



1. Introduction
The Iberian Chains (Cadenas Ibéricas) are two isolated NW–SE-striking mountain ranges of pre-Variscan rocks within the Central Iberian Ranges in NE Spain. The Western Iberian Chains are composed of rocks of Cambrian and Ordovician age, whereas the Eastern Iberian Chains expose rocks ranging from the Neoproterozoic era to the Carboniferous period. Tectonic deformation and faulting of these rocks occurred during the Variscan and Alpine orogenies as well as during postorogenic extensions (e.g. Carls et al. Reference Carls, Valenzuela-Ríos, Pocovi and Lago2002; Álvaro et al. Reference Álvaro, Bauluz, Gil-Imaz and Ubide2018). The Iberian Chains have been subdivided into three tectonostratigraphic units: the Badules unit which comprises the Western Iberian Chains and the southwestern part of the Eastern Iberian Chains; the Herrera unit which constitutes most of the northeastern part of the Eastern Iberian Chains; and the Mesones unit which is sandwiched between the two other units (Gozalo & Liñán, Reference Gozalo and Liñán1988; Álvaro et al. Reference Álvaro, Bauluz, Gil-Imaz and Ubide2018). The units are separated by two NW–SE-striking first-order structures, termed the Jarque and Datos thrust faults in the southwest and in the northeast, respectively (e.g. Gozalo & Liñán, Reference Gozalo and Liñán1988; Álvaro et al. Reference Álvaro, Bauluz, Gil-Imaz and Ubide2018; Fig. 1b). The Iberian Chains are widely considered as eastern extensions of two of the major tectonostratigraphic zones recognized in the Iberian Massif, i.e. the Cantabrian zone and the West Asturian Leonese zone (e.g. Gozalo & Liñán, Reference Gozalo and Liñán1988; Calvín-Ballester & Casas, Reference Calvín-Ballester, Casas, Llana-Fúnez, Marcos and Bastida2014; Álvaro et al. Reference Álvaro, Bauluz, Gil-Imaz and Ubide2018). Although the Jarque fault has traditionally been considered to separate these two zones because of structural and stratigraphic similarities between the Narcea antiform in the Cantabrian Mountains and the Paracuellos antiform in the Eastern Iberian Chains (Gozalo & Liñán, Reference Gozalo and Liñán1988; Sdzuy & Liñán, Reference Sdzuy and Liñán1993), the study of structural and metamorphic features associated with the major tectonic boundaries in the Iberian Massif, the Demanda Massif and the Iberian Chains showed that the Datos fault is more likely to represent this boundary (Álvaro et al. Reference Álvaro, Bauluz, Gil-Imaz and Ubide2018).
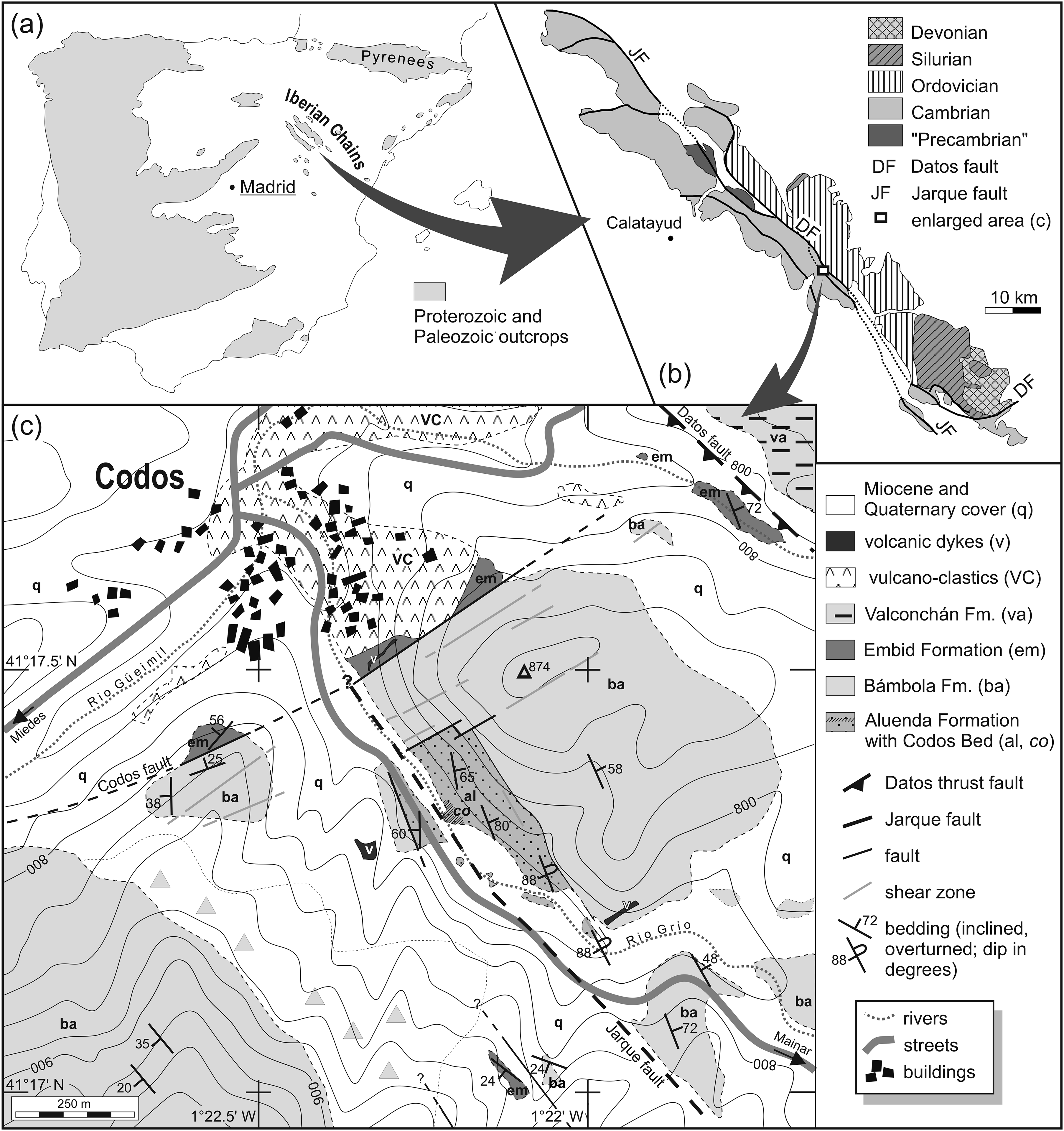
Fig. 1. Geological setting of the study area as part of the Eastern Iberian Chains. (a) Outcrops of Proterozoic and Palaeozoic rocks of the Iberian Peninsula (modified from Lotze, Reference Lotze1961, fig. 1). (b) Geology of the Eastern Iberian Chains with position of study area indicated (modified from Álvaro & Vennin, Reference Álvaro and Vennin1998, fig. 2). (c) Geological map of the study area.
The Paracuellos Group represents the oldest sedimentary sequence of the Iberian Chains. It has been formally subdivided into four siliciclastic formations which are, in stratigraphic order, the Sestrica, Saviñán, Frasno and Aluenda formations (Liñán & Tejero, Reference Liñán and Tejero1988). Dominant lithologies in the Sestricia, Saviñán and Aluenda formations are shales and fine- to medium-grained greywackes, whereas the Frasno Formation is characterized by an alternation of laminated cherts and shales. The stratigraphic framework of Liñán & Tejero (Reference Liñán and Tejero1988) was revised by Álvaro & Blanc-Valleron (Reference Álvaro and Blanc-Valleron2002) who recognized the same units but changed the name Frasno Formation to Frasno Bed. In addition, they studied the stratigraphy of the Codos area in the central Eastern Iberian Chains where they recognized two stratigraphic units of the Paracuellos Group, a new limestone bed, introduced as Codos Bed, and equivalents of the Aluenda Formation overlying this bed. Gámez Vintaned et al. (Reference Gámez Vintaned, Liñán, Navarro and Zhuravlev2018), in contrast, considered the Codos succession to be lithologically distinct from the established units of the Paracuellos Group, and consequently described the Codos Bed together with the overlying siliciclastics informally as Codos formation. The stratigraphy of Álvaro & Blanc-Valleron (Reference Álvaro and Blanc-Valleron2002) is followed herein.
