



1. Introduction
The transition of plants from water to land took hundreds of millions of years, with numerous progressive innovations and adaptations, reshaping Earth’s terrestrial landscapes in profound ways (Kenrick et al., Reference Kenrick, Wellman, Schneider and Edgecombe2012; De Vries & Archibald, Reference De Vries and Archibald2018). Macrofossils, spores and phylogenetic models provide various answers as to when vascular plants colonized land, contributing to a multitude of hypotheses (Kenrick et al., Reference Kenrick, Wellman, Schneider and Edgecombe2012; Edwards et al., Reference Edwards, Morris, Richardson and Kenrick2014; Edwards & Kenrick, Reference Edwards and Kenrick2015; Salamon et al., Reference Salamon, Gerrienne, Steemans, Gorzelak, Filipiak, Le Hérissé, Florentin, Cascales- Miñana, Brachaniec, Misz-Kennan, Niedźwiedzki and Wieslaw2018). While molecular phylogenies suggest that vascular plants originated in the Middle Cambrian period (∼500 Ma) (Morris et al., Reference Morris, Puttick, Clark, Edwards, Kenrick, Pressel, Wellman, Yang, Schneider and Donoghue2018; Servais et al., Reference Servais, Cascales-Miñana, Cleal, Gerrienne, Harper and Neumann2019; Donoghue et al., Reference Donoghue, Harrison, Paps and Schneider2021), the presence of spores attributed to non-vascular land plants, known as cryptospores, is documented only from Middle Ordovician (Darriwilian stage; 468-461 Ma) sediments onwards (Steemans et al., Reference Steemans, Hérissé, Melvin, Miller, Paris, Verniers and Wellman2009; Morris et al., Reference Morris, Wright and Edwards2012; Edwards & Kenrick, Reference Edwards and Kenrick2015). Additionally, tetrad spores, widely considered to originate from vascular plants, emerged during the Late Ordovician (Katian stage; ∼450 Ma) (Fig. 1a; Table 1) (Strother et al., Reference Strother, Al-Hajri and Traverse1996; Wellman & Gray, Reference Wellman and Gray2000; Steemans et al., Reference Steemans, Hérissé, Melvin, Miller, Paris, Verniers and Wellman2009; Edwards and Kenrick, Reference Edwards and Kenrick2015). Macrofossils of vascular plants, exemplified by Cooksonia, appear only in the mid- to late Silurian (Wenlockian stage ∼ 430 Ma onwards), though fragmentary remains seem to appear earlier in the mid- to late Ordovician (Fig. 1a; 1b; Table 1) (Wellman et al., Reference Wellman, Osterloff and Mohiuddin2003; Xu et al., Reference Xu, Wang, Huang, Tang, Wang, Liu and Yan2022).

Figure 1. (a) Ordovician-Silurian geologic time scale with interpreted ages associated with the Cooksonia micro- and macrofossil record (Strother et al. Reference Strother, Al-Hajri and Traverse1996; Salomon et al. 2018; Edwards & Feehan, Reference Edwards and Feehan1980; Edwards et al. Reference Edwards, Feehan and Smith1983; Edwards et al. Reference Edwards, Bassett and Rogerson1979; Edwards & Davies, Reference Edwards and Davies1976; Edwards & Rogerson, Reference Edwards and Rogerson1979. (b) Cooksonia plant reconstruction by Tim Bertelink.
Table 1. Micro- and macrofossil compilation of vascular plants from previous studies estimating the fossil age from biostratigraphy

References 1. Strother et al. (Reference Strother, Al-Hajri and Traverse1996) ; 2. Salomon et al. (2018); 3. Edwards & Feehan, (Reference Edwards and Feehan1980); 4. Edwards et al. (Reference Edwards, Feehan and Smith1983); 5. Edwards et al. (Reference Edwards, Bassett and Rogerson1979); 6. Edwards & Davies, (Reference Edwards and Davies1976); 7. Edwards & Rogerson, (Reference Edwards and Rogerson1979).
