbonneau2013; Reference Dressing and Charbonneau2015). These authors find that RDs have on average {0.56}−0.05 +0.06 Earth-sized planets (1–1.5R⊕) and {0.46}−0.05 +0.07 super-Earths (1.5–2R⊕) with periods shorter than 50 days. Recent estimates of the occurrence of Earth-sized planets in the HZs around Kepler RDs give that over 40% are expected to harbour an Earth-sized planet in their HZ (Kopparapu Reference Kopparapu2013); the more recent work of Dressing & Charbonneau (Reference Dressing and Charbonneau2015) estimates that within a conservatively defined HZ, based on the moist greenhouse inner limit and maximum greenhouse outer limit, RDs have on average {0.16}−0.07 +0.17 Earth-sized planets and {0.12}−0.05 +0.10 super-Earths. Adopting broader HZ boundaries (Venus to Mars) yields a higher estimate of {0.24}−0.08 +0.18 Earth-sized planets and {0.21}−0.06 +0.11 super-Earths per RD. If correct, this would mean that nearly every second star in the Milky Way has a terrestrial planet within its HZ and that the nearest potentially habitable Earth-sized planets could be less than 10 light years away (<3 pc; as this review is intended to be interdisciplinary, we will use light years rather than parsecs, which are more common in astronomy publications). If most Earth-sized HZ RDPs were suitable for life (as we will discuss in the following sections), then this would imply that about half of the stars in the Milky Way could harbour a biotic planet. Given these considerations, one may calculate the likely abundance of life using the analyses of Wandel (Reference Wandel2015).
Potential clement conditions for life on RD planets
Despite the great advances of the last 15 years in the detection of extra-solar system planets, we still have no evidence for the existence of life anywhere but on Earth. Consequently, discussions of life and the ecological potential for life are constrained by the caveat of ‘life as we know it’. This will also be the limitation of the following discussion, although we recognize that life elsewhere may be very different and have environmental–ecological requirements dissimilar to those with which we are familiar (Pohorille & Pratt Reference Pohorille and Pratt2012; Azuo-Bustos & Vega-Martinez Reference Azuo-Bustos and Vega-Martinez2013). In this review, ‘clement conditions’ refer especially to the requirements of OP.
Much single-celled life exists below the surface of Earth, where it utilizes sources of energy other than light (Gold Reference Gold1999). Some complex life has developed on Earth in the dark, in ecosystems originating around sub-ocean hydrothermal vents. These ecosystems are presently exceptional, although very significant, as they may have been the source of Earth's first life (van Dover Reference Van Dover2000). Furthermore, the existence of such hydrothermal life, which utilizes, non-photosynthetic sources of energy, is of great interest in the search for life in ecological niches outside of Earth. This could be either within the Solar System, such as in the subsurface liquid water of the satellites Europa (of Jupiter) and Enceladus (of Saturn) or outside the Solar System, on planets (and their satellites) of other stars.
Early photosynthesis on Earth did not split water and release oxygen (Blankenship Reference Blankenship1992; Hohmann-Marriot & Blankenship Reference Hohmann-Marriott and Blankenship2011). Many examples of such photosynthesis are still extant (e.g. Ritchie & Runcie Reference Ritchie and Runcie2013). Although today much life on Earth is not dependent on OP, nearly all advanced multi-cellular life is: either directly (plants) or indirectly (herbivores, carnivores and parasites). While transmuting radiant to chemical energy and providing food, OP also supports the oxygen-rich atmosphere essential for the aerobic respiration that drives complex life forms and provides protection from excessive ultraviolet radiation (Wayne Reference Wayne1992; and see below). The possibility and conditions for life on RDPs, have been discussed by many authors (e.g. Heath et al. Reference Heath, Doyle, Joshi and Haberle1999; Tarter et al. Reference Tarter2007), long before the discoveries of the Kepler telescope. In the next three sections, we briefly summarize these major ‘astrophysical’ considerations, which show that the basic ingredients of complex life can persist on RDPs. We then argue that conditions favouring OP may exist on RDPs and extend this argument, with reference to conditions in certain analogous ecosystems on Earth.
Habitability of planets in a multi-star system
Most stars are part of binary or multiple systems (65–100% in the middle of the main sequence, e.g. Duquennoy & Mayor Reference Duquennoy and Mayor1991; Duchene & Kraus Reference Duchene and Kraus2013). For low-mass stars such as RDs the fraction may be somewhat lower (e.g. Bardalez et al. Reference Bardalez, Daniella, Gelino and Burgasser2015 find ~45%, but they admit that they may have failed to identify some of the objects in their sample as binary or multiple systems). Even if only half of the apparent RDs were actually binary systems, the fraction of individual RDs in such systems would be ~67%. For the purpose of estimating the abundance of habitable planets, it is therefore important to know whether planets of binary or multiple stars could be habitable.