The age of the Paracuellos Group has traditionally been considered as Neoproterozoic based on its stratigraphic position (e.g. Lotze, Reference Lotze1961), the occurrence of a trace fossil identified as Torrowangea aff. rosei Webby in the middle part of the Saviñán Formation (Liñán & Tejero, Reference Liñán and Tejero1988), and fossils identified as or compared with Cloudina Germs (Liñán et al. Reference Liñán, Grant and Navarro Vázquez1994; M. Streng, unpub. Diploma thesis, Univ. Würzburg, 1996; Álvaro & Blanc-Valleron, Reference Álvaro and Blanc-Valleron2002). The group is exposed in two geographically separated areas where it forms the core of faulted anticlines, i.e. the Paracuellos antiform and the Codos antiform in the northwestern and central Eastern Iberian Chains, respectively (Álvaro & Blanc-Valleron, Reference Álvaro and Blanc-Valleron2002). In the Paracuellos antiform, the boundary between the Paracuellos Group, i.e. the Aluenda Formation, and the overlying Bámbola Formation is an erosive contact and indicates a hiatus (Liñán & Tejero, Reference Liñán and Tejero1988; Álvaro & Blanc-Valleron, Reference Álvaro and Blanc-Valleron2002). In contrast, a more gradual transition between the shales of the Aluenda Formation and the sandstones and conglomerates of the Bámbola Formation has been described in the Codos antiform (M. Streng, unpub. Diploma thesis, Univ. Würzburg, 1996; Álvaro & Blanc-Valleron, Reference Álvaro and Blanc-Valleron2002; but see Discussion, Section 3). The Ediacaran–Cambrian boundary was traditionally placed at the base of the Bámbola Formation.
The putative Cloudina fossils mentioned by Liñán et al. (Reference Liñán, Grant and Navarro Vázquez1994) and the specimens described and illustrated as Cloudina? sp. in an unpublished thesis (M. Streng, unpub. Diploma thesis, Univ. Würzburg, 1996) are from the Codos Bed underlying the Aluenda Formation in the Codos antiform (Álvaro & Blanc-Valleron, Reference Álvaro and Blanc-Valleron2002). However, in the first description of the specimens it was already pointed out that the tube-shaped fossils lack the cone-in-cone structure characteristic of Cloudina (M. Streng, unpub. Diploma thesis, Univ. Würzburg, 1996). In addition, cross-sections of a variety of other tubular small shelly fossils had been observed within the carbonate bed. Some of these cross-sections showed internal septate structures and/or external ornamentation (M. Streng, unpub. Diploma thesis, Univ. Würzburg, 1996) indicating a diversity of shelly fossils unusual for Neoproterozoic strata. Similar observations were also made by Álvaro et al. (Reference Álvaro, Shields-Zhou, Ahlberg, Jensen and Palacios2016) who placed the Aluenda Formation and the Codos Bed questionably into the early Cambrian (‘Tommotian’, i.e. ≈ Cambrian Stage 2). When Gámez Vintaned et al. (Reference Gámez Vintaned, Liñán, Navarro and Zhuravlev2018) restudied the section (their ‘Codos formation’), they also reported a variety of shelly fossils from the carbonate bed. They mention the presence of at least seven genera, including taxa such as Cupitheca Duan, Cobboldiella Kerber and Pseudorthotheca Cobbold as well as halkieriid sclerites. However, none of the taxa was further described or illustrated except for a new genus and species with a tube-shaped shell introduced as Codositubulus grioensis. Despite the rich fossil content and the identification of common early Cambrian taxa, Gámez Vintaned et al. (Reference Gámez Vintaned, Liñán, Navarro and Zhuravlev2018) argue for a latest Ediacaran or earliest Cambrian age for this association. One of their reasons for this age assignment is the supposed absence of molluscan shelly fossils in the Codos Bed, an argument they also used to dismiss the correlation of the Codos Bed with the Heraultia Limestone Member of the Marcou Formation of the Montagne Noire (southern France) as previously suggested by Álvaro et al. (Reference Álvaro, Monceret, Monceret, Verraes and Vizcaïno2010, Reference Álvaro, Shields-Zhou, Ahlberg, Jensen and Palacios2016, Reference Álvaro, Cortijo, Jensen, Lorenzo, Palacios and Pieren2019).
Reinvestigation of crack-out material of the Codos Bed presented herein has revealed the presence of molluscan taxa (internal moulds of the helcionelloids Anabarella cf. plana Vostokova and Igorella? sp.) alongside common tubular fossils. In addition, the geology of the Codos anticline region is supplemented and amended in this article.
2. Material and methods
All specimens described herein are from the Codos Bed exposed along Río Grío at 41° 17′' 20″ N, 1° 22′ 14″ W, c. 600 m SE of Iglesia Santa María Magdalena, the church of the nearby village Codos. At the sample locality, c. 1 m of the Codos Bed is exposed, representing a phosphate-rich, dolomitic wackestone, which contains packstone laminae and lenses rich in ooids and skeletal debris (see also Álvaro & Blanc-Valleron, Reference Álvaro and Blanc-Valleron2002; Gámez Vintaned et al. Reference Gámez Vintaned, Liñán, Navarro and Zhuravlev2018; Fig. 2e–i). Tubular fossils were obtained by the dissolution of packstone-rich samples in diluted formic acid (c. 10 %) and subsequent mechanical preparation with a steel needle. The two helcionellid specimens were recovered by carefully splitting collected bulk material of the bed. Specimens were studied using incident light photography, scanning electron microscopy (SEM) and energy-dispersive spectroscopy (EDS). For incident light photography the specimens were coated with a sublimate of ammonium chloride to enhance contrast and then photographed with a Nikon DS-5M digital camera attached to a stereomicroscope. For SEM photography the ammonium chloride coating was removed and specimens were either first sputter-coated with a gold–palladium alloy (helcionellids) or left uncoated (tubes). SEM photography and EDS analyses of specimens and rock matrix were performed using a field emission scanning electron microscope (Zeiss Supra 35VP) equipped with an Apollo X Ametek energy-dispersive spectrometer.
Fig. 2. Different components of the Codos Bed revealed through treatment of the rock with formic acid. (a) Longitudinal fracture of a tubular fossil (SNSB-BSPG PIW1996IV6) showing traces of potential internal compartmentalization; remnant of original shell appears white. (b) EDS analysis of specimen shown in (a) indicating that shell remnants and sediment filling of the tube have high concentrations of phosphate and calcium. (c) Elliptical cross-section of a tubular fossil; mould is enriched in phosphate and calcium (d). Note generally denser texture of sediment within the tubes (moulds) shown in (a) and (c) versus the surrounding matrix. (e, f) Details of an oolitic lamina of the Codos Bed showing cross-sections of unevenly phosphatized ooids; phosphate-rich areas are lighter in colour as confirmed by EDS analysis. Specimens shown in (a), (c), (e) and (g–i) are uncoated and photographed under polarized incident light. Picture width equals 1 mm in (a), (c), (e), (g), (h), and 2 mm in (i). Abbreviations: P = phosphate, Ca = calcium, Mg = magnesium, Si = silicon.