One challenge arises from the lack of close modern analogues for cryptospores, except for a few spore configurations found in select liverworts (Renzaglia et al., Reference Renzaglia, Crandall-Stotler, Pressel, Duckett, Schuette and Strother2015a, b). Cryptospores exhibit a sudden decline in taxonomic diversity and abundance during the Early Devonian period (latest Lochkovian, ∼410 Ma), with only a few forms persisting through to the late Early Devonian (Emsian stage; 408-393 Ma) (Wellman et al., Reference Wellman, Steemans and Vecoli2013). Another complication concerns the preservation of diagnostic features of vascular plants, such as conducting tissue. The earliest preserved fragments displaying stomata-bearing epidermis, trilete spores and well-developed tracheids with distinct lateral wall facets, but without sporangia, date back to the lower Sandbian period (Upper Ordovician, ∼ 458 Ma) of Inner Mongolia (Xu et al., Reference Xu, Wang, Huang, Tang, Wang, Liu and Yan2022). However, there is then a gap until these features are recorded, with sporangia first appearing in the Wenlockian period (∼430 Ma) in Europe (Edwards & Davis, Reference Edwards and Davies1976).
In the Early Paleozoic, the development of primitive root hair structures by Cooksonia wielded a transformative influence on Earth. This transformation was manifested by amplified weathering rates, soil development, landscape stabilization and consequential alterations to Earth’s biogeochemical cycles, encompassing atmospheric oxygenation, and the carbon and nitrogen cycles (Wellman, Reference Wellman2010; Lenton et al., Reference Lenton, Crouch, Johnson, Pires and Dolan2012; Salamon et al., Reference Salamon, Gerrienne, Steemans, Gorzelak, Filipiak, Le Hérissé, Florentin, Cascales- Miñana, Brachaniec, Misz-Kennan, Niedźwiedzki and Wieslaw2018; Yuan et al., Reference Yuan, Liu, Chen, Xing, Spicer, Chen, Them, Wang, Li, Guo and Zhang2023). These alterations reverberated across climate systems and terrestrial ecosystems with some studies associating early lands plants as a potential catalyst of the Late Ordovician Mass Extinction (LOME) and the facilitators for the transition of arthropods from aquatic to terrestrial environments (Garwood & Edgecombe, Reference Garwood and Edgecombe2011; Kenrick et al., Reference Kenrick, Wellman, Schneider and Edgecombe2012; Dunlop et al., Reference Dunlop, Scholtz, Selden, Minelli, Boxshall and Fusco2013; Lenton et al., Reference Lenton, Dahl, Daines, Mills, Ozaki, Saltzman and Porada2016; Wallace et al., Reference Wallace, Shuster, Greig, Planavsky and Reed2017; Dahl &Arens, Reference Dahl and Arens2020). Additionally, previous studies linking the colonization of land plants to the Hirnantian glaciation – triggering the onset of LOME – demonstrates the necessity for more precise dating of terrestrial plants during the Ordovician period to refine the accuracy of phylogenetic trees, biogeochemical cycles and extinction models (Lenton et al., Reference Lenton, Crouch, Johnson, Pires and Dolan2012). The actual time at which these first vascular plant fossils appeared is often hard to determine, because it is difficult to correlate biostratigraphy from lake, river and coastal environment deposits in which they occur, with the standard marine-based geological time scale (Finney & Chen, Reference Finney and Chen1990; Pogson, Reference Pogson2009; Brookfield et al., Reference Brookfield, Catlos and Suarez2021). Hence, a robust U-Pb dating approach becomes imperative.
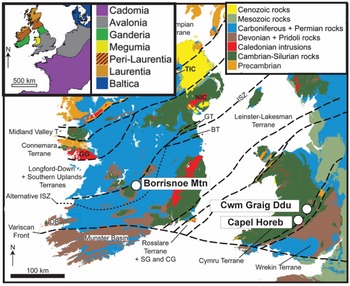
In this study, we present comprehensive data encompassing mineralogical X-ray diffraction (XRD), X-ray fluorescence (XRF) and the first detrital zircon U-Pb Laser Ablation-Inductively Coupled Plasma-Mass Spectrometry (LA-ICP-MS) dates from the three oldest sites (Middle to Late Silurian) containing Cooksonia macrofossils. These notable locations are situated in Ireland (Borrisnoe Mountain) and Wales (Capel Horeb and Cwm Graig Ddu, United Kingdom) (Fig. 2). We employ the well-established ‘Law of Detrital Zircon’ – a sedimentary rock is age-equivalent or younger than its youngest constituent zircon grains to establish maximum depositional ages (MDAs) for the siltstones bearing Cooksonia macrofossils within these localities, previously dated solely via biostratigraphy. These dates refine the timing of the earliest well-preserved vascular plant Cooksonia macrofossils.

Figure 2. Generalized terrane map of the UK and Ireland showing localities hosting Cooksonia’s earliest macrofossils (after Fairey et al., Reference Fairey, Kerrison, Meere, Mulchrone, Hofmann, Gärtner, Sonntag, Linnemann, Kuiper, Ennis and Mark2018).
2. Geological setting
2. a. Capel Horeb Quarry, Powys, Wales, UK
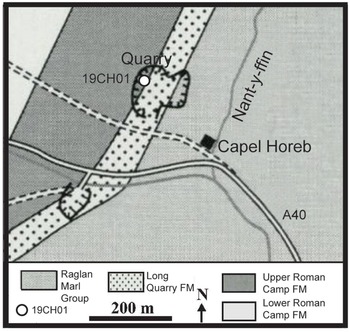
Capel Horeb Quarry in Powys, Wales, is the third-oldest known site harbouring Cooksonia’s macrofossils within the Upper Roman Camp Formation (Figs. 1a & 3; Table 1) (Specimen-NMW 69.64G, National Museum of Wales) (Cleal & Thomas, Reference Cleal and Thomas1995; Edwards & Richardson, Reference Edwards and Richardson2004). The Silurian stratigraphic section at this location was initially interpreted as Ludlovian by Potter & Price (Reference Potter and Price1965). Their description includes mudstone, shale, siltstone and sandstone. The stratigraphic sequence begins with the Black Cock Beds, estimated to be 870 ft.-thick, comprising silty mudstone, flaggy mudstone and siltstone and containing abundant marine fauna fossils. Subsequently, the Black Cock Beds are succeeded by the ∼810 ft.-thick Cwm Clyd Beds, which are characterized by siltstone, flaggy shaly sandstone and silty mudstone with a profusion of marine fauna fossils. Overlying the Cwm Clyd Beds are the Roman Camp Beds, further divided into the Lower Roman Camp Beds (360 ft. thick) and the Upper Roman Camp Beds (310 ft. thick). The Lower Roman Camp Beds are primarily composed of shaly siltstone, grey flaggy siltstone and sandstone, all bearing abundant marine fossils. In contrast, the Upper Roman Camp Beds are alternating grey and blue shaly siltstone and flaggy massive calcareous sandstone. Notably, Potter and Price (Reference Potter and Price1965) observed a decrease in fossils within the Upper Roman Camp Beds but reported the presence of fossilized plant remains.

Figure 3. Capel Horeb geologic map showing location of quarry and sample 19CH01. Modified from Lane (Reference Lane and Thomas2000).
The plant remains were initially found by Heard (Reference Heard1939) in the Upper Roman Camp Beds, and Edwards and Davies (Reference Edwards and Davies1976) described the Cooksonia macrofossils with an assigned Upper Ludlow (Ludfordian) age based on invertebrate and acritarchs biostratigraphy (Edwards and Rogerson, Reference Edwards and Rogerson1979; Edwards & Richardson, Reference Edwards and Richardson2004). However, there have also been reports of Pridoli age Cooksonia macrofossils in the sequence (Cleal & Thomas, Reference Cleal and Thomas1995). The plant fragments are most abundant in the grey blue flaggy siltstone beds within the Upper Roman Camp Beds. Based on the biostratigraphy, the depositional environment has been interpreted as an inshore marine environment (Cleal & Thomas, Reference Cleal and Thomas1995; Edwards & Richardson, Reference Edwards and Richardson2004).
2. b. Cwm Graig Ddu, Powys, Wales, UK
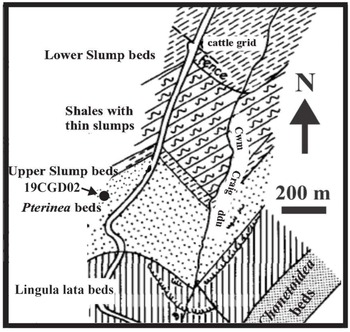
The Cwm Graig Ddu Quarry in Powys, Wales, is the second oldest known location containing Cooksonia’s macrofossils (Figs. 1a & 4; Table 1). Straw (Reference Straw1952) characterized the Silurian stratigraphic sequence at Cwm Graig Ddu as a succession of Lower and Upper Ludlow age shale, mudstone and siltstone. The sequence begins with the 1150 ft.-thick Lower Main Slump Group mudstone, followed by the 350 ft.-thick Shales with Thin Slumps, and the 140 ft-thick Upper Slump Group Shales. All these units are associated with the Lower Ludlow age graptolite Monograptus scanicus. The Upper Slump Group Shales are succeeded by the 1150 ft.-thick Pterinea teniustriata shale beds, the 750 ft.-thick Lingula lata shale beds and the 300 ft.-thick Chonetoidea grayi mudstone beds, attributed to the Lower Ludlow age Monograptus tumescens and Monograptus leintwardinensis graptolites. Straw (Reference Straw1952) documented marine fauna in the Lower Ludlow Pterinea beds, consisting of graptolites, crinoids, brachiopods, cephalopods, pelecypods, gastropods and plant fragment remains. The Pterinea beds are comprised of platy shale with its lower half exhibiting laminated shale and its upper half with interbedded shale with coarse lenticular silststone (Straw, Reference Straw1952). Cooksonia specimen-NMW 79.17G housed in the National Museum of Wales was located within the Pterinea Beds (Cleal & Thomas, Reference Cleal and Thomas1995; Edwards & Richardson, Reference Edwards and Richardson2004).

Figure 4. Cwm Graig Ddu geologic map showing location of sample 19CGD02. Modified from Straw (Reference Straw1952).
Cooksonia specimens from the Pterinea beds were described by Edwards et al. (Reference Edwards, Bassett and Rogerson1979) based on Straw’s (1952) detailed account (Edwards & Richardson, Reference Edwards and Richardson2004). The lithology and fossil content of the Pterinea beds and the surrounding area indicate a near shore deltaic depositional environment (Cleal & Thomas, Reference Cleal and Thomas1995). This setting is particularly suitable for understanding the transition of plants from marine to terrestrial environments (Edwards et al., Reference Edwards, Bassett and Rogerson1979). Cooksonia macrofossils were recovered from light grey micaceous flaggy siltstones and dark grey siltstones. The lithology in this area primarily consists of slumped fine-grained siltstones and sandstones, along with shales and mudstones. The presence of slumped beds within the Pterinea beds suggests a paleo-slope in the eastern Welsh basin, implying that the plant remains likely originated from the direction of the shelf and land (Edwards et al., Reference Edwards, Bassett and Rogerson1979).
Notably, the graptolite zones have been reevaluated since Straw’s (1952) interpretations, with the original Ludlow Monograptus tumescens zone being revised to the assessed Ludlow Gorstian age Monograptus incipiens zone (Edwards et al., Reference Edwards, Bassett and Rogerson1979; Edwards & Richardson, Reference Edwards and Richardson2004). It is worth mentioning that the currently established graptolite biozones have undergone a reassessment, leading to changes in nomenclature, including the removal of the Monograptus tumescens and Monograptus incipiens zones, and the renaming of Monograptus leintwardinensis to Saetograptus leintwardinensis (Melchin et al., Reference Melchin, Sadler, Cramer, Gradstein, Ogg, Schmitz and Ogg2020).