Early researches suggested that planets in such systems would have highly erratic orbits. This would result in chaotic climates – most unsuitable for the evolution of advanced life forms. However, recent calculations show that planetary orbits in multiple star systems could be stable, producing climates, albeit different from those on Earth, but not impossible for life (Joshi Reference Joshi2003). Thus, multiple star systems need to be taken into account when discussing habitability and the abundance of biotic exoplanets (O'Malley-James et al. Reference O'Malley-James, Raven, Cockell and Greaves2012). Here we show that this argument is even stronger for RD stars.
Planets orbiting much closer to one of the stars of a binary (or multiple) star system than the distance between the stars, are dominated by the radiation from the nearby star. Hence, their climate is relatively weakly influenced by the radiation from the other star(s) in the system (unless the other star(s) is much brighter than the planet's host star). This particular situation is especially likely for RDPs, for two reasons. First, binary systems consisting of two RDs are by far the most common of all multiple star systems. Second, the HZs of RD stars are significantly smaller than those of Solar-type (or other) stars – of the order of ~0.1–0.4 AU, and hence more likely to be smaller than the distances between stars in a binary or multiple system.
In unfavourable orbital configurations, planets in multiple star systems may be dynamically unstable (the planet would be expelled from its orbit relatively quickly, being either ejected from the system altogether or moved to a closer or further-out orbit). Even orbits that are stable may be unsuitable for the evolution of life because of extreme variations in surface temperature during different parts of the orbit. As described above, planets that orbit just one star in a binary pair (a configuration called ‘S-type’ orbit), may have a relatively isothermal biosphere if the HZ is much smaller than the distance between the two stars of the binary, provided the other star is not much brighter.
In general, the condition for a planet in an S-type orbit to be isothermal is
 $${D_{\rm p}} \ll {D_{12}}{\left( {{L_2}/{L_1}} \right)^{1/2}},$$
$${D_{\rm p}} \ll {D_{12}}{\left( {{L_2}/{L_1}} \right)^{1/2}},$$
where D p is the distance of the planet from the host star, L 1 is the luminosity of that star, D 12 is the average distance between the two components of the binary and L 2 is the luminosity of the other star in the binary system. For planets orbiting around both stars (‘P-type’ or ‘circum-binary’ orbits) the condition for thermal stability is obviously D p >> D 12, that is, the planet should be relatively far from the host binary, so it is weakly affected by the changing positions of the two stars relative to the planet.
It is estimated that 50–60% of all binary stars are capable of supporting habitable terrestrial planets within stable orbital ranges. This figure comes from simulations demonstrating that the presence of a binary companion may improve the rate of planet formation with stable orbits by stirring and increasing the accretion rate of the protoplanets within the proto-planetary disc (Quintana & Lissauer Reference Quintana, Lissauer and N.2010). As argued by these authors, detecting planets in multiple star systems is more difficult than around single stars, whether by the Doppler or the transit methods, which may explain the relatively low numbers of planets actually observed in binary or triple star systems. However, the lower rate of planets around multiple stars may be real, as demonstrated by recent analyses of the Kepler database (corrected for observational bias). This could imply that a stellar companion at distances smaller than 1500 AU suppresses planet formation (Wang et al. Reference Wang, Fischer, Xie and Ciardi2014). This observation is supported also by the indirect evidence in close binary systems with RDs, that a close companion may disrupt the proto planetary disc and thus limit or stop planet formation (Meibom et al. Reference Meibom, Mathieu and Stassun2007). On the other hand, in certain extended (>1000 AU) stellar systems planetary systems appear to be as common as around single stars (Zuckerman Reference Zuckerman2014). In conclusion, it presently seems that multiple stellar systems may have habitable planets and hence should be taken into account for estimating the abundance of biotic planets. This conclusion is much stronger if one or more of the stars of a binary (or multiple) system is a RD star. Not only most of the stars in the Milky Way are found in such binaries, but planets in S-type orbits around a RD-member of a binary are more likely to have a stable climate, than those around a Solar-type (or other) member of a multiple-star system.
Energetic radiative flux at early evolutionary stages of RDs
Young RDs have strong coronal X-ray radiation with values of X-ray-to-total luminosity ratio up to 100 times that of Solar-type stars, and exhibit strong coronal ultraviolet radiation flares and ultraviolet surface fluxes up to 100 times those of comparable age (or rotation) Solar-type stars (e.g. Houdebine Reference Houdebine2003). The energy of these flares may be estimated by measuring their ultraviolet lines, in particular the hydrogen Lyman-α line which has been shown to correlate with the ultraviolet emission of RDs (Shkolnik et al. Reference Shkolnik, Rolph, Peacock and Barman2014; Shkolnik & Barman Reference Shkolnik and Barman2014; Jones & West Reference Jones and West2015).
Rugheimer et al. (Reference Rugheimer, Kaltenegger, Segura, Linsky and Mohanty2015) recently examined how ultraviolet emission affects Earth-like planets orbiting M dwarfs, in particular the detectability of their life signatures. They model the atmospheres and spectra of Earth-like planets orbiting RDs with and without ultraviolet radiation, as well as observed RDs with ultraviolet radiation data. They focus on the effect of ultraviolet activity levels on detectable atmospheric features that indicate habitability on Earth, such as H2O, O3, CH4, N2O and CH3Cl.
Taking into account that planets within the HZ of RDs are much closer to the host star (0.1–0.4 AU) than in the case of Solar-type stars (~1 AU), these radiation events could be dangerous for both, the atmosphere and surface life. For example, it has been thought that the strong X-ray and ultraviolet radiation (XUV) activity of RDs could strip the atmosphere off RDPs (Tian Reference Tian2009). Indeed, high ultraviolet radiation and charged particle flux may cause an evaporation of the planet's atmosphere, which could explain the high fraction of hot dense planets around cool stars in the current exoplanet population (e.g. Wu & Lithwick Reference Wu and Lithwick2013). This led many astronomers and astrobiologists to assume that life (as we know it) cannot evolve on RDPs.
However, in recent years the possibility for the evolution of life on planets around RDs has been reassessed (e.g. Tarter et al. Reference Tarter2007). Guinan & Engle (Reference Guinan and Engle2013) found that during the first 3 billion years of the life of a RD, its X-ray radiation level decreases by a factor of 30, bringing it down to the level of Solar-type stars (relative to the optical luminosity). Moreover, the lifetime of RDs is much longer than that of Solar-type stars. Calculations show that, depending on their mass, RDs could live over 100 billion years, much longer than the (current) age of the Universe. Since the huge radiation flares occur only in the early evolution of RDs (e.g. Silvestri et al. Reference Silvestri, Hawley and Oswalt2005; West et al. Reference West, Hawley, Bochanski, Covey, Reid, Dhital, Hilton and Masuda2008), this leaves a long period during which the level of coronal X-rays and radiation flares of RDs are low. Life on Earth took less than 4 billion years to evolve from the first mono-cellular life to multi-cellular complex life. If this is a characteristic duration, it seems that RDPs would experience a long enough low-radiation epoch to allow the evolution of life.
The atmospheric-stripping effect of the XUV radiation during the early epoch of RDs has been reassessed (Erkaev et al. Reference Erkaev, Lammer, Odert, Kulikov, Kislyakova, Khodachenko, Güdel, Hanslmeier and Biernat2013). Recent calculations demonstrated that RDP atmospheres may survive the high XUV radiation epoch. For example, a super-Earth planet with 6–10 Earth masses may retain a primary CO2 atmosphere even while orbiting a young RD with XUV radiation levels of up to 1000 that of the Sun (Tian Reference Tian2009). The latter may be present during the early evolutionary phases of RDs. Atmospheres consisting of lighter gases (such as the H–He massive atmospheres possessed by sufficiently massive planets), or heavier gases in atmospheres of lower mass planets, could be less immune. For example, Lammer et al. (Reference Lammer, Lichtenegger, Khodachenko, Kulikov, Griessmeier, Beaulieu, Dieteres and Tinetti2011) showed that an Earth-sized planet with a nitrogen atmosphere may be affected by a radiation level only ten times the Solar X and ultraviolet radiation, if present for a sufficiently long time, such as the early evolutionary period of low-mass RDs. Therefore, unless rebuilt during the later, more stable stages of the host star, such RDPs may suffer from a rarified atmosphere.
Liquid water and tidal locking within HZs of RD stars
The HZ is defined as the region around a star in which the equilibrium surface temperature of a planet with an atmosphere similar to that of Earth could support liquid water at least on parts of the surface, at least for part of the time. This is a sine qua non for life (e.g. Kasting et al. Reference Kasting, Whitmire and Reynolds1993) – but always with the caveat of ‘life as we know it’. In the Solar System, the HZ extends approximately between the orbits of Venus and Mars, with Earth approximately at its center. Planets with a thicker atmosphere would be able to have surface liquid water further out from the Sun, because of the greenhouse effect. Planets with a significantly higher albedo could support liquid water on orbits nearer to the Sun. It should be noted, however, that liquid water has been detected in other places in the Solar System, on planet satellites (e.g. Europa and Enceladus) far outside the HZ (Kargel et al. Reference Kargel, Kaye, James and Head2000).
Life on RDPs that evolves under water (as did life on Earth for the first 3 billion years) would be protected from flares and XUV radiation, as those would penetrate only the thin upper layer of water. However, even RDP oceans are not immune to evaporation by the radiation from the host RD during its extended pre-main sequence phase, causing a runaway greenhouse effect. During this early runaway phase, photolysis of water on RDPs in the HZ (for Earth-sized and smaller planets) could result in hydrogen/oxygen escape into space of large quantities of water, up to several times that of all the Earth's oceans (Tian et al. Reference Tian, France, Linsky, Mauas and Vieytes2014; Luger & Barnes Reference Luger and Barnes2015). This does not necessarily preclude the existence of surface water on RDPs during later stages. First, the extended runaway scenario may be less important for RDPs at the outer HZ and for higher-mass RDs. Second, water may be acquired by accretion of cometary and asteroidal water after the runaway period, if the latter is shorter than the period of bombardment by comets and icy asteroids. Third, in the case of massive super-Earth RDPs, water may be reproduced from the oxygen and hydrogen retained in the atmosphere after early-on splitting of water.
As noted above, the HZ for planets orbiting RD stars would be much closer to their host star than for Solar-type stars. This proximity would probably result in tidal locking, with only one hemisphere continuously exposed to the star, much as our Moon is tidally locked to Earth (Gladman et al. Reference Gladman, Quinn, Nicholson and Rand1996). It was originally thought that tidally locked planets, having one side continuously illuminated, with the other hemisphere in the dark, could not be hospitable for life, as their dark side would be permanently frozen and their atmospheres would be too turbulent, as a result of planet-wide winds of hurricane proportions. However, recent calculations indicate that even at the light–dark terminator, winds would be mild, especially in the presence of planet oceans, which mediate heat transport (Hu & Yang Reference Hu and Yang2014). Moreover, the drag effect of a massive atmosphere like that of Venus could counteract the tidal locking, and recent calculations show that even a relatively thin atmosphere can drive terrestrial planets’ rotation away from synchronicity (Leconte et al. Reference Leconte, Wu, Menou and Murray2015). Other consequences of tidal locking are discussed below.
The importance of OP for the evolution of complex life
Oxygen is a highly active chemical element; hence its presence in the atmosphere, in more than traces, indicates that it is being continuously produced. On Earth it is a result of the activity of plant OP, especially oceanic cyanobacteria and algae Lee (Reference Lee2008). Cyanobacteria were the first water splitting photosynthetic organisms on Earth. Consequently, oxygen is a major biosignature in the search for life outside the Earth. However, as noted above, significant amounts of oxygen could be produced on young RDPs abiotically (e.g. by near ultraviolet radiation splitting water molecules). This abiotic oxygen could also produce a false biosignature (Tian et al. Reference Tian, France, Linsky, Mauas and Vieytes2014; Luger & Barnes Reference Luger and Barnes2015). Earth's oxygen-rich atmosphere has been both good and bad for life in general and for plants in particular (Gale Reference Gale2009). One of the early effects of OP was inducing ice ages, although their driver was not only the increase of oxygen. The concomitant reduction of atmospheric CO2 and CH4, which absorb NIR radiation also, contributed to planet cooling. The evolution of Earth's oxygen-rich atmosphere is depicted in Fig. 1.
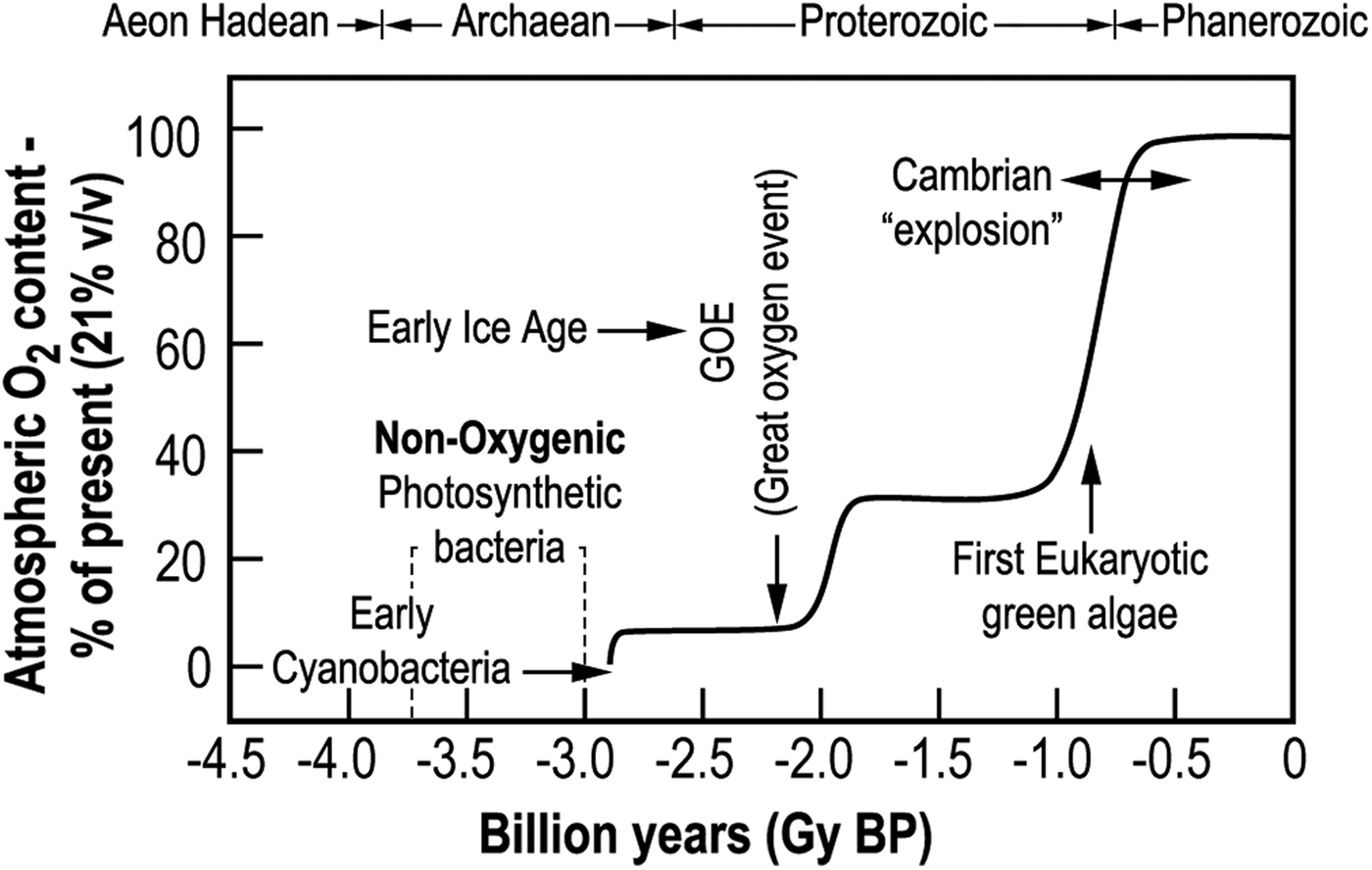
Fig. 1. Time course of the development of Earth's oxygen-rich atmosphere as a percentage of the present oxygen content (21% v/v).
Another negative effect of the oxygen-rich atmosphere on Earth's life is that it inhibits nitrogenase, the enzyme that certain anaerobic bacteria use to fix N2. Although nitrogen is required by all life forms and constitutes about 80% of Earth's atmosphere, this is the only known process by which advanced life can (indirectly) acquire nitrogen from the atmosphere (Raymond et al. Reference Raymond, Siefert, Staples and Blankenship2003).
Today, the OP mechanism (especially in higher plants) is itself partly inhibited in its CO2 fixing activity by the very oxygen it produces. The primary plant enzyme that captures CO2 is ribulose bi-phosphate carboxylase (RUBISCO). At today's atmospheric level of O2 (21% v/v.) the oxygen competes with CO2 (only 0.04% v/v.) for the fixation site. This is true for 80% of the world's vegetation. Only in the very recent 150 million years have new higher plants evolved (of the so-called C4 type) which can overcome this effect. In the majority of C3 plants, oxygen reduces CO2 fixation by about 25% (see e.g. Lambers et al. Reference Lambers, Chapin and Pons1998; Sage Reference Sage2003, for a full discussion).
As noted, the high concentration of O2 in the atmosphere provides protection from ultraviolet radiation (<300 nm). Without it, the unfiltered ultraviolet reaching Earth is energetic enough to cause damaging ionization in bacteria, plant and animal cells. The oxygen in the atmosphere absorbs some of the ultraviolet and in doing so produces ozone, which strongly absorbs ultraviolet radiations. This protective atmosphere appeared on early Earth shortly following the evolution of the cyanobacteria, the first plants which released oxygen from water (Ward et al. Reference Ward, Kirschvink and Fischer2015). It has been called the Great Oxygen Event (GOE) (Fig. 1). Eventually it enabled life to emerge from the protection of the oceans and shaded niches and invade dry land (Wayne Reference Wayne1992; Kasting & Catling Reference Kasting and Catling2003).
The high oxygen atmosphere also enabled the evolution of aerobic respiration, which released nine times more energy (per molecule of carbohydrate or other substrate) than that produced by anaerobic respiration (e.g. Nelson et al. Reference Nelson, Lehninger and Cox2008). A rapid supply of energy is a primary requirement of mechanically active, complex, multi-cellular life. Advanced nervous systems are also heavy expenders of energy. The downside for plants was that this also enabled the evolution of some of the strongest competitors of the Earth's vegetation such as insects and animals. Homo sapiens sapiens is only the latest scourge. Together with the ultraviolet protection of the high O2 atmosphere, aerobic respiration is considered to have been a major factor that drove the Cambrian ‘explosion’ (Catling et al. Reference Catling, Glein, Zahnle and McKay2005). This was a period from ~550 to 470 million years ago when life advanced from the 3.3 billion year era of mainly single-celled organisms to multi-cellular, complex life forms (Fig. 1). For all these reasons, OP and conditions for OP are primary targets in the search for advanced extra-terrestrial life (Cockell et al. Reference Cockell, Raven, Kaltenegger and Logan2009). Although there were (and still are), a number of problems for the biota resulting from the oxygen-rich atmosphere produced by OP, without its appearance on Earth there would probably have been no complex life as we know it.
The environment on RD planets in relation to OP and plant growth patterns on Earth
Our Sun is a G-type star, with a surface temperature of 5860 K. At this temperature, the Solar radiation spectrum incident on Earth after passing through the atmosphere extends from ~250 to 2000 nm. Chlorophyll, the main active pigment of OP in nearly all of Earth's vegetation, has evolved to utilize only about 48% of this radiation, in the waveband extending from 400 to 700 nm, where its absorbance sharply drops off. This is called the photosynthetically active radiation (PAR) waveband (Fig. 2).

Fig. 2. Absorption, action and quantum yield spectra of chlorophyll during OP, showing the sharp cut-off at 700 nm. (Data collected from different sources.)
RDs are late K- and M-type stars. Their surface temperatures are significantly lower than that of G stars, such as our Sun. Consequently, the spectrum of their radiation is shifted towards longer wavelengths, in the NIR. In view of the sharp chlorophyll absorption cut off at 700 nm (Fig. 2), this was first thought to be a precluding factor for OP on RD star planets. However, a considerable part of the radiation would still be in the PAR waveband (Fig. 3). This, together with the continuous illumination on the tidally locked RD planets, could well provide sufficient energy for OP and growth (Figs. 3, 5 and 6).

Fig. 3. Black-body Planck curves of different temperatures, showing the effect of the shifting of the radiation of RD stars towards the NIR, on the energy available in the oxygenic photosynthesis waveband. (All stars have very nearly black-body spectra.)
Contrary to some early and oft-quoted reports (e.g. Emerson & Lewis Reference Emerson and Lewis1943) the OP drop is not caused by lack of energy in photons of wavelength >700 nm. The theoretical energy requirement for splitting a water molecule is 1.23 eV (albeit in reversible systems at standard temperature and pressure and 0 pH), while the quantum energy of a photon at 700 nm is 1.77 eV. The quantum energy of the photon only drops below 1.23 eV at wavelengths >1000 nm. It has also been pointed out that under certain circumstances water splitting may be possible at wavelengths >700 nm in stepped photosynthetic systems (Kiang et al. Reference Kiang, Siefert, Govindjee and Blankenship2007a, Reference Kiang, Segura, Tinetti, Govindjee, Blankenship, Cohen, Siefert, Crisp and Meadowsb) but at a cost of lower overall efficiency (Wolstencroft & Raven Reference Wolstencroft and Raven2002). Splitting of water actually occurs at 680 nm. Part of the quantum energy of the 680 nm photon is also used to drive other electron transfer reactions of photosynthesis. Moreover, some cyanobacteria do carry out OP at wavelengths >700 nm; e.g. Synechococcus leopoliensis has a peak absorbance at 670 nm and uses Chlorophyll a, and Acaryochloris marina has a unique pigment (Chlorophyll d) with peak absorbance at 710 nm (Mielke et al. Reference Mielke, Kiang, Blankenship, Gunner and Mauzerall2011).
The possibility of OP at wavelengths >700 nm raises the question as to why, in 3 billion years of photosynthesis evolution, most Earth plants have not evolved to utilize more than the 400–700 nm waveband, which contains only about 48% of the available Solar electromagnetic radiation. This question is very relevant to RDPs. A first possible explanation is that plants and OP evolved in an environment lacking NIR radiation. A layer of ~15 cm of pure water absorbs almost all NIR >700 nm. The primeval oceans, beneath an atmosphere with little oxygen, did not consist of pure water but also contained other species including, for example, dissolved, reduced Fe++ salts, which also strongly absorb NIR (Curcio & Petty Reference Curcio and Petty1951).
A second explanation is that on dry land lack of radiation energy is rarely a major factor limiting OP and growth of plants from germination to seed. Moreover, Milo (Reference Milo2009), calculated that OP at around 700 nm is the most theoretically efficient in its use of quantum energy. The present OP mechanism expends ~12 photons in the fixation of each molecule of CO2; OP at longer wavelengths would require more (Raven Reference Raven2007). Consequently, there would have been no environmental evolutionary pressure to utilize radiation >700 nm. When plants emerged from sea to land, not only did they not evolve mechanisms to utilize the now available NIR, but actually rejected it. Plant leaves reflect and do not absorb a large part of the incident NIR (Fig. 4).

Fig. 4. Reflectance of a typical plant leaf in the visible and NIR (after Gates et al. Reference Gates, Keegan, Schleter and Weidner1956).
As seen in Fig. 4, about 45% of the 700–2000 nm waveband, which contains >40% of the Solar electromagnetic radiation energy incident on the planet surface, is reflected, while in many plant species a further portion of the NIR is transmitted. Evidently reduction of leaf NIR absorbance, and hence of heating, transpiration and water loss, was more advantageous in the evolution of land plants than extra photosynthesis.
As RDPs in HZs would probably be tidally locked, only plants on the star-facing hemisphere would be able to carry out OP. There is however a clear advantage for such plants as they would be exposed to continuous radiation. The situation is analogous to plants growing on Earth, in the summer months, in far northerly (and southerly) latitudes. In such regions, vegetation is often lush, despite there being only a very short growing season, bordered by long periods of low light and extreme cold. The reason for this burst of growth is that during the short growing season Solar radiation is almost ideal for OP, with moderate light intensities for almost 24 h a day. This contrasts to conditions in the concomitant summer periods at lower latitudes, where for much of the day the radiation intensity exceeds that which can be utilized for OP. This is shown schematically in Fig. 5.

Fig. 5. PAR during the ‘Grand Period of Growth’ incident on northern versus mid-latitude plants, on Earth – an analogue for plants growing on tidally locked RD star planets.
The super-optimum radiation (>P max) during much of the day in low-latitude ecosystems (Fig. 5) may even be damaging to the OP mechanism itself and cause photo-oxidation. Before the onset of photo-oxidation, P max is mainly determined by the capacity of the photosynthesis system (Lambers et al. Reference Lambers, Chapin and Pons1998). Radiation absorption certainly leads to an increase in water loss, which could reduce OP. This happens initially by causing leaf stomata to close, which interferes with the diffusion of CO2 into the leaf, and then, eventually, by damaging the photosynthetic apparatus. Excessive incident radiation would be absent on RDPs as the vegetation on such tidally locked planets would distribute itself to those regions having the radiation intensity most adapted to its species requirements (Heath et al. Reference Heath, Doyle, Joshi and Haberle1999 and Fig. 6).

Fig. 6. The distribution of radiation on the surface of a planet tidally locked to its star. The sub-stellar point corresponds to 0°.
As depicted in Fig. 6, radiation on the curved, star-facing surface of the planet is reduced in accordance with the angle. It would decrease from a maximum, in the region immediately facing the star, to zero at the terminator.
RDs and the abundance of biotic life
We now demonstrate the effect of adding RDPs to the population of potential biotic worlds on the probability of detecting extra-terrestrial life. Consider the distance to our nearest biotic neighbour planets, given by (Wandel Reference Wandel2015)
 $${d_{\rm b}}\sim 10{\left( {{R_{\rm b}}{F_{\rm b}}} \right)^{ - 1/3}}{\rm light}\;{\rm years,}$$
$${d_{\rm b}}\sim 10{\left( {{R_{\rm b}}{F_{\rm b}}} \right)^{ - 1/3}}{\rm light}\;{\rm years,}$$
where F b is the probability of biotic life appearing on an Earth-sized planet in the HZ, and R b is the rate at which habitable planets are formed in the Galaxy
 $$\; {R_{\rm b}} = {R^*}{F_{\rm s}}{F_{\rm p}}{F_{\rm e}}{n_{{\rm hz}}}.$$
$$\; {R_{\rm b}} = {R^*}{F_{\rm s}}{F_{\rm p}}{F_{\rm e}}{n_{{\rm hz}}}.$$
In equation (3), there are two ‘stellar’ factors R* – the rate of star birth in the Milky Way (~10 per year) and F s – the fraction of stars suitable for evolution of life; both are well established. The three other ‘planetary’ factors, F p, F e and n hz were totally unknown up to a few years ago, and could be reasonably well estimated only after the Kepler results. F p is the fraction of stars that have planets, F e the fraction of Earth-sized planets, and n hz is the number of such planets per star, within the HZ.
As discussed in the introduction, the Kepler mission and other recent observations have shown that most, if not all stars have planetary systems, so F p ~ 1.
If only planets around Solar-type stars are considered, then the fraction of stars suitable for evolution of life is the well-known fraction of Solar-type stars in the Milky Way, F s ~ 0.1 (~8% of all main sequence stars are G-type). As discussed above, according to recent analyses of the Kepler data only 2–4% of all Solar-type stars have small, Earth-sized planets orbiting within their HZ (Foreman-Mackey et al. Reference Foreman-Mackey, Hogg and Morton2014; Farr et al. Reference Farr, Mandel and Stroud2014), hence for Solar-type stars F s F e n hz ~ 0.002–0.004. If in addition, one excludes planets around stars in multiple stellar systems (70% or more of all individual stars, in particular RDs, as discussed in the section ‘Potential clement conditions for life on RD planets’), this figure could be smaller by a factor of 3–5, which gives the most severe constraint on the rate at which habitable planets are formed in the Galaxy, R b ~ 10 × 0.2 × 0.002 = 0.004 per year.
However, if in addition RD stars (single or in multiple systems) may host habitable planets, then F s ~ 0.9. According to the analyses of Dressing & Charbonneau (Reference Dressing and Charbonneau2015) and Kopparapu (Reference Kopparapu2013) about half of the RD stars have habitable planets, which leads to much larger figures, F e n hz ~ 0.4 and R b ~ 4 planets per year.
Depending on the assumptions as to which stellar type is suitable for life, the rate R b could thus vary by a factor of 1000, in the range, ~0.004–4. Using equation (3), this would yield a factor of 10 in the estimate of the distance to our nearest biotic neighbour planets. This is shown in Fig. 7, which shows the distance given by equation (2) as a function of the biotic factor F b, for three choices of host-stars. Figure 7 also provides planet-statistics, as discussed above: (a) Solar type, single stars with a low fraction of Earth-sized HZ planets, (R b = 0.004); (b) Solar type, single or multiple stars with a high fraction of Earth-sized HZ planets (R b = 0.1); and (c) single or multiple RDs (R b = 4). It is easy to see that assuming life can evolve on RDPs significantly enhances the abundance of potential biotic planets. For example, if F b = 0.1 the estimate of the distance to our nearest biotic neighbour would vary between 140 light years in the conservative single Solar-type star case and 14 light years in the in the case of a RD, single or in a binary/multiple system.

Fig. 7. The distance to our nearest biotic neighbours as a function of the biotic factor F b, for several choices of host star types and Earth-like planet fractions. The upper, short-dashed curve labelled ‘Solar A’ represents the most conservative case – only single, Solar-type host stars, assuming that 2% of them have Earth-sized planets in the HZ.
The middle long-dashed curve labelled ‘Solar B’ is an intermediate case – Solar-type host star single or in multiple stellar systems, assuming that 20% of them have Earth-sized planets in the HZ. The lower solid curve marked RD depicts RD host stars, single or in binary/multiple systems assuming that 50% of them have Earth-sized planets in the HZ.
As a concrete example we assume the probability of biotic life, F b, to be between 0.001 and 1. Then, according to Fig. 7, the distance to the nearest biotic extra-solar planet could vary in the range of 10–100 light years in the RD case, and 100–1000 light years in the conservative Solar-type host star case. In the near future, the probability F b may be estimated observationally by spectral analyses of the light reflected from exoplanets (e.g. Deming et al. Reference Deming2009; Loeb & Maoz Reference Loeb and Maoz2013), using future telescopes such as the James Webb Space Telescope.
Conclusions
Adding RDPs to the potentially life-supporting planets increases the probability of finding biotic planets by a factor of up to 1000, and reduces the distance to our nearest biotic neighbour by up to 10. Recent works have demonstrated that the HZs of RD stars may be less hostile to life than thought previously. For all the reasons given above, OP and perhaps complex life on planets orbiting RD stars may be possible. Moreover, the vast number of RDPs in the Galaxy seems to make this possibility probable, although to date we still have only the single example of life on Earth. It has been argued that photosynthetic pathways may develop to exploit RD radiation at wavelengths between 700 and 1000 nm, despite lower overall photosynthetic efficiency. However, we suggest here that the evolutionary pressure to do so on RDPs would be small, as tidal locking will provide continuous illumination with a considerable component of photosynthetic 400–700 nm radiation. This is analogous to the lush summer growth of vegetation on Earth, at very high latitudes. Even though, any life that does develop would not be the same as on Earth. For example, there would be no circadian and seasonal rhythms in the flora and fauna, nor would complex life forms have daily sleep patterns. Moreover, extra-terrestrial life may develop using a genetic code different from the single example of life that appeared on Earth – with unpredictable consequences.
The James Webb, Giant Magellan and Thirty Meter (TMT) telescopes, planned to be operational in the next decade, may provide evidence of the life predicted here. This could be obtained by the detection of biomarkers in the atmosphere of planets transiting compact stars. Oxygen produced by OP could be such a biosignature, although remnants of abiotic O2 may confound the results.
Acknowledgement
We thank editor Inge Loes ten Kate and the anonymous referees for many helpful comments.