Specimens illustrated and described herein are deposited in the Bavarian State Collection for Palaeontology and Geology (Bayerische Staatssammlung für Paläontologie und Geologie) in Munich, Germany (acronym SNSB-BSPG PIW) and the Museum of Natural Sciences of the University of Zaragoza (Museo de Ciencias Naturales, Universidad de Zaragoza) in Spain (acronym MPZ).
3. Geology of the grander Codos anticline
The study area southeast of the village of Codos (Fig. 1) is part of a geotectonically complex region of the Eastern Iberian Chains. The area is characterized by NW–SE-striking faults and to a lesser degree by faults roughly perpendicular to this direction. In addition, various types of magmatic activity are associated with the faults, and several intrusive rocks as well as widely distributed coarse volcanoclastic sediments (greywacke of Lago et al. Reference Lago, Andrés, Arranz, Galé and Pocoví2005; agglomerate of Gámez Vintaned et al. Reference Gámez Vintaned, Liñán, Navarro and Zhuravlev2018) can be observed in the area (M. Streng, unpub. Diploma thesis, Univ. Würzburg, 1996; Fig. 1c). All three tectonostratigraphic units of the Iberian Chains are exposed in proximity there.
A major structural element of the area is the NW–SE-trending Datos fault, which separates the upper Cambrian (Furongian) Valconchán Formation of the Herrera Unit in the NE from the lower Cambrian Embid Formation of the Mesones Unit in the SW (Fig. 1c), entailing a displacement of more than 2300 m (based on stratigraphic data in Álvaro, Reference Álvaro1995). Parallel to and c. 1 km southwest of the Datos fault runs another fault, which appears to be equivalent to the axial plane of a SE-plunging anticline, described as Codos antiform by Álvaro & Blanc-Valleron (Reference Álvaro and Blanc-Valleron2002). The fault divides the Bámbola and Aluenda formations into a NE- and a SW-dipping limb before its course is covered beneath younger Neogene and volcanoclastic sediments towards the northwest (Fig. 1c). The fault has been interpreted to be equivalent to the Jarque fault (e.g. Álvaro et al. Reference Álvaro, Bauluz, Gil-Imaz and Ubide2018; Gámez Vintaned et al. Reference Gámez Vintaned, Liñán, Navarro and Zhuravlev2018), and accordingly the NE limb of the anticline would belong to the Mesones, and its SW limb to the Badules unit. The Codos Bed is only known from the NE limb (= Mesones unit), where it represents the stratigraphically lowest exposed unit. As the fault plane of neither the Datos nor the Jarque fault is exposed in the area, their inclination and vergence are unknown. However, a NE vergence of the Jarque fault is indicated by differences in the degree of inclination of the strata of the Aluenda Formation between the two limbs of the anticline. Beds of the Aluenda Formation of the NE limb are almost vertical to slightly overturned, contrasting with the more moderately inclined layers of the SW limb (Fig. 1c).
Another significant structural element of the study area is a fault running NE–SW, roughly perpendicular to the Jarque and Datos faults. This fault, termed Codos fault herein, can be followed from outcrops southeast and south of Codos to exposures c. 1.5 km southwest of the village along the road (A-1504) to Miedes (M. Streng, unpub. Diploma thesis, Univ. Würzburg, 1996). It displaces the Embid Formation in the NW against the Bámbola Formation in the SE and is locally associated with up to 2 m broad dikes of quartz andesite cross-cutting the Embid Formation (Fig. 1c). Accompanying the Codos fault are several shear planes within the Bámbola Formation, indicated by cataclastites and coarsely brecciated zones parallel to the fault. The SW–NE-striking faulted contact between the Bámbola and Aluenda formations in the NE limb of the anticline (Fig. 1c) is also parallel to the fault and therefore interpreted to be related to it. The NE–SW trend of the Codos fault agrees with late Variscan extensional events (e.g. Lago et al. Reference Lago, Álvaro, Arranz, Pocoví and Vaquer1992, Reference Lago, Andrés, Arranz, Galé and Pocoví2005), and the fault is thus interpreted as a normal fault which is supported by the associated volcanic intrusion. The opposition of beds from the lower and middle Bámbola Formation against the Embid Formation suggests a displacement of more than 100 m for the Codos fault.
Discussion. As outlined above, the Datos and Jarque faults are the two main structural elements of the Eastern Iberian Chains, separating the three recognized tectonostratigraphic units. The displacements observed along these faults are substantial (e.g. Sanz et al. Reference Sanz, Lago, Gil, Galé, Ramajo, Ubide, Pocoví, Tierz and Larrea2013, fig. 11; Álvaro et al. Reference Álvaro, Bauluz, Gil-Imaz and Ubide2018, fig. 2), and this is also true for the Datos fault in the study area (2300 m). However, the displacement along the fault southeast of Codos (equated with the Jarque fault to date) is insignificant. This fault matches the axial plane of a NE-verging anticline, and accordingly the formations on either side of the fault match up (Fig. 1c), implying no or only a minor displacement. The reconstructed NE vergence of the fault matches the Jarque fault (e.g. Álvaro et al. Reference Álvaro, Bauluz, Gil-Imaz and Ubide2018), but its character as a thrust or reverse fault with large displacement cannot be confirmed. Hence, it appears questionable that the fault dividing the limbs of the Codos antiform is an equivalent of the Jarque fault.
The SW–NE-striking tectonic contact between the Aluenda and Bámbola formations has also been noted by Álvaro et al. (Reference Álvaro, Bauluz, Gil Imaz and Simón2008). However, they interpreted the contact (= their ‘Codos fault’) as the result of early Cambrian synsedimentary extensional tectonics, and the associated breccias as sedimentary channel infills. Gámez Vintaned et al. (Reference Gámez Vintaned, Liñán, Navarro and Zhuravlev2018, fig. 2a) measured the section of their Codos formation (= Aluenda Formation herein) and the transition to the overlying Bámbola Formation close to this tectonic contact. They described the boundary between the two units as unconformable and erosional, indicated by basal breccias in the Bámbola Formation. This contrasts with earlier observations of a gradual transition between the two formations at this locality where the transition was logged in a shallow valley c. 300 m southeast of the tectonic contact (M. Streng, unpub. Diploma thesis, Univ. Würzburg, 1996, fig. 6; see also Álvaro & Blanc-Valleron, Reference Álvaro and Blanc-Valleron2002). The erosional contact with basal breccias described by Gámez Vintaned et al. (Reference Gámez Vintaned, Liñán, Navarro and Zhuravlev2018) is here interpreted to be most likely related to the proximity of the measured section to the tectonic contact rather than being representative for the Codos succession. The possibility of a lateral facies change, however, cannot readily be excluded. In the Paracuellos antiform, the lower Bámbola Formation is known for its variable facies and thickness, and beside the typical unconformity between the Aluenda and Bámbola formations, gradual transitions have been observed there as well (Álvaro et al. Reference Álvaro, Bauluz, Gil Imaz and Simón2008).
Gámez Vintaned et al. (Reference Gámez Vintaned, Liñán, Navarro and Zhuravlev2018) reported a NNE–SSW-trending fault that supposedly divides the Aluenda Formation into two lithosomes, a ‘lower’ and an ‘upper’ one. The upper lithosome is equivalent to the described section of their Codos formation, which has a fossiliferous phosphatic dolostone at its base and is exposed close to the SW–NE-striking tectonic contact with the Bámbola Formation (Gámez Vintaned et al. Reference Gámez Vintaned, Liñán, Navarro and Zhuravlev2018, fig. 2a). Although not explicitly mentioned by Gámez Vintaned et al. (Reference Gámez Vintaned, Liñán, Navarro and Zhuravlev2018), it must be assumed that the phosphatic dolostone is an equivalent to the Codos Bed as previously described and illustrated from the same succession (Álvaro & Blanc-Valleron, Reference Álvaro and Blanc-Valleron2002; Álvaro et al. Reference Álvaro, Bauluz, Gil Imaz and Simón2008, fig. 9A). The lower lithosome is exposed less than 100 m to the southeast of the measured section and purportedly lacks this phosphatic dolostone. However, the locality of the Codos Bed reported herein is c. 150 m SE of the tectonic contact with the Bámbola Formation, in an area technically representing the lower lithosome of Gámez Vintaned et al. (Reference Gámez Vintaned, Liñán, Navarro and Zhuravlev2018). The existence of a fault dividing the two supposed lithosomes cannot be confirmed herein. Nevertheless, the results of Gámez Vintaned et al. (Reference Gámez Vintaned, Liñán, Navarro and Zhuravlev2018) imply that the Codos Bed might occur at not less than two distinct nearby localities. This also explains the differences in the reported thicknesses of the Codos Bed, for which up to 4 m (Álvaro & Blanc-Valleron, Reference Álvaro and Blanc-Valleron2002; Álvaro et al. Reference Álvaro, Bauluz, Gil Imaz and Simón2008) and 7 m (Gámez Vintaned et al. Reference Gámez Vintaned, Liñán, Navarro and Zhuravlev2018) have been reported close to the tectonic contact, whereas only 1 m is exposed 150 m to the southeast (see Section 2; Fig. 1c).
4. Preservation in Codos Bed
Acid treatment of the samples collected from the Codos Bed led to a partial dissolution of the rock without the rock completely disintegrating. The partial dissolution revealed common tubular fossils that were more resistant to the acid treatment than the rock matrix. Fossils are typically preserved as internal moulds and differ from the undissolved remaining rock matrix by their somewhat darker colour and denser texture. Individual specimens can preserve remnants of the original shell which is recognizable by its white colour (Fig. 2a). EDS analysis of the specimens revealed that the internal moulds are consistently enriched in phosphorus and calcium when compared with the rock matrix (Fig. 2b, d). The magnesium content is lower in the mould than in the matrix, whereas silica as well as iron and aluminium are evenly distributed except for areas with shell preservation. All analysed shell remnants are preserved as calcium phosphate. Although the shell of most tubular fossils is either absent or poorly preserved, one specimen shows a more solidly preserved shell (Fig. 3a, f), which also consists of calcium phosphate. Other components are ooids, which are concentrated in discontinuous laminae. Ooids are spherical or compressed spherical in shape and vary in diameter from 0.15 to 0.5 mm. In cross-section, ooids reveal white and grey zones as well as remnants of their concentric ultrastructure and occasionally a nucleus (Fig. 2e–i). EDS analyses of the ooids indicate that they consist mainly of calcium phosphate similar to the preserved shell material.

Fig. 3. Tubular fossils of the Codos Bed mechanically freed from surrounding matrix following acid treatment of the rock. (a, f) Specimen SNSB-BSPG PIW1996IV2 in lateral view, and detail of narrow end of tube. (b, e, g) Internal mould of specimen SNSB-BSPG PIW1996IV1, a potential coleolid, showing oblique ‘ornamentation’ and elliptical cross-section. (c, d) Lateral views of two conical(?), spine-shaped internal moulds with elliptical cross-sections (specimens SNSB-BSPG PIW1996IV7 and SNSB-BSPG PIW1996IV8). Incident light-photography except (f) (= SEM photography). Scale bar equals 1 mm except for (f), 280 µm.
Helcionellid specimens from untreated crack-out material are, similar to most tubular fossils, preserved as internal moulds without any remnants of the original shell. The phosphate concentration of the mould is distinctly higher than that of the rock matrix, as in the tubular fossils.
Discussion. The incomplete dissolution of the samples during acid preparation and the magnesium content of the residual rock matrix agree with previous descriptions of the Codos Bed as a dolostone (Álvaro & Blanc-Valleron, Reference Álvaro and Blanc-Valleron2002; Álvaro et al. Reference Álvaro, Bauluz, Gil Imaz and Simón2008). High phosphate contents of the moulds indicate diagenetic phosphatization processes which apparently not only affected the moulds but also shells and ooids. The phosphate content of the ooids is not homogeneous but typically irregularly distributed (Fig. 2e). This indicates diagenetic processes during which the original calcium carbonate was replaced by calcium phosphate. In analogy with the phosphatization of the ooids, the phosphatic composition of the shell remnants of the tubular fossils is considered to be of secondary origin. However, a primary phosphatic composition is possible for the single tubular specimen (SNSB-BSPG PIW1996IV2) which differs in having a distinctly stronger shell than the remaining specimens. Here, diagenetic processes might have overprinted the original phosphatic composition through recrystallization which destroyed ultrastructural details, as well as a possible faint external ornamentation, but preserved the structural strength of the shell (Fig. 3a, f).
5. Systematic palaeontology
Phylum indet.
Tubular fossils spp. indet.
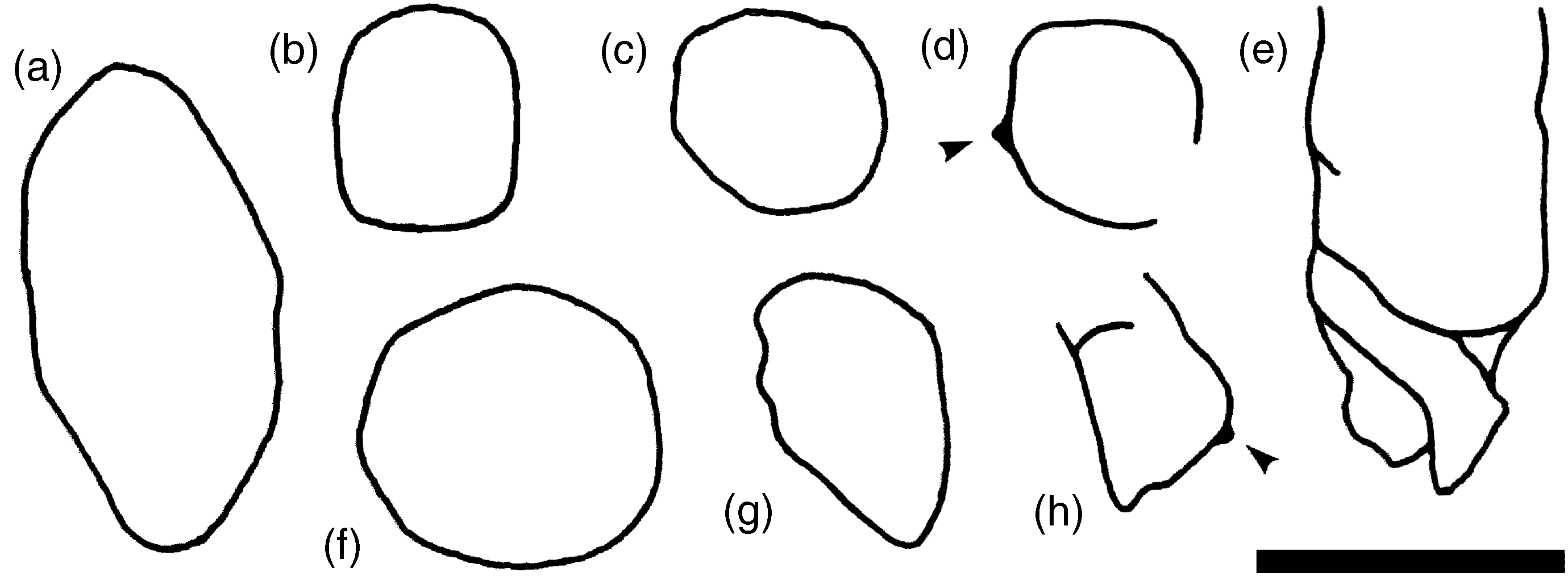
Fig. 4. Camera lucida drawings of various cross-sections of tubular fossils observed in the Codos Bed. Remnants of potential external ornamentation are preserved in (d) and (h) (arrows) whereas (e) and (h) show internal compartmentalization. Scale bar equals 1 mm.
Material. About 20 specimens from the Codos Bed, SE of Codos, eastern Iberian Chains, Spain; all specimens are incomplete and the best five are illustrated (SNSB-BSPG PIW 1996IV1, SNSB-BSPG PIW 1996IV2, SNSB-BSPG PIW 1996IV6, SNSB-BSPG PIW1996IV7, SNSB-BSPG PIW1996IV8).
Description. Specimens mainly represent internal moulds of tapering tubular structures (Figs 2a, 3a–e). They have elliptical, subrectangular to subcircular cross-sections (Figs 2c, 3g, 4a–c, f, g) with maximum measured diameters of c. 1.7 mm and lengths of c. 5 mm. Moulds straight or slightly curved, sometimes lined with remnants of original mineralized shell, which appears as white phosphatic substance under incident light (e.g. Fig. 2a). Cross-cut specimens occasionally reveal traces of a potential internal compartmentalization or septation (Figs 2a, 4e, h) as well as local shell thickenings reminiscent of external ornamentation (Fig. 4d, h). Surface of moulds smooth, but one specimen with evenly spaced, oblique transverse ribs which are inclined at c. 60° to the long axis of the specimen (Fig. 3b, e). All specimens appear to be incomplete, being broken at either end or incompletely phosphatized.
Discussion. Considering the observed variability of the tubular structures, it is evident that several species have been summarized in the description above. A clear distinction between species is mainly hampered by the generally incomplete preservation of the specimens. Even the actual shape of the specimens is impossible to determine precisely, i.e. whether the specimens represent tapering tubes or broken pointy cones. Furthermore, only a few specimens are exposed three-dimensionally, contrasting with the majority of tubes which are only seen in transverse sections (Fig. 2c). The majority of the specimens appears to have an elliptical cross-section, which is likely to portray the original shape rather than resulting from compression or distortion as neither the tubes nor the observed long axes of the elliptical cross-sections show a preferred orientation. However, compaction is also observed in the Codos Bed, indicated by oriented elliptical outlines of the ooids (Fig. 2i).
The few better-exposed specimens (Figs 2a, 3) merit a short discussion: Specimen SNSB-BSPG PIW1996IV2 (= Cloudina-like microfossil of Álvaro & Blanc-Valleron, Reference Álvaro and Blanc-Valleron2002, fig. 4C; Fig. 3a, f) represents the specimen with the best-preserved shell remnants, which appears to indicate that this specimen had a primary phosphatic shell (see Section 4 above). The 5 mm long specimen has a subcircular diameter of 0.8 mm at the narrow end and an elliptical cross-section with a maximum diameter of 1.4 mm at the broader end. Characters such as the tapering and slightly curved shell in addition to the elliptical cross-section and the presumed primary phosphatic shell mineralogy are suggestive of the genus Torellella Holm and related taxa (e.g. Parkhaev & Demidenko, Reference Parkhaev and Demidenko2010). However, uncertainties regarding the shell ultrastructure and original shell mineralogy of the Codos specimen do not allow a confident assignment. The surface of the tube shows minute tubercles (5–6 µm in diameter; Fig. 3f), which match those considered to be diagnostic for Codositubulus, a tube-shaped taxon recently introduced from the same bed (Gámez Vintaned et al. Reference Gámez Vintaned, Liñán, Navarro and Zhuravlev2018). However, in contrast to specimen SNSB-BSPG PIW1996IV2, Codositubulus shows distinct transverse annuli similar to the orthothecid Khetatheca Missarzhevsky. The analogous minute tubercular ornamentation is here interpreted as the result of recrystallization of the original shell substance, rather than representing primary ornamentation, and hence is of no systematic value.
The 4 mm long specimen SNSB-BSPG PIW1996IV1 (Fig. 3b, e, g) represents an internal mould which has a broadly elliptical cross-section, a distinct bend near the narrow end (Fig. 3g) and, in contrast to all other specimens, bears an enigmatic ornamentation of regularly spaced parallel ridges that are inclined towards the main axis of the specimen. Such features are reminiscent of the problematic family Coleolidae Fisher, Reference Fisher and Moore1962, a most likely polyphyletic group of organisms characterized by a gently tapering, elongate conical calcareous shell which may be ornamented by oblique ridges (Fisher, Reference Fisher and Moore1962; Malinky et al., Reference Malinky, Wilson, Holmer, Lardeux, Webby, Paris, Droser and Percival2004). Salopiella Cobbold, originally described from the early Cambrian Strenuella Limestone of Comley, England (Cobbold, Reference Cobbold1921), is the only coleolid taxon in which the ornamentation of the shell is also reproduced on the shell interior. Thus, internal moulds of Salopiella might appear similar to the Codos specimen. Of the two described species of Salopiella, specimen SNSB-BSPG PIW1996IV1 is most similar to the type species Salopiella obliqua Cobbold, Reference Cobbold1921, which, however, has a less inclined ornamentation as well as asymmetric and more densely spaced ridges. Nevertheless, an affiliation of the Codos specimen with the coleolids and especially Salopiella appears likely.
The orthothecid Turcutheca Missarzhevsky is characterized by having a smooth internal mould, a gently tapering conch and an oval cross-section (Missarzhevsky, Reference Missarzhevsky, Rozanov, Missarzhevsky, Volkova, Voronova, Krylov, Keller, Korolyuk, Lendzion, Mikhnyak, Pykhova and Sidorov1969), features also observed in two c. 5 mm long specimens from the Codos Bed (SNSB-BSPG PIW1996IV7, SNSB-BSPG PIW1996IV8; Fig. 3c, d). However, as with all the other specimens described herein, the incomplete preservation (missing apices) would make any systematic placement conjectural. Their resemblance to many of the tubular taxa characteristic of the early Cambrian (late Fortunian to early Cambrian Stage 2) is emphasized though.
Compartmentalization or septation in early Cambrian biomineralizing taxa is known from various orthothecid hyoliths (e.g. Missarzhevsky, Reference Missarzhevsky, Rozanov, Missarzhevsky, Volkova, Voronova, Krylov, Keller, Korolyuk, Lendzion, Mikhnyak, Pykhova and Sidorov1969; Landing, Reference Landing1988; Kouchinsky et al. Reference Kouchinsky, Bengtson, Landing, Steiner, Vendrasco and Ziegler2017), aldanellid mollucs (Parkhaev, Reference Parkhaev, Ponder and Lindberg2008), and cap-shaped mollusc-like taxa such as Tannuella Missarzhevsky (e.g. Missarzhevsky, Reference Missarzhevsky, Rozanov, Missarzhevsky, Volkova, Voronova, Krylov, Keller, Korolyuk, Lendzion, Mikhnyak, Pykhova and Sidorov1969; Brock & Paterson, Reference Brock and Paterson2004). However, unlike in these taxa in which the septa are arranged roughly perpendicular to the outer shell layer and convex towards the apex, the septa in the specimens from the Codos Bed appear to be distinctly inclined to the outer shell wall and non-curved (Figs 2a, 4e), leaving their systematic significance puzzling.
Phylum Mollusca Cuvier, Reference Cuvier1797
Helcionelloids sensu Peel, Reference Peel, Simonetta and Morris1991
Remarks. The plethora of small, cap-shaped to partly coiled, presumably endogastric mollusc-like shells of the Cambrian and Ordovician is typically accommodated within the class Helcionelloidea Peel, Reference Peel, Simonetta and Morris1991 (e.g. Peel, Reference Peel, Simonetta and Morris1991; Geyer, Reference Geyer1994; Gubanov & Peel, Reference Gubanov and Peel2000, Reference Gubanov and Peel2001; Devaere et al. Reference Devaere, Clausen, Steiner, Álvaro and Vachard2013; Geyer et al. Reference Geyer, Valent and Meier2019; Li et al. Reference Li, Zhang, Skovsted, Yun, Pan and Li2021). However, detailed relationships of groups within the class are highly debated and controversial, as is their systematic relationship with molluscan crown groups, particularly with the Gastropoda (e.g. discussions in Geyer et al. Reference Geyer, Valent and Meier2019; Li et al. Reference Li, Zhang, Skovsted, Yun, Pan and Li2021 vs Parkhaev, Reference Parkhaev2017 a, b). Consensus about helcionelloid relation- and interrelationships is mainly hampered by lack of knowledge regarding anatomical details of the proposed helcionelloid families, specifically whether they would have undergone developmental torsion or not. Recently discovered preserved bundles of chitinous chaetae in an early Cambrian species of Pelagiella, a taxon also commonly placed with the helcionelloids, allowed for the first time the reconstruction of the soft tissue of a supposed helcionelloid (Thomas et al. Reference Thomas, Runnegar and Matt2020). As a result, Pelagiella has been interpreted to shows signs of torsion and placed in the stem lineage of the Gastropoda (Thomas et al. Reference Thomas, Runnegar and Matt2020). However, Landing et al. (Reference Landing, Geyer, Jirkov and Schiaparelliin press) oppose this conclusion and interpret the chaetae as paleal fan-arrays characteristic for sedentary polychaetes. They argue convincingly that the majority of the so-called pelagiellids are distinct from the type species of the genus and should be referred to the sabellid polychaetes under the new name Pseudopelagiella. The opposing results of Thomas et al. (Reference Thomas, Runnegar and Matt2020) and Landing et al. (Reference Landing, Geyer, Jirkov and Schiaparelliin press), however, do not imply that all helcionelloids are either stem-gastropods or polychaetes, but rather that the ‘pelagiellids’, i.e. Pseudopelagiella and potential related taxa, should be regarded as distinct from the group. Helcionelloids, with the possible exception of the ‘pelagiellids’ (see Landing et al. Reference Landing, Geyer, Jirkov and Schiaparelliin press), most likely represent a paraphyletic grade that is part of the stem lineage of several molluscan lineages (e.g. Peel, Reference Peel, Simonetta and Morris1991, Reference Peel2004, Reference Peel2006; Kouchinsky, Reference Kouchinsky1999; Thomas et al. Reference Thomas, Runnegar and Matt2020). Likewise, it cannot be excluded that many helcionelloid families are also paraphyletic. Accordingly, no family assignment of the taxa described in the following is attempted.
Genus Anabarella Vostokova, Reference Vostokova1962
Type species. Anabarella plana Vostokova, Reference Vostokova1962, from ‘Aldanian’ Regional Stage (late Fortunian – early part of Cambrian Stage 2 according to Kouchinsky et al. Reference Kouchinsky, Bengtson, Landing, Steiner, Vendrasco and Ziegler2017); Kenyada river area, Olenyok District, northern Siberian Platform.
Diagnosis. See Gubanov & Peel (Reference Gubanov and Peel2003).
Anabarella cf. plana Vostokova, Reference Vostokova1962
Fig. 5a, b, d, e

Fig. 5. Helcionellid specimens of the Codos Bed. (a, b, d, e) Internal mould of Anabarella cf. plana Vostokova, Reference Vostokova1962 (MPZ 2021/52). (a) Lateral view; (d, e) oblique lateral views showing indistinct ornamentation; compare with schematic reconstruction of specimen (b). (c, f) Lateral and oblique view of internal mould of Igorella? sp. (MPZ 2021/53). SEM photography; scale bar equals 1 mm.
Material. Single specimen (MPZ 2021/52) from the Codos Bed, SE of Codos, eastern Iberian Chains, Spain.
Description. Specimen preserved as a phosphatized internal mould that is partly exposed from the rock matrix; appears laterally compressed, 2.1 mm in length and 1.3 mm in height; highest point posterior to mid-length (at c. 56 % of length); loosely coiled, representing about half whorl with anterior field evenly convex and posterior field short and concave; diameter of mould rapidly expanding from a blunt but pointed apex towards a long aperture (c. 1.8 mm); apex projects c. 250 µm past the posterior margin of aperture. Surface of mould with five or six faint transverse ridges (Fig. 4d); most of apical part appears smooth (Fig. 5b).
Discussion. The specimen is quite similar to moulds of Anabarella plana described and illustrated by Gubanov & Peel (Reference Gubanov and Peel2003, pl. 2, figs 1–10) from the Pestrotsvet Formation of the southeastern part of the Siberian Platform but with a less coiled apical part. Anabarella plana has been described from many lower Cambrian strata, mainly from Russia (Siberian Platform), but also from Estonia (Baltica), southeastern Newfoundland (Avalonia) and Spain (western Gondwana) among others (see Kouchinsky et al. Reference Kouchinsky, Bengtson, Landing, Steiner, Vendrasco and Ziegler2017 for additional data). According to Kouchinsky et al. (Reference Kouchinsky, Bengtson, Landing, Steiner, Vendrasco and Ziegler2017), the species is restricted to late Fortunian and the early part of Cambrian Stage 2.
Igorella Missarzhevsky, Reference Missarzhevsky, Rozanov, Missarzhevsky, Volkova, Voronova, Krylov, Keller, Korolyuk, Lendzion, Mikhnyak, Pykhova and Sidorov1969
Type species. Igorella ungulata Missarzhevsky, Reference Missarzhevsky, Rozanov, Missarzhevsky, Volkova, Voronova, Krylov, Keller, Korolyuk, Lendzion, Mikhnyak, Pykhova and Sidorov1969, Tommotian Regional Stage, early Cambrian, West Anabar and Uchur-Maya regions, Siberian Platform.
Diagnosis. Devaere et al. (Reference Devaere, Clausen, Steiner, Álvaro and Vachard2013) provided an emended diagnosis of the genus, but Geyer et al. (Reference Geyer, Valent and Meier2019) consider the morphological concept of Igorella as ambiguous.
Igorella? sp.
Fig. 5c, f
Material. Single specimen (MPZ 2021/53) from the Codos Bed, SE of Codos, eastern Iberian Chains, Spain.
Description. Partly exposed, cap-shaped incomplete internal mould, c. 1.1 mm long and 0.9 mm high; with coarse transverse ridges that become more distinct towards the aperture. Anterior field convex, posterior field concave, poorly visible. Details of apex and aperture not preserved.
Discussion. The poor preservation of the specimen does not allow a confident systematic placement. Ornamentation and outline are reminiscent of species of Igorella, especially of I. emeiensis (Yu, Reference Yu1987) from the Meishucunian Regional Stage (Cambrian Stage 2) of Yunnan, South China, as illustrated in Parkhaev (Reference Parkhaev, Ponder and Lindberg2008, fig. 3.12A).
6. Age of the Codos fauna
In their description of Codositubulus from the Codos Bed, Gámez Vintaned et al. (Reference Gámez Vintaned, Liñán, Navarro and Zhuravlev2018) provide no clear-cut statement regarding the age of their new species and of the associated fauna. In the title of their article, they refer to the fauna as ‘the oldest Cambrian skeletal fossils of Spain’, i.e. implying an earliest Cambrian age, whereas in the text they also consider a pre-Terreneuvian age. Arguments for their age estimates are (1) an association dominated by simple tube-shaped shelly fossils as it is characteristic for the Ediacaran–Cambrian boundary interval and (2) the absence of other skeletal fossils, such as helcionellids, that would indicate a younger age. With the description of the two helcionellid taxa herein, the age of the Codos fauna needs to be revised as it clearly falls within the early Cambrian, more precisely into the late Fortunian or early part of Cambrian Stage 2 as indicated by Anabarella cf. plana (see Kouchinsky et al. Reference Kouchinsky, Bengtson, Landing, Steiner, Vendrasco and Ziegler2017). However, does this new age estimate still make the Codos skeletal assemblage the oldest in Spain? Comparable pre-trilobitic early Cambrian skeletal fossils have been reported from various localities in central Spain where Neoproterozoic to early Cambrian sequences are exposed in several anticlines and dome structures (for overview see e.g. Álvaro et al. Reference Álvaro, Cortijo, Jensen, Lorenzo, Palacios and Pieren2019, fig. 1). However, not all reported occurrences are without problems.
From the Alcudia anticline, simple tubular fossils were first reported by Vidal et al. (Reference Vidal, Palacios, Moczydłowska and Alvarez1995) in strata of the uppermost ‘Lower Alcudian’ and lowermost ‘Upper Alcudian’. The fossil assemblage was tentatively compared with the Anabarites trisulcatus fauna of the Siberian Platform and the specimens were subsequently referred to as anabaritids (Palacios et al. Reference Palacios, Vidal, Moczydlowska, Liñán, Gámez Vintaned and Gozalo1996; Vidal et al. Reference Vidal, Palacios, Moczydłowska and Gubanov1999; Fernández-Remolar, Reference Fernández-Remolar2001). From beds higher up in the ‘Upper Alcudian’, Vidal et al. (Reference Vidal, Palacios, Moczydłowska and Alvarez1995) also reported common helcionellid specimens which they preliminarily assigned to Anabarella and Bemella (= Anabarella plana of Palacios et al. Reference Palacios, Vidal, Moczydlowska, Liñán, Gámez Vintaned and Gozalo1996) and later described in detail as Anabarella sp. cf. A. plana (Vidal et al. Reference Vidal, Palacios, Moczydłowska and Gubanov1999). For the transitional beds at the ‘Lower’ to ‘Upper Alcudian’ boundary interval, a basal Cambrian or late ‘Vendian’ age was assumed whereas the upper ‘Upper Alcudian’ beds were compared with the Tommotian Regional Stage in Siberia (Vidal et al. Reference Vidal, Palacios, Moczydłowska and Alvarez1995; Palacios et al. Reference Palacios, Vidal, Moczydlowska, Liñán, Gámez Vintaned and Gozalo1996). In contrast to the helcionellid specimens, the alleged anabaritids from the transitional beds were never described in detail or illustrated. Also, the exact stratigraphic levels and details of the lithology of the fossiliferous beds were not mentioned, aggravating a correlation with the subsequently revised stratigraphy of the Alcudia anticline (Pieren & García-Hidalgo, Reference Pieren and García-Hidalgo1999; Álvaro et al. Reference Álvaro, Cortijo, Jensen, Lorenzo, Palacios and Pieren2019). In contrast to Vidal et al. (Reference Vidal, Palacios, Moczydłowska and Gubanov1999) who regarded the Alcudia succession as entirely conformable, Álvaro et al. (Reference Álvaro, Cortijo, Jensen, Lorenzo, Palacios and Pieren2019) describe it as non-continuous, characterized by two major stratigraphic gaps. The Ediacaran Lower Alcudian-Domo Extremeño Supergroup is unconformably followed by a probably late Ediacaran to earliest Cambrian succession of a local facies development of the Ibor Group comprising the Tamújar, Hinojosas and Cabezarrubias formations, which are nonconformably overlain by Terreneuvian units described as San Lorenzo and Fuentepizarra formations. Comparing the stratigraphy of Vidal et al. (Reference Vidal, Palacios, Moczydłowska and Gubanov1999, fig. 1D) with the revised concept of Álvaro et al. (Reference Álvaro, Cortijo, Jensen, Lorenzo, Palacios and Pieren2019, fig. 4), the ‘Alcudian’ transitional beds are best compared with the late Ediacaran to earliest Cambrian sequence despite being entirely part of the ‘Upper Alcudian’ sensu Pieren & García-Hidalgo (Reference Pieren and García-Hidalgo1999). As a strong dolomitization of the purported anabaritids was reported (Vidal et al. Reference Vidal, Palacios, Moczydłowska and Alvarez1995), it is assumed here that they might originate from the Hinojosas Formation which is the only unit of this sequence comprising carbonate beds (Álvaro et al. Reference Álvaro, Cortijo, Jensen, Lorenzo, Palacios and Pieren2019; see also Simón, Reference Simón2017). Trace fossils found within the Hinojosas Formation presumably indicate an early Cambrian age of this unit (Simón, Reference Simón2017; Álvaro et al. Reference Álvaro, Cortijo, Jensen, Lorenzo, Palacios and Pieren2019 and references therein). However, the systematic nature of the anabaritids remains enigmatic as they might also represent poorly preserved specimens of Cloudina, Sinotubulites or tubular problematica similar to those described from the late Ediacaran strata of the typical expression of the Ibor Group in the nearby Navalpino anticline (Cortijo et al. Reference Cortijo, Martí Mus, Jensen and Palacios2015). Stratigraphic harmonization of the ‘Upper Alcudian’ Anabarella specimens with the revised stratigraphy is less problematic. They are from phosphatic carbonate beds or phosphorites of what is now the middle Fuentepizzara Formation (Pieren & García-Hidalgo, Reference Pieren and García-Hidalgo1999; Álvaro et al. Reference Álvaro, Cortijo, Jensen, Lorenzo, Palacios and Pieren2019). From coeval levels, Álvaro et al. (Reference Álvaro, Cortijo, Jensen, Lorenzo, Palacios and Pieren2019, fig. 13E–G) recently reported and illustrated also tubular microfossils as well as additional specimens of Anabarella.
The occurrence of anabaritids was also marked in the middle ‘Río Huso Group’ of the Valdelacasa anticline (Palacios et al. Reference Palacios, Vidal, Moczydlowska, Liñán, Gámez Vintaned and Gozalo1996, fig. 1; Vidal et al. Reference Vidal, Palacios, Moczydłowska and Gubanov1999, fig. 1; Fernández-Remolar, Reference Fernández-Remolar2001, fig. 2) where they supposedly co-occur with halkieriids and sponges (Palacios et al. Reference Palacios, Gámez Vintaned, Fernández-Remolar and Liñán1999; Gozalo et al. Reference Gozalo, Liñán, Palacios, Gámez Vintaned and Mayoral2003; Zhuravlev et al. Reference Zhuravlev, Liñán, Gámez Vintaned, Debrenne and Fedorov2012). However, similar to the anabaritids from the Alcudia anticline, the specimens from the Valdelacasa anticline were never illustrated or described in detail. The same is true for the co-occurring other small shelly fossils. Only Perconig et al. (Reference Perconig, Vazquez Guzman, Velando and Leyva1983) and Reitner et al. (Reference Reitner, Luo and Duda2012) illustrated and described in some detail sponge spicules from potentially equivalent levels in the adjacent Navalpino anticline. The only illustrated pre-trilobitic Cambrian skeletal fossils from the Valdelacasa anticline are macroscopic Scenella-like molluscs which are also from the middle ‘Río Huso Group’ but from a somewhat higher level than the aforementioned small shelly fossils (Martí Mus et al. Reference Martí Mus, Palacios and Jensen2008). The middle ‘Río Huso Group’ is equivalent to the Pusa Formation which is known for the common occurrences of phosphorites in its middle part (e.g. Álvaro et al. Reference Álvaro, Cortijo, Jensen, Lorenzo, Palacios and Pieren2019). Reiter et al. (Reference Reitner, Luo and Duda2012) described their sponge spicules from these phosphate-rich levels and it is likely that the other mentioned small shelly fossils are from there as well. Furthermore, a direct correlation between the phosphate-bearing beds of the middle Pusa Formation and those of the Fuentepizarra Formation containing tubular fossils and Anabarella (see above) is likely (Álvaro et al. Reference Álvaro, Cortijo, Jensen, Lorenzo, Palacios and Pieren2019).
The Ediacaran–Cambrian boundary in central Iberia can best be traced in successions of the Ibor Group (sensu Álvaro et al. Reference Álvaro, Cortijo, Jensen, Lorenzo, Palacios and Pieren2019) where it occurs in the group’s upper part. The boundary interval is particularly well studied in the type area of the group, the Ibor and Navalpino anticlines. Here, Cloudina-bearing limestones of the heterolithic Villarta Formation indicate a late Ediacaran age (Cortijo et al. Reference Cortijo, Martí Mus, Jensen and Palacios2015; Álvaro et al. Reference Álvaro, Cortijo, Jensen, Lorenzo, Palacios and Pieren2019) whereas the overlying siliciclastic Arrocampo Formation includes in its basal part Central Iberia’s earliest record of Treptichnus pedum as well as specimens of Sabellidites in levels above, indicating a basal Cambrian age (Álvaro et al. Reference Álvaro, Cortijo, Jensen, Lorenzo, Palacios and Pieren2019). Simón (Reference Simón2018) studied a succession of the Ibor Group including the Ediacaran–Cambrian transition in the Abenójar dome. The succession there is heterolithic and was informally subdivided into three main units: a lower siltstone, a middle carbonate and an upper sandstone unit (Simón, Reference Simón2018). The lithology of the units and the recovered fossils suggest a direct comparison of the succession with the three formations of the Ibor Group as defined by Álvaro et al. (Reference Álvaro, Cortijo, Jensen, Lorenzo, Palacios and Pieren2019). Accordingly, the lower siltstone unit would correspond to the Castañar Formation, the middle carbonate unit comprising specimens of Cloudina to the Villarta Formation, and the upper sandstone unit with occurrences of Treptichnus pedum to the Arrocampo Formation. The fossils from the middle carbonate and upper sandstone unit bracket the Ediacaran–Cambrian transition analogous to the Ibor and Navalpino anticlines. Controversially, Simón (Reference Simón2018) described potential protoconodonts from basal limestones of his middle carbonate unit co-occurring with Cloudina. Protoconodonts are widely accepted to be part of the oldest Cambrian skeletal assemblages (e.g. Kouchinsky et al. Reference Kouchinsky, Bengtson, Landing, Steiner, Vendrasco and Ziegler2017; Wood et al. Reference Wood, Liu, Bowyer, Wilby, Dunn, Kenchington, Hoyal Cuthill, Mitchell and Penny2019) and would indicate a Cambrian age of the middle carbonate unit. If the middle carbonate unit is equivalent to the Villarta Formation, the presence of protoconodonts would have serious implications for the position of the Ediacaran–Cambrian boundary in Central Iberia. The nature of the alleged protoconodonts needs further investigation, however, because their shape, outline of cross-sections and ultrastructural details do not precisely match those of known protoconodonts. The specimens are here considered as tube-like problematica.
Other known early Cambrian associations of small shelly fossils from Spain are younger than Cambrian Stage 2 as indicated by co-occurring trilobites. Examples of such associations are faunas from the upper Pusa Formation of the Valdelacasa anticline (Palacios et al. Reference Palacios, Vidal, Moczydlowska, Liñán, Gámez Vintaned and Gozalo1996; Jensen et al. Reference Jensen, Palacios and Martí Mus2010) or the lower Pedroche Formation of the Cordoba area (e.g. Vidal et al. Reference Vidal, Palacios, Gámez Vintaned, Díez Balda and Grant1994; Fernádez-Remolar, Reference Fernández-Remolar2001; Liñán et al. Reference Liñán, Dies, Gámez Vintaned, Gozalo, Mayoral and Muñiz2005).
In summary, the Codos fauna is one of Spain’s oldest Cambrian skeletal assemblages but probably not the oldest. The fossil content of the Codos Bed, consisting of tubular skeletal fossils, alleged halkieriids, and anabarellids, as well as its lithology permit a direct comparison with the fossiliferous phosphorites and phosphatic levels of the middle member of the Pusa Formation and the middle Fuentepizarra Formation (see also Álvaro et al. Reference Álvaro, Cortijo, Jensen, Lorenzo, Palacios and Pieren2019, fig. 16). Accordingly, a roughly coeval age is assumed for the three fossiliferous stratigraphic levels. However, the limited biostratigraphic value of the known fossil remains does not allow a precise correlation. The purported anabaritids from the Alcudia anticline (Vidal et al. Reference Vidal, Palacios, Moczydłowska and Alvarez1995) might represent the oldest Cambrian skeletal fossils of Spain if their stratigraphic level (Hinojosas Formation; see above) and systematic affiliation can be confirmed.
7. Conclusions
The area around Codos is geologically more complex than previously reported. Displacement along the fault dividing the Codos anticline is insignificant, thus challenging its common interpretation as the equivalent of the Jarque thrust fault, a major first-order structure in the Eastern Iberian Chains. The Codos anticline is limited to the northwest by a normal fault striking perpendicular to the anticline’s axial plane. This fault, introduced as the Codos fault, entails a displacement of more than 100 m and is deemed to considerably affect the course of the anticline.
Helcionelloid specimens and common tubular fossils recovered from the Codos Bed are characteristic for the Terreneuvian Series and indicate a late Fortunian or early Cambrian Age 2 age of the Codos Bed. The lithology and fossils of the Codos Bed suggest correlation with comparable fossiliferous beds of the middle member of the Pusa Formation and the middle Fuentepizarra Formation in central Spain. The helcionellid specimens in combination with the taxa reported by Gámez Vintaned et al. (Reference Gámez Vintaned, Liñán, Navarro and Zhuravlev2018), i.e. Cobboldiella and Pseudorthotheca, also provide concrete palaeontological support for the previously proposed correlation of the Codos Bed with the Heraultia Limestone Member of the Marcou Formation of the Montagne Noire, southern France, a correlation that was previously mainly based on lithological similarities (Álvaro et al. Reference Álvaro, Monceret, Monceret, Verraes and Vizcaïno2010, Reference Álvaro, Shields-Zhou, Ahlberg, Jensen and Palacios2016, Reference Álvaro, Cortijo, Jensen, Lorenzo, Palacios and Pieren2019). The notion that the association from the Codos Bed is the oldest skeletal assemblage of Spain cannot be upheld.
The fossils from the Codos Bed unambiguously confirm that the Paracuellos Group is not in its entirety Neoproterozoic in age, but partly early Cambrian. The Ediacaran–Cambrian boundary lies stratigraphically below the Codos Bed within the 300 to 400 m thick sequence of the underlying formations. Palaeontological reinvestigation of the sandstones and shales of particularly the Saviñán Formation, as well as a re-evaluation of the stratigraphic significance of Torrowangea aff. rosei, are required to further constrain the position of the Ediacaran–Cambrian boundary.
Acknowledgements
This study is part of the results of a two-month field survey of the geology and palaeontology of the area around Codos in 1995. The study was initiated by G Geyer (Würzburg) and was made possible through a permit arranged by E Liñán (Zaragoza). I am grateful to J Keller and G Geyer (both Würzburg) for their commitment and assistance in the field. JJ Álvaro (Madrid) and S Jensen (Badajoz) are thanked for discussion and for providing literature. The constructive reviews of G Geyer and another, anonymous referee, in addition to the efforts of the associate editor S Jensen, improved the quality of the paper and are gratefully acknowledged.
Declaration of interest
None. This research received no specific grant from any funding agency, commercial or not-for-profit sectors.






 Open access
Open access