2. c. Borrisnoe Mountain, County Tipperary, Ireland
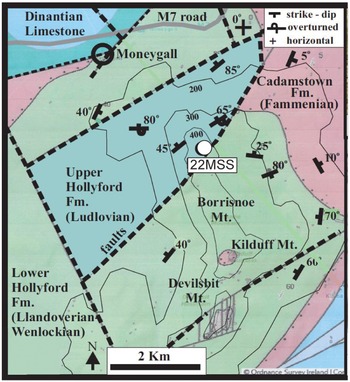
Borrisnoe Mountain in County Tipperary, Ireland, hosts the oldest macrofossils of the Cooksonia genus in the Hollyford Formation (Figs. 1a & 5; Table 1) (Edwards & Feehan Reference Edwards and Feehan1980; Edwards et al., Reference Edwards, Feehan and Smith1983; Edwards & Kenrick, Reference Edwards and Kenrick2015). Tectonism and fault systems in the area coupled with limited rock exposures led to considerable variations in stratigraphic and lithological descriptions of the region. Cope (Reference Cope1954, Reference Cope1959) initially divided the area’s stratigraphic sequences into two groups, the Lower Ludlow-age Cloncannon Formation defined by the Monograptus tumescens graptolite zone and the Upper Wenlock-age assemblage characterized by Cyrtograptus lundgreni. According to Cope’s (1959) interpretation, both assemblages share similar lithologies, consisting of mudstone, siltstone and greywacke. Siltstone constitutes approximately half of the Cloncannon Formation’s volume. Cope (Reference Cope1959) identified four types of siltstones facies, described as laminated, flaggy, graded-bed and micaceous. Plant remains were reported in the micaceous siltstones, suggesting a brackish water environment (Cope, Reference Cope1959).

Figure 5. Geologic and tectonic map of Borrisnoe Mountain in County Tipperary, Ireland, with location of analysed sample 22MSS (modified from Irish Geological Survey solid map and Cope, Reference Cope1959).
Doran (Reference Doran1974) and Archer (Reference Archer1981) recognized a single formation at Borrisnoe Mountain and termed it the Hollyford Formation. Cope’s (1959) Cloncannon Formation occurs in an isolated fault-bounded block and is younger than the rest of the sediments, and we use the term upper Hollyford Formation for the Ludlow age sediments, and lower Hollyford Formation for the mass of Wenlock age sediments surrounding them. According to Doran (Reference Doran1974), the lithological composition of the Hollyford Formation consists of 60% greywacke, 25% mudstone and 15% laminated siltstone, indicating a marine environment associated with the Upper Wenlock age based on Cyrtograptus lundgreni. Archer (Reference Archer1981) described the Hollyford Formation as being characterized by extensive turbidite deposits comprising mudstone, sandstone and conglomerage, indicative of a marine fan environment.
Edwards and Feehan (Reference Edwards and Feehan1980) described and attributed the Cooksonia fossil (Specimen-TCD 22944, Geological Museum, Trinity College Dublin) from the micaceous siltstones to the Cloncannon Formation, which in this study will be attributed to the Hollyford Formation. The unit containing the plant fossils within the Hollyford Formation has been reassessed to the graptolite zone of Monograptus ludensis, placing it in the mid-Silurian (Late Wenlock ∼ 427.4 – 430.5 Ma) (Edwards & Feehan, Reference Edwards and Feehan1980). As described by Edwards et al. (Reference Edwards, Feehan and Smith1983), the depositional environment of the Hollyford Formation is a marine succession within a submarine fan, featuring turbiditic sandstones, conglomerates, mudstones, and laminated siltstones with sparse volcanic clasts (Edwards & Feehan, Reference Edwards and Feehan1980; Edwards et al., Reference Edwards, Feehan and Smith1983).
3. Methods
This study involved the data acquisition of XRF, XRD and U-Pb detrital zircon dates from three siltstone samples (22MSS, 19CGD02, 19CH01), bearing the three earliest-recorded Cooksonia macrofossils found in sites across Wales and Ireland. Sample 22MSS is sourced from the Hollyford Formation, situated in Borrisnoe Mountain, County Tipