



1. Introduction
Hinde (Reference Hinde1890) first described radiolarians from the Southern Uplands of Scotland where they occur in a zone of red and green ribbon-bedded and massive grey cherts, mudstones and shales tectonically intercalated between Llandeilian and Caradocian graptolitic shale beds. He reported and illustrated various forms, providing the first contribution to studies of lower Palaeozoic radiolarians. At that time, studies of microfossils were based largely on optical microscope observations of thin-sections using transmitted light. More than 100 years later, Aitchison (Reference Aitchison1998) examined radiolarians extracted from red cherts of the nearby Ballantrae Complex using scanning electron microscopy (SEM). He described a moderately well-preserved Arenig radiolarian assemblage. Subsequently, Danelian reported moderately well-preserved assemblages of radiolarians from red cherts of the Southern Uplands terrane in the Hawkwood Burn and outcrops of grey cherts near Crawford (Danelian & Clarkson, Reference Danelian and Clarkson1998; Danelian, Reference Danelian1999; Danelian & Floyd, Reference Danelian and Floyd2001). The faunas present were tentatively assigned to the upper Llanvirn – Caradoc Haplentactinia juncta – Inanigutta unica assemblage according to Nazarov’s (Reference Nazarov1988) biozonation. In addition, they were considered to show affinity with a lower Caradocian radiolarian assemblage reported by Wang (Reference Wang, Blueford and Murchey1993) from the Pingliang Formation in Gansu Province, China. During this investigation, we examined samples collected from outcrops of the Crawford Group in both the Hawkwood Burn and the Leadhills. We discuss revised systematics for the taxa we recovered, introduce a new species with the aid of microcomputed tomography (micro-CT) and recognize six additional forms that have not previously been described from the Southern Uplands, but are known from Middle and Upper Ordovician localities elsewhere.
2. Regional setting
The Southern Uplands terrane is a classic example of a palaeo-subduction complex in which numerous thrust slices of accreted ocean-floor rocks succeeded by trench-fill turbidites are preserved (Leggett et al. Reference Leggett, McKerrow and Eales1979). Lower Palaeozoic strata in the Southern Uplands include multiple individual fault-bounded slivers containing successions of ocean-floor basalts, radiolarian cherts, black shales and turbidites similar to those now commonly recognized elsewhere as relict representatives of an ocean plate stratigraphy (OPS) (Isozaki et al. Reference Isozaki, Maruyama and Furuoka1990). Collectively, these rocks represent a classic subduction complex that developed as Iapetean oceanic lithosphere was subducted under a continental arc (represented by the Grampian Highlands) that developed along the SE margin of Laurentia (Leggett et al. Reference Leggett, McKerrow and Eales1979, Reference Leggett, McKerrow, Casey and Leggett1982; Stone et al. Reference Stone, McMillan, Floyd, Barnes and Phillips2012; Fujisaki et al. Reference Fujisaki, Asanuma, Suzuki, Sawaki, Sakata, Hirata, Maruyama and Windley2015).
The middle Arenigian – lower Caradocian Crawford Group represents the oldest Ordovician strata known from the Southern Uplands terrane (Stone et al. Reference Stone, McMillan, Floyd, Barnes and Phillips2012). It incorporates the Raven Gill and Kirkton formations that include numerous layers of bedded chert, mudstone and pillow basalt. The Kirkton Formation is succeeded by the Moffat Shale Group, which is dominated by black shale and grey mudstone and spans the remainder of the Caradocian strata. Above the Moffat Shale, a thick succession of turbiditic sandstones extends up to the lower Silurian System.
3. Materials and methods
The faunas reported here were extracted from red-brown ribbon-bedded chert samples collected from the Hawkwood Burn (55° 30′ 15″ N, 3° 37′ 34″ W WGS84; Fig. 1b) and grey ribbon-bedded chert samples from the Leadhills (55° 26′ 03″ N, 3° 46′ 15″ W WGS84; Fig. 1c). Both localities lie among Ordovician strata of the accretionary complex. Cherts from both the Hawkwood Burn and Leadhills areas have previously been examined for radiolarians (Danelian, Reference Danelian1999; Danelian & Floyd, Reference Danelian and Floyd2001; Kakuwa & Floyd, Reference Kakukwa and Floyd2017).
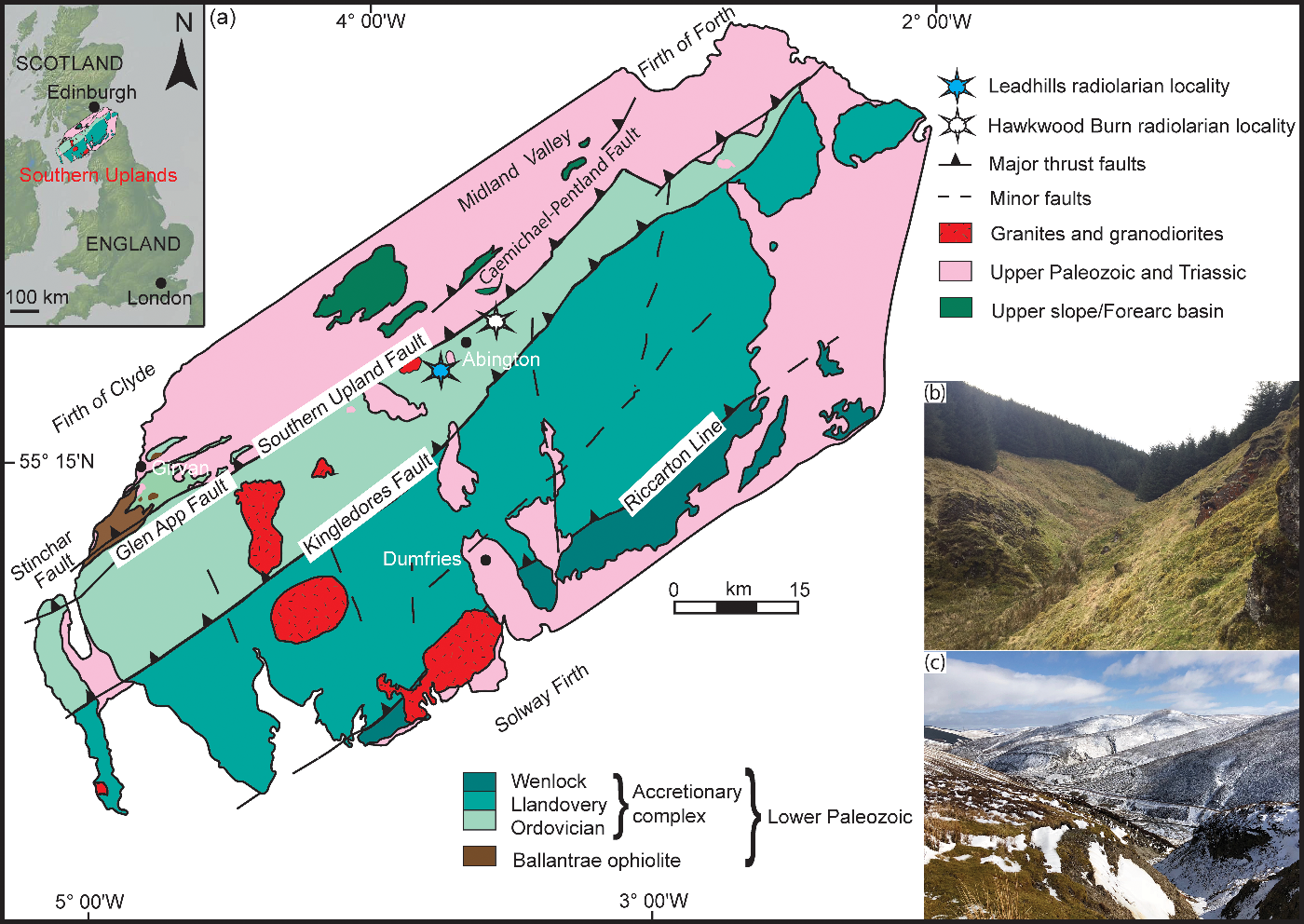
Fig. 1. (a) Geological map of the Southern Uplands terrane, modified after Leggett et al. (Reference Leggett, McKerrow and Eales1979, Reference Leggett, McKerrow, Casey and Leggett1982); inset map shows the Southern Uplands region in relation to the UK. (b) Field photograph of Hawkwood Burn area viewed looking upstream (to the south). (c) Field photograph of Gripps Cleugh, Leadhills viewed looking downstream (to the NE).
Samples were fragmented into approximately 5 cm3 pieces and chemically digested using standard hydrofluoric acid leaching techniques (Pessagno & Newport, Reference Pessagno and Newport1972) in the micropalaeontology laboratory of the School of Earth and Environmental Sciences (SEES), The University of Queensland. The acid residues were wet-sieved after 24 hours, and the fractions between 250 µm and 63 µm were collected and air dried. The procedure was repeated several times until a representative assemblage of radiolarians was recovered. Well-preserved specimens were individually picked under a Leica M80 light microscope and later imaged using a desktop Hitachi TM3030 SEM.
One specimen with an uncertain taxonomic affiliation, here described as Haplotaeniatum albaensis, was examined using the Zeiss Xradia Versa XRM-500 high-resolution scanner at the Julius Kruttschnitt Mineral Research Centre, The University of Queensland, following the method developed by Kachovich et al. (Reference Kachovich, Sheng and Aitchison2019). Prior to scanning, machine parameters and configuration requirements were set according to the standard set-up developed by Kinney & Nichols (Reference Kinney and Nichols1992). In order to obtain an optimum exposure and maximum contrast, the specimens were scanned for 4 hours under a beam voltage of 50 keV and X-ray source current of 79 μA. A stack of 969 images of 0.63 µm voxel size was produced for the specimen. Volume reconstruction software built into the Xradia 500 was used to convert the reconstructed volumetric data into 32-bit z-stacks, which were imported as TIFF images into Avizo® 9.7 software for further processing. Avizo volume rendering software was used to generate a three-dimensional model of the specimen. Before segmentation, the specimen was carefully separated from the adhesive using density filters. The cleaned specimen was then manually dissected into separate whorls. To better understand the relationship of spines to the whorls and proloculus, each spine was manually isolated by removing the attached meshwork until the point of origin was surfaced. All figured specimens are deposited in the micropalaeontology collection of the SEES, University of Queensland, Australia. SEES/170403 refers to the collection number, while the middle two digits and the last three characters of the specimen identification number (SEES/170403-XX-YYY) correspond to the sample number and species identification number, respectively.
4. Biostratigraphic discussion
Cherts from the Hawkwood Burn yield a diverse Middle Ordovician radiolarian assemblage. The majority of samples produced a moderately well-preserved radiolarian assemblage with potential for use in biostratigraphy. Samples SEES/170403-01 and SEES/170403-04 exhibited higher species diversity. The assemblage is dominated by spumellarians, with a subordinate population of entactinarians. The most abundant species is Haplotaeniatum albaensis sp. nov., while the rest of the assemblage includes: Proventocitum procerulum, Inanibigutta sp. cf. I. verrucula, Inanibigutta sp. cf. I. pinglianensis, ?Inanibigutta inconstans, Inanigutta sp. cf. I. complanata, Inanigutta gansuensis, ?Inanihella penrosei, ?Oriundogutta ramificans, Haplentactinia armillata, Syntagentactinia sp. and a few other undistinguishable forms of Inaniguttidae affinity. Radiolarians extracted from the Leadhills area are poorly preserved and entirely dominated by spherical forms.
The Inaniguttidae characterize the majority of the Hawkwood Burn fauna described here. This is also true for all other assemblages previously reported from the Southern Uplands. Overall, the assemblage is similar to part of the fauna described from eastern Kazakhstan and Pingliang Formation, China (Nazarov & Popov, Reference Nazarov and Popov1980; Wang, Reference Wang, Blueford and Murchey1993). The dominant taxon is similar to a form in the fauna illustrated by Hinde (Reference Hinde1890) that he referred to as Haliomma vetustum, the present status of which is regarded as nomina dubia (Aitchison et al. Reference Aitchison, O’Dogherty and Suzuki2017 a). The Hawkwood Burn fauna includes at least seven species present in both the Haplentactinia armillata – Proventocitum procerulum and Haplentactinia juncta – Inanigutta unica assemblages of the biozonation established by Nazarov & Ormiston (Reference Nazarov, Ormiston, Blueford and Murchey1993). The ‘Haplentactinia armillata – Proventocitum procerulum’ assemblage was later modified as ‘Proventocitum procerulum’ in Aitchison et al. (Reference Aitchison, Suzuki, Caridroit, Danelian and Noble2017 b).
The conodonts Pygodus anserinus and Periodon aculeatus constrain the age of the chert sequences in the Hawkwood Burn in the Kirkton Formation to the latest Darriwilian – earliest Sandbian (late Llanvirnian – early Caradoc in terms of British series) (Armstrong et al. Reference Armstrong, Clarkson and Owen1990, Reference Armstrong, Floyd, Tingqing and Barron2002; Danelian & Floyd, Reference Danelian and Floyd2001). In addition, based on a poorly preserved, less diverse radiolarian assemblage, Danelian (Reference Danelian1999) tentatively correlated the same chert sequences with the Haplentactinia juncta – Inanigutta unica zone which was described from eastern Kazakhstan. The assemblage reported here can be placed within the P. anserinus conodont zone with confidence, as the fauna described comes from strata in the same area that were previously described by Armstrong et al. (Reference Armstrong, Clarkson and Owen1990, Reference Armstrong, Floyd, Tingqing and Barron2002) and Danelian (Reference Danelian1999) (Fig. 2a). This suggests the possibility of extending the stratigraphic range of Proventocitum procerulum assemblage (Aitchison et al. Reference Aitchison, Suzuki, Caridroit, Danelian and Noble2017 b) until at least to the end of the Darriwilian Age since the last appearance datum of P. procerulum and H. armillata can be extended to the upper boundary of the P. anserinus biozone (Fig. 3).
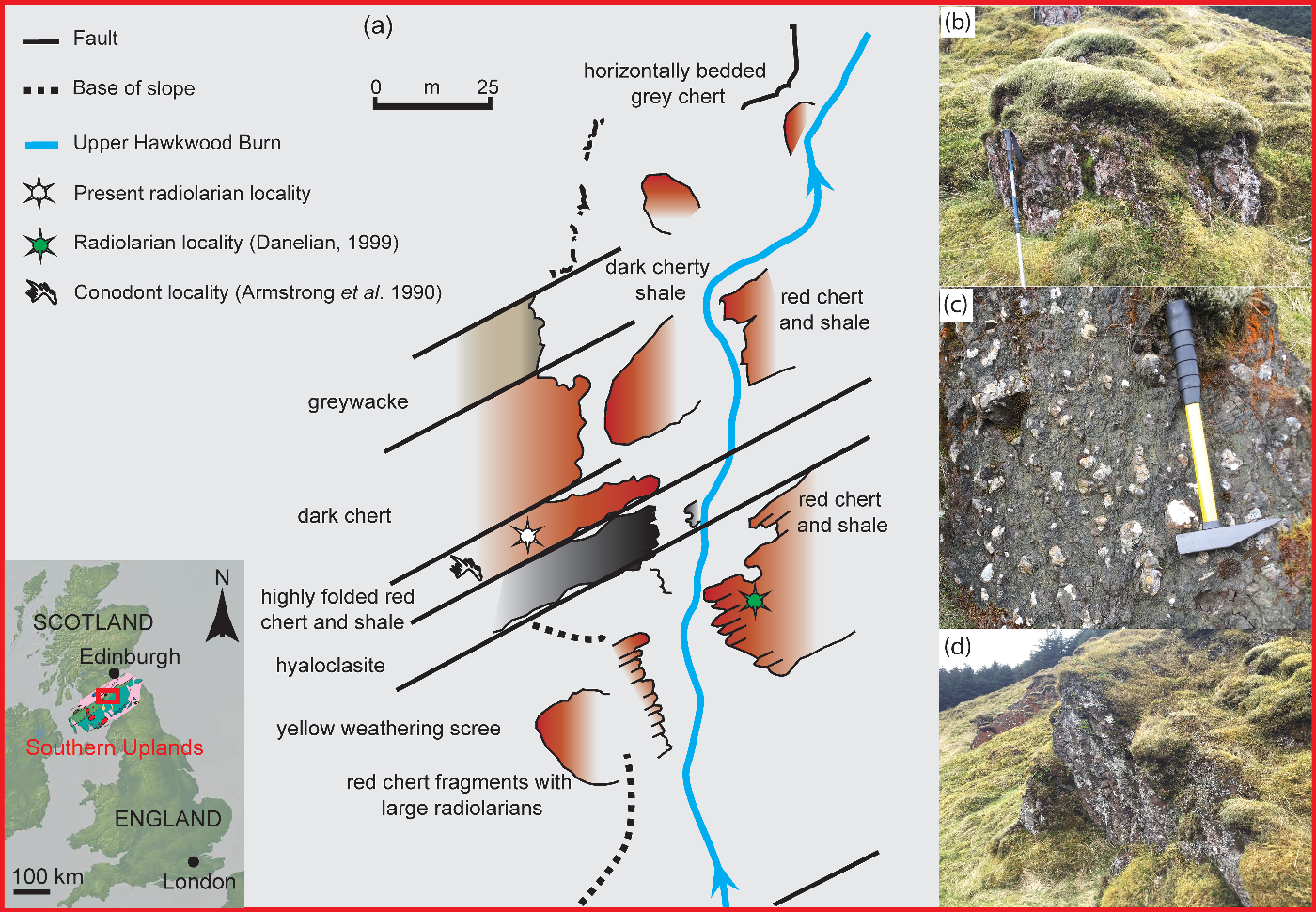
Fig. 2. (a) Sketch map of the geology of Hawkwood Burn modified after Armstrong et al. (Reference Armstrong, Clarkson and Owen1990); (b) field photograph of cherts from Hawkwood Burn; (c) hyaloclastite layer at Hawkwood Burn; and (d) the chert outcrop, which yielded the best-preserved radiolarian assemblage.
5. Systematic palaeontology
Phylum RETARIA Cavalier-Smith, Reference Cavalier-Smith1999, emend. Cavalier-Smith, Reference Cavalier-Smith2002
Subphylum RADIOZOA Cavalier-Smith, Reference Cavalier-Smith1987
Superorder POLYCYSTINA Ehrenberg, Reference Ehrenberg1838, emend. Riedel, Reference Riedel1967
Order ENTACTINARIA Kozur & Mostler, Reference Kozur and Mostler1982
Family HAPLENTACTINIIDAE Nazarov in Nazarov & Popov, Reference Nazarov and Popov1980
Genus Haplentactinia Foreman, Reference Foreman1963
Type species. Haplentactinia rhinophyusa Foreman, Reference Foreman1963
Haplentactinia armillata Nazarov & Popov, Reference Nazarov and Popov1980
Figure 4g, j
1980 Haplentactinia armillata Nazarov in Nazarov & Popov, p. 55, pl. 16, figs 9–10, text-fig. 31
1988 Haplentactinia armillata; Nazarov, pl. IX, fig. 12
1998 Haplentactinia armillata; Aitchison, p. 79, pl. 1, fig. U
Holotype. Specimen No. 4333/58 in the Geological Institute, Moscow. Known from the Lower–Middle(?) Ordovician, Arenigian–Llanvirnian(?), Kogashsky Horizon, Kysylzharskaya Suite, Central Kazakhstan, near Sary-Kum Station.
Description. Comprised a sub-spherical spongy outer shell with a diameter (Ø) of 280–300 µm. Primary spines differ from each other in diameter, length and the morphology of the distal end. One of the primary spines show a strong bifurcation, while the rest taper towards the distal end. Tapering primary spines show an average length of 155 µm. The bifurcated spine gradually tapers from Ø 15–20 µm at its base and culminates at pointed distal ends. Numerous short by-spines extend from the shell.
Remarks. Nazarov & Popov (Reference Nazarov and Popov1980) originally placed this form with Haplentactinia as the thick spongy layer obstructed observation of a microsphere.
Materials. Seven specimens identified from sample SEES/170403-01 and SEES/170403-04 collected from Hawkwood Burn, Southern Uplands terrane, Scotland.
Genus Syntagentactinia Nazarov in Nazarov & Popov, Reference Nazarov and Popov1980
Type species. Syntagentactinia biocculosa Nazarov
Syntagentactinia sp.
Figure 4b
Description. Flattened test comprised two polygonal-shaped lattices, developed around at least six needle-like slender spines that originate eccentrically. The lattice is woven around the main spines that vary in length (100–130 µm).
Remarks. The fragmentary and poor preservation of the specimen is such that it is not possible to assign the specimen to any established species of the genus with confidence. Although the spine arrangement is closely related to Haplentactinia juncta, the presence of two distinguishable lattices places it within Syntagentactinia.
Materials. Single specimen identified from sample SEES/170403-02 collected from Hawkwood Burn, Southern Uplands terrane, Scotland.
Order SPUMELLARIA, Ehrenberg, Reference Ehrenberg1876
Family HAPLOTAENIATIDAE Won et al. Reference Won, Blodgett and Nestor2002
Genus Haplotaeniatum Nazarov & Ormiston, Reference Nazarov, Ormiston, Blueford and Murchey1993
Type species. Haplotaeniatum labyrintheum Nazarov & Ormiston, Reference Nazarov, Ormiston, Blueford and Murchey1993
Diagnosis. ‘Spherical skeleton consisting of small proloculus overlain by several weakly to discontinuously spiral labyrinthine layers. Commonly pylomate, with or without major outer spines’ (Jones & Noble, Reference Jones and Noble2006).
Remarks. Revision of the diagnosis of this genus by Jones & Noble (Reference Jones and Noble2006) resulted in the removal of forms with distinct medullary and cortical areas and restricted the genus to forms showing weak labyrinthine to discontinuous spiraliform layering. However, the configuration of layering may appear contrasting in different orientations and is dependent on how the specimen has been sectioned. The original description by Nazarov & Ormiston (Reference Nazarov, Ormiston, Blueford and Murchey1993) included no details about a pylome, although later revisions suggested the possible presence of a pylome with or without a margin. However, when a pylome without a margin is present, it is not readily visible and it is challenging to distinguish it from irregular-shaped pores in the labyrinthine meshwork.
Haplotaeniatum albaensis sp. nov.
Figure 4e, k
1890 Haliomma vetustum Hinde in Hinde, p. 51, pl, 3, fig. 11 (nomen dubium)
Etymology. Named after the Scottish Gaelic name for Scotland ‘Alba’.
Holotype. Specimen no. SEES/170403-04-HAL1 (Fig. 4e) in the micropalaeontology collection of SEES, University of Queensland, Australia. Known from red ribbon-bedded chert of the Crawford Group in the Hawkwood Burn, Southern Uplands terrane, Scotland.
Paratypes. Specimen nos SEES/170403-04-HAL2 (Fig. 4k) and SEES/170403-04-HAL3 (Fig. 5 and online Supplementary video S1, available at http://journals.cambridge.org/geo) from red ribbon-bedded chert at the type locality.
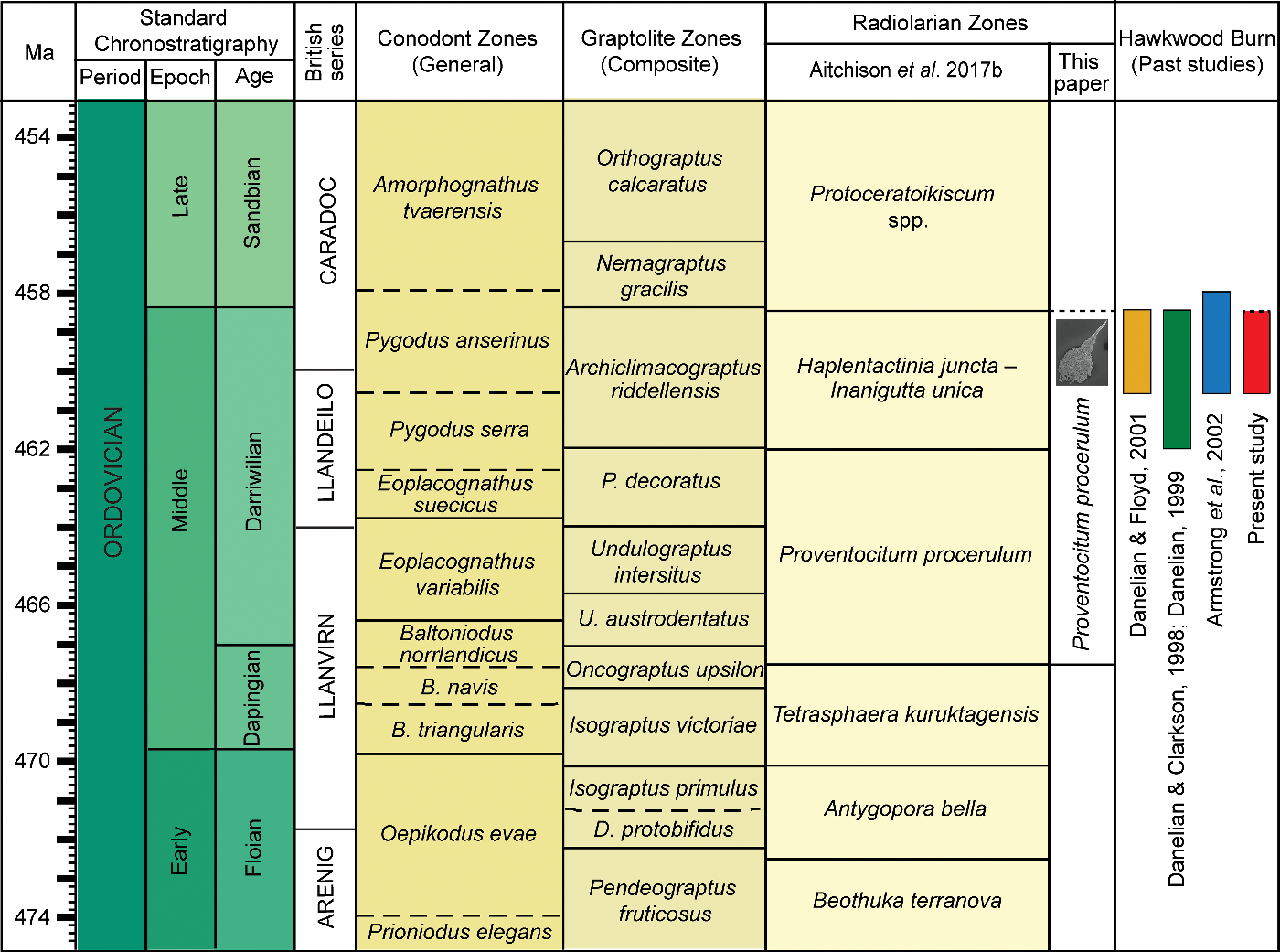
Fig. 3. Correlation of Ordovician microfossil zones and biostratigraphic data for Hawkwood Burn, Southern Uplands terrane. Calibration to geological time scale of Ogg et al. (Reference Ogg, Ogg and Gradstein2016) performed using TS Creator (https://timescalecreator.org/index/index.php).

Fig. 4. SEM images of radiolarians recovered from the Crawford Group at the Hawkwood Burn locality, Southern Uplands terrane, Scotland. Scale bar: 100 µm. (a) Proventocitum procerulum Nazarov, SEES/170403-01-PPR1. (b) Syntagentactinia sp. SEES/170403-02-SSP1. (c) ?Inanihella penrosei (Ruedemann & Wilson), SEES/170403-01-IPE1. (d) Inanigutta gansuensis Wang, SEES/170403-01-IGA1. (e) Haplotaeniatum albaensis sp. nov. SEES/170403-04-HAL1. (f) Inanibigutta sp. cf. I. pinglianensis Wang, SEES/170403-04-IPI1. (g) Haplentactinia armillata (Nazarov), SEES/170403-01-HAR1. (h) Inanibigutta sp. cf. I. verrucula Nazarov, SEES/170403-01-IVE1. (i) Inanigutta sp. cf. I. complanata Nazarov, SEES/170403-05-ICO1. (j) Haplentactinia armillata (Nazarov), SEES/170403-04-HAR2. (k) Haplotaeniatum albaensis sp. nov. SEES/170403-04-HAL2. (l) Inaniguttid gen. et sp. indet. SEES/170403-05-ISP1. (m) ?Oriundogutta ramificans Nazarov, SEES/170403-01-ORA1. (n) Inaniguttid gen. et sp. indet. SEES/170403-05-ISP2. (o) ?Inanibigutta inconstans Nazarov, SEES/170403-01-IIN1. (p) Light micrograph and (q) SEM image of Inaniguttid gen. et sp. indet. SEES/170403-02-ISP3.

Fig. 5. Reconstruction of possible ontogenetic growth stages from a micro-CT model of Haplotaeniatum albaensis sp. nov. from chert sequences in Hawkwood Burn, Scotland. All images were digitally processed using Avizo® software: (a) separated whorls; (b) cross-section showing proloculus and whorls in labyrinthine arrangement; (c) whorls connected through bars at multiple points; (d) bandaged outer mesh; (e) small pores; (f) branched apophyses; and (g) primary spines starting from proloculus. Scale bar: (a, b, f, g) 100 µm; and (c, d, e) 25 µm.
Diagnosis. Sub-spherical skeleton consisting of a proloculus overlain by three weakly discontinuous labyrinthine whorls forming a dense meshwork. Outermost whorl constructed from bandage-like mesh, while the rest of the skeleton is formed of rods with irregular thicknesses. At least 12 conical spines protrude from surface of the outermost whorl, out of which seven spines originate from the proloculus.
Description. When investigated using micro-CT the sub-spherical test of the H. albaensis sp. nov. can be seen to be constructed of three weakly discontinuous labyrinthine whorls growing around a centrally located proloculus (Ø, 70–80 µm) (see Fig. 5a). The lattice of the proloculus shows irregularly sized and shaped pores. There are numerous short connecting bars of variable thicknesses between the first whorl and the proloculus and between the three whorls (see Fig. 5c). Each whorl grows up to 30 µm in thickness and maintains an average distance of 10 µm between the proloculus and adjacent whorl at places with no connecting bars. The nature of the mesh of the outer whorl is bandage-like (see Fig. 5d), while the rest of the skeleton is formed of thin bars of irregular thicknesses. Randomly positioned circular pores (Ø c. 4 µm) (see Fig. 5e) can be seen on bandages and bars of the mesh. Seven primary spines that are straight or slightly curved originate directly from the proloculus (see Fig. 5g) and pass through whorls to protrude from the outer surface. Randomly distributed apophyses branch from the primary spines to develop arches on the surface of the shell (see Fig. 5f). By-spines arbitrarily initiate from three whorls. The average diameter of the base of any spines is c. 20 µm. Primary spines show a maximum length of 55 µm from the outer surface of the shell.
Remarks. More than ten specimens identified as H. albaensis sp. nov. were examined by SEM; however, their internal structure was only revealed by micro-CT observation. Hinde (Reference Hinde1890) documented a fragmentary specimen with a ‘thick cortical shell and a medullary shell bearing nine spines on the exposed surface’ that may be similar, but is regarded as nomina dubia. The diameters of the cortical shell and the medullary shell of Haliomma vetustum Hinde are approximate to the diameter of the entire shell and the proloculus of our material. Use of the term ‘proloculus’ is adopted here for the innermost shell after considering the definition of Won & Iams (Reference Won and Iams2011), which states ‘microsphere differs from the proloculus by its complete separation from surrounding shell wall’. The three whorls arranged in the labyrinthine configuration are hardly separable in a physically segmented specimen. However, micro-CT observation of a perpendicular slice through an equatorial plane reveals that the dense mesh can be readily separated into three whorls. Without the aid of a micro-CT model, it is very challenging to differentiate primary and secondary spines. This species also bears some superficial similarity with Beothuka echinata Won & Iams (Reference Won and Iams2013) as a result of the incomplete layering of shells, but differs greatly as the proloculus and the rest of the shell of B. echinata demonstrates a strongly bandaged appearance and lack of labyrinthine configuration. The inaniguttid genus Triplococcus also shares three shells, but is restricted to closely spaced perfectly perforated spheres connected with five to six rod-like beams.
Materials. Abundant in samples collected from red ribbon-bedded chert of the Crawford Group in Hawkwood Burn, Southern Uplands terrane, Scotland.
Age and distribution. Middle Ordovician (Darriwilian), Hawkwood Burn, Crawford Group, Southern Uplands terrane, Scotland, so far as is known.
Family INANIGUTTIDAE Nazarov & Ormiston, Reference Nazarov, Ormiston, Petrushevskaya and Stepanjants1984 sensu Danelian & Popov, Reference Danelian and Popov2003
Remarks. Genus level classification within this family requires a thorough understanding of structural features such as the number of spheres and their individual dimensions, together with the exact number of spines. However, since the establishment of this family by Nazarov & Ormiston (Reference Nazarov, Ormiston, Petrushevskaya and Stepanjants1984), well-preserved specimens of the type necessary for genus- and species-level diagnosis have rarely been described. Type materials are commonly vaguely sketched or poorly illustrated within distinct transmitted light microscope images. Multiple studies have questioned the validity of the number of spheres and spines as a diagnostic feature, and some researchers claim that the presence or absence of these features relate to ontogeny (Noble & Aitchison, Reference Noble, Aitchison, Blome, Whalen and Reed1995; Danelian & Popov, Reference Danelian and Popov2003; Suzuki, Reference Suzuki2006; Maletz, Reference Maletz2011; S Kachovich, unpub. PhD thesis, The University of Queensland, 2018; Kachovich & Aitchison, Reference Kachovich and Aitchison2020). This is evident from the variations in the shell dimensions and numbers of spines reported in many of the studies that have reported inaniguttids (Nazarov & Popov, Reference Nazarov and Popov1980; Nazarov & Ormiston, Reference Nazarov, Ormiston, Petrushevskaya and Stepanjants1984, Reference Nazarov, Ormiston, Blueford and Murchey1993; Nazarov, Reference Nazarov1988; Wang, Reference Wang, Blueford and Murchey1993; Li, Reference Li1995; Danelian & Clarkson, Reference Danelian and Clarkson1998; Danelian, Reference Danelian1999; Danelian & Floyd, Reference Danelian and Floyd2001; Noble & Webby, Reference Noble and Webby2009; Pouille et al. Reference Pouille, Danelian, Ghobadi and Popov2013). Most specimens described as inaniguttids are tentatively assigned at either genus or species level because it is not always possible to clearly determine the presence or otherwise of medullary shells.
Genus Inanibigutta Nazarov & Ormiston in Nazarov, Reference Nazarov1988
Type species. Entactinosphaera aksakensis (Nazarov) Reference Nazarov1975
?Inanibigutta inconstans (Nazarov), Reference Nazarov1975
Figure 4o
1975 Entactinosphaera inconstans Nazarov, p. 69, pl. 17, figs 2–4, pl. 21, fig. 1
1980 Entactinosphaera inconstans; Nazarov & Popov, p. 36, pl. 3, fig. 7, pl. 11, fig. 7, pl. 13, fig. 6
Holotype. Specimen no. 4333/31 in the Geological Institute, Moscow. Known from the Middle Ordovician, Nograd Horizon, Bestamak Formation, eastern Kazakhstan, Chingiz.
Description. Spherical outer shell is perforated by sub-rounded pores up to Ø 10 µm. Two straight rod-like primary spines of c. 100 µm length extend from the outer shell, while traces of two other primary spines can be noted. Numerous short by-spines protrude from pore–bar junctions. The average width of the base of primary spines is 16 µm and they taper distally. The outer shell has Ø c. 225 µm and the presence of a medullary shell can be noted, despite the poor preservation and fragmentary nature.
Remarks. This species is tentatively assigned to Inanibigutta inconstans as the inner part of its test is ambiguous and the genus-level assignment is uncertain. Our specimen superficially resembles Nazarov’s type material, although it is slightly smaller in size.
Materials. Five specimens from SEES/170403-01 in Hawkwood Burn, Southern Uplands terrane, Scotland.
Inanibigutta sp. cf. I. pinglianensis Wang, Reference Wang, Blueford and Murchey1993
Figure 4f
1993 Inanibigutta pinglianensis Wang, p. 100, pl. 5, fig. 2
Holotype. Inanibigutta pinglianensis Wang, Reference Wang, Blueford and Murchey1993. Specimen cat. no. R0066 in Nanjing Institute of Geology & Palaeontology, Academia Sinica, China. Known from Middle Ordovician, Pingliang Formation, Gansu Province, China.
Description. Sub-spherical to spherical outer sphere with average Ø of 220 µm. Rod-like primary spines develop significant apophyses and taper towards their distal end. Cone-shaped secondary spines may be developed at pore–bar junctions and rarely display bifurcated ends. Dimensions of the oval to sub-spherical pores are variable.
Remarks. The outer spheres of specimens (Ø, 220–360 µm) among the type material from the Pingliang Formation are commonly larger than those of the specimens (Ø, 210–230 µm) found in this study. However, our specimens exhibit similar characteristics among their primary and secondary spines. The inner sphere is less evident due to the state of preservation. The diameters of the specimens found in this study approximate that of the outer sphere of I. verrucula. Inanibigutta sp. cf. I. pinglianensis can be distinguished as a result of tapering primary spines and bifurcate apophyses on primary as well as on secondary spines. Due to uncertainty at the species level, we tentatively identify specimens as Inanibigutta sp. cf. I. pinglianensis.
Materials. Tens of specimens from SEES/170403-01 and SEES/170403-04 in Hawkwood Burn, Southern Uplands terrane, Scotland.
Inanibigutta sp. cf. I. verrucula (Nazarov & Popov) Reference Nazarov and Popov1976
Figure 4h
1976 Entactinosphaera verrucula Nazarov in Nazarov & Popov, p. 408–409, fig. 1d
1980 Entactinosphaera verrucula; Nazarov & Popov, p. 38, pl. 3, fig. 6, pl. 11, fig. 6, text-fig. 17
1988 Inanibigutta verrucula; Nazarov, fig. 31
1993 Inanibigutta verrucula; Wang, p. 100, pl. 6, figs 2–3, 5–8
1995 Inanibigutta verrucula; Li, p. 90, pl. 3, fig. 9
1998 Inanibigutta verrucula; Danelian & Clarkson, p. 135, fig. 2g
1999 Inanibigutta sp. cf. I. verrucula; Danelian, p. 630, fig. 4H
2001 Inanibigutta (?) sp. O; Danelian & Floyd, p. 493, fig. 4b
Holotype. Entactinosphaera verrucula Nazarov, Reference Nazarov and Popov1976. Specimen No. GIN 4333/18 in the Geological Institute, Moscow. Known from the Middle Ordovician, Bestamak Formation, Southwestern foothills of Chingiz range, eastern Kazakhstan.
Description. Diameter of outer shell in the range 195–210 µm. It is perforated by uniformly arranged rounded pores of equal size. Principal spines are of consistent diameter along their length and arranged at near-right angles to one another. Apophyses are more concentrated on the proximal ends of the principal spines. Not all the principal spines are visible on any given specimen because of poor preservation. Short, slender and pointed secondary spines are located at pore–bar junctions.
Remarks. The uniform diameter maintained by the rod-like primary spines for most of their length makes this species readily distinguishable from other inaniguttids. Although Nazarov & Popov (Reference Nazarov and Popov1980) emphasized the combination of one long and five shorter principal spines for their diagnosis, all the encountered specimens are fragmentary and that feature cannot be relied upon for identification. In addition, several authors have reported different dimensions for the outer sphere of Inanibigutta verrucula leading to uncertainty among identification of specimens. (I. verrucula outer sphere Ø, 160–200 μm in Nazarov & Popov, Reference Nazarov and Popov1976, Reference Nazarov and Popov1980; 131 μm in Danelian & Clarkson, Reference Danelian and Clarkson1998; 180–260 μm in Wang, Reference Wang, Blueford and Murchey1993). Our specimens are tentatively assigned to this species based on the diameter of the cortical shell and the apparently uniform thickness of the principal spine.
Materials. Tens of specimens from SEES/170403-01, SEES/170403-04 in Hawkwood Burn, Southern Uplands terrane, Scotland.
Genus Inanigutta Nazarov & Ormiston, Reference Nazarov, Ormiston, Petrushevskaya and Stepanjants1984
Type species. Entactinia unica (Nazarov) Reference Nazarov1975
Inanigutta sp. cf. I. complanata (Nazarov) Reference Nazarov1975
Figure 4i
1975 Entactinia complanata Nazarov, p. 56, pl. XV, figs 11–12, pl. XX, figs 7–8
1980 Entactinia complanata; Nazarov & Popov, p. 29, pl. 1, figs 2, 5, pl. 7, figs 3, 4, pl. 11, figs 3, 4, text-fig. 10
1984 Inanigutta complanata; Nazarov & Ormiston, pl. IV, fig. 1
1993 Inanigutta complanata; Wang, p. 99, pl. 10, figs 1, 2
1992 Inanigutta cf. I. complanata; Goto et al., p. 159, 160, pl. 9, figs 1–3
2001 ?Inanigutta complanata; Danelian & Floyd, p. 493, fig. 4a
2009 Inanigutta complanata; Noble & Webby, pl. 5, figs 10–11, pl. 6, fig. 14
Holotype. Entactinia complanata (Nazarov) Reference Nazarov1975. Specimen No. GIN 4333/29 in the Geological Institute, Moscow. Known from the Middle Ordovician, Bestamak Formation, Southwestern foothills of Chingiz range, eastern Kazakhstan.
Description. Spherical skeleton with an irregularly porous cortical shell having average Ø of 215 µm. Length of the major spines is c. 240 µm. Bears numerous short by-spines and two very long primary spines, which taper gradually towards their distal end. Primary spines are commonly slightly curved and apophyses can be seen. Signs of medullary shells are not visible due to the poor preservation.
Remarks. Precise identification at species level is challenging given the state of preservation. The dimensions of the cortical shell and the length of the major spines bear close similarity with Entactinia complanata Nazarov as described in Nazarov & Popov (Reference Nazarov and Popov1980) and Nazarov & Ormiston (Reference Nazarov, Ormiston, Petrushevskaya and Stepanjants1984). However, subsequent contradictory descriptions of I. complanata cast doubt on the exact dimensions and arrangement of the internal skeleton (Wang, Reference Wang, Blueford and Murchey1993; Danelian & Floyd, Reference Danelian and Floyd2001; Noble & Webby, Reference Noble and Webby2009).
Materials. Tens of specimens from SEES/170403-01 and SEES/170403-04 of the Hawkwood Burn, Crawford Group, Southern Uplands terrane, Scotland.
Inanigutta gansuensis Wang, Reference Wang, Blueford and Murchey1993
Figure 4d
1993 Inanigutta gansuensis Wang, pl. 7, fig. 1–8, pl. 8, fig. 1–11
2013 Inanigutta gansuensis; Pouille et al. Reference Pouille, Danelian, Ghobadi and Popov2013, fig. 6.6
Holotype. Inanigutta gansuensis Wang, Reference Wang, Blueford and Murchey1993. Specimen cat. no. R0087 in Nanjing Institute of Geology & Palaeontology, Academia Sinica, China. Known from Middle Ordovician, Pingliang Formation, Gansu Province, China.
Description. Spherical shell, Ø c. 180 µm with six primary spines, which taper towards their end. Primary spines bear apophyses. Pores are rounded to oval in shape with a polygonal ridge surrounding them.
Remarks. Spines on the specimens in our material are commonly broken and shells are in-filled with secondary minerals, hampering observation. The shell diameter along with the structure and arrangement of spines suggests assignment to I. gansuensis.
Materials. Tens of specimens from SEES/170403-01 and SEES/170403-04 in Hawkwood Burn, Southern Uplands terrane, Scotland.
Genus Inanihella Nazarov & Ormiston, Reference Nazarov, Ormiston, Petrushevskaya and Stepanjants1984
Type species. Helioentactinia bakanasensis (Nazarov) Reference Nazarov1975
?Inanihella penrosei (Ruedemann & Wilson) Reference Ruedemann and Wilson1936
Figure 4c
1980 Helioentactinia penrosei; Nazarov & Popov, p. 50, pl. 4, fig. 6, pl. 13, figs 1–2
1993 Inanihella penrosei; Wang, p. 102, pl. 1, figs 1–6, pl. 2, figs 1–9, pl. 3, figs 1–3, pl. 4, figs 1–3
2008 Inanigutta penrosei group; Maletz & Bruton, p. 1193, pl. 8, figs. H, I, N
Holotype. Neotype Specimen no. 4333/41 in the Geological Institute, Moscow (see Helioentactinia penrosei; Nazarov & Popov Reference Nazarov and Popov1980, pl. 4, fig. 6)
Description. Sub-spherical porous shell with more than six long straight primary spines gradually tapering towards distal ends. Outer shell diameter averages 200 µm. Length of primary spines exceeds the diameter of the outer shell and they rarely have apophyses. Short, secondary spines developed along pore–bar junctions. Traces of a medullary sphere can be seen through the fragmented surface.
Remarks. Although Wang (Reference Wang, Blueford and Murchey1993) considered Haliomma penrosei Ruedemann & Wilson as a synonymy for I. penrosei, the vaguely sketched specimens in Ruedemann & Wilson (Reference Ruedemann and Wilson1936) do not provide an adequate basis for comparison. In addition, Haliomma Ehrenburg refers to an extant genus. Therefore, Haliomma penrosei is now regarded as a nomen dubium and no longer valid as a synonymy. Accordingly, Helioentactinia penrosei, illustrated in Nazarov & Popov (Reference Nazarov and Popov1980), can be validly established as the neotype. Although Wang (Reference Wang, Blueford and Murchey1993) describes this taxon as having ‘numerous major spines’, his specimens appear to have no more than eight major spines. Additionally, the diameter of the outer sphere documented by Nazarov & Popov, Reference Nazarov and Popov1980 (Ø, 280–340 µm) and Wang, Reference Wang, Blueford and Murchey1993 (Ø, 240–440 µm) is significantly larger than the outer sphere diameter reported here. It is therefore not possible to assign the encountered specimen to I. penrosei with confidence, although it bears a superficial resemblance. Noble & Aitchison (Reference Noble and Aitchison2000) and Maletz & Bruton (Reference Maletz and Bruton2008) adopted the term ‘Inanigutta penrosei group’, which aggregates together many long-spined inaniguttids including Inanihella penrosei.
Materials. Four specimens from chert of the Crawford Group, Southern Uplands terrane, Scotland.
Genus Oriundogutta Nazarov & Ormiston in Nazarov, Reference Nazarov1988
Type species. Astroentactinia ramificans (Nazarov)
?Oriundogutta ramificans (Nazarov) Reference Nazarov1975
Figure 4m
1975 Astroentactinia ramificans Nazarov in Nazarov, 87, pl. 19, fig. 2–6, pl. 21, figs 3–5
1980 Astroentactinia ramificans; Nazarov & Popov, p. 43, pl. 3, figs 2–3, pl. 1, fig. 1, pl. 13, figs 4–5, text-fig. 21)
1988 Oriundogutta ramificans; Nazarov, p. 57, pl. 10, fig. 2
Holotype. Specimen No 4333/6 in the Geological Institute, Moscow. Known from the Middle Ordovician, Linograd Horizon, Bestamak Suite, east Kazakhstan, Chingiz.
Description. The shell is spherical with average Ø of 190 µm. Rod-like primary spines are longer than numerous by-spines protruding from the outer surface. Lengths of secondary spines range over 15–70 µm, whereas primary spines are fragmentary. The average width of the base of a primary spine is 20 µm. Rounded pores on the outer shell are of varying sizes with a maximum Ø of 18 µm. Inner structure is dubious and cannot be identified precisely as a result of the secondary infillings.
Remarks. Our specimens are slightly smaller in diameter and have shorter by-spines compared with the holotype. Since the internal structures of the specimens are difficult to resolve, making the genus-level characters indeterminate, the specimens are tentatively assigned to ?Oriundogutta ramificans.
Inaniguttid gen. et sp. indet.
Figure 4l, n, p, q
Remarks. Specimens are poorly preserved and features of the external shell are not visible as a result of secondary infillings and their fragmentary nature. However, the presence of tapering spines and signs of a sub-spherical test distinguish them from other families.
The specimen illustrated in Figure 4l bears strong resemblance to figure 8H illustrated in Danelian et al. (Reference Danelian, Popov, Tolmacheva, Pour, Neyevin and Mikolaichuk2011). Their figured specimen has three spines located on the same plane, of which two are in a nearly sub-parallel position and the third is at the opposite pole. Our specimen (Fig. 4l) shows one of the two sub-parallel primary spines with the third spine located in the opposite position. Danelian et al. (Reference Danelian, Popov, Tolmacheva, Pour, Neyevin and Mikolaichuk2011) placed their specimen illustrated in their fig. 8H in open nomenclature, as it was not clear whether this is an original arrangement or an effect of distortion. Since the same features are encountered in both specimens, we regard it as more likely to be an original feature. However, better-preserved material is required to establish this form as a species.
The specimen illustrated in Figure 4p, q can be compared with Dorysphaera reticulata Hinde (nomina dubia) figured by Hinde (Reference Hinde1890). Neither the preservation of our material nor Hinde’s description or illustration provide an ideal basis upon which to comment on the inner structure of the specimen. Although the specimens are not fragmentary, dense secondary infilling covers the details of the internal structure. Notwithstanding this, the taxon accounts for a considerable percentage of the total assemblage reported from this study.
Order INCERTAE SEDIS
Family PROVENTOCITIDAE Aitchison, Reference Aitchison1998
Genus Proventocitum Nazarov & Ormiston in Nazarov, Reference Nazarov1988; sensu Nazarov & Ormiston (Reference Nazarov, Ormiston, Blueford and Murchey1993)
Type species. Popofskyellum? procerulum Nazarov
Proventocitum procerulum Nazarov, Reference Nazarov1988
Figure 4a
1980 Popofskyellum? procerulum; Nazarov & Popov, p. 68–70, pl. XV, fig. 6
1988 Proventocitum procerulum Nazarov, p. 71, pl. IX, fig. 9, text-fig. 21b
1993 Proventocitum procerulum; Nazarov & Ormiston, p. 43, pl. 4, fig. 11, text-fig. 9b
1998 Proventocitum procerulum; Aitchison, p. 78, figs 3E, H, I
1998 Proventocitum sp. Aitchison; p. 76, figs 2H, J, K
Holotype. Specimen No. 4333/55 (1–2) in the Geological Institute, Moscow. Described from the Lower–Middle(?) Ordovician, Arenigian–Llanvirnian(?), Kogashsky Horizon, Kysylzharskaya Suite, Central Kazakhstan near Sary-Kum Station.
Description. Conical apical portion of the test is tri-segmented and rigid. Upper and intermediate segments are devoid of any pores or lamellar structure. Basal segment of the apical portion shows porous structure that continues to spread towards the spheroidal middle section of the shell. This section is approximately 230 µm in width and possesses at least two rod-like spines with apophyses. The length of these rod-like spines may vary, and they are directed downwards upon reaching their distal end. The length of the axial apical spine is approximately similar to the height of the apical portion. The distal end of the shell is open and the porous network appears to be hanging at the end.
Remarks. A single specimen was recovered from the Hawkwood Burn. It bears a remarkable resemblance to the holotype described by Nazarov in Nazarov & Popov (Reference Nazarov and Popov1980). Whether conical forms described in open nomenclature as Radiolaria familia nova spp. (Fortey & Holdsworth, Reference Fortey and Holdsworth1971) and all the specimens illustrated by Nazarov & Popov (Reference Nazarov and Popov1980), Nazarov (Reference Nazarov1988) and Nazarov & Ormiston (Reference Nazarov, Ormiston, Blueford and Murchey1993) can be assigned under P. procerulum is not clear because of imperfect preservation. Although the polarization of the skeleton of Proventocitum is considered by some researchers to be a characteristic feature of primitive nassellarians, this remains a subject of debate (Afanasieva et el. 2005; Maletz, Reference Maletz2011). We therefore follow Noble et al. (Reference Noble, Aitchison, Danelian, Dumitrica, Maletz, Suzuki, Cuvelier, Caridroit and O’Dogherty2017) and continue to regard the order-level assignment of the Proventocitidae as uncertain.
Materials. Single specimen identified from sample SEES/170403-01 collected from Hawkwood Burn, Southern Uplands terrane, Scotland.
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.1017/S001675682000028X
Acknowledgements
The research was financially supported by the Australian Research Council grant no. ARC DP 1501013325 (to JCA). We thank Dr Cathy Evans at the Julius Kruttschnitt Mineral Research Center, University of Queensland for conducting micro-CT measurements of the radiolarian specimens. Constructive remarks by reviewers Professor T. Danelian and Dr Paula Noble significantly improved the manuscript. This paper is a contribution to the International Geoscience Programme (IGCP) Project 653, The onset of the Great Ordovician Biodiversification Event.
Declaration of interest
None.








 Open access
Open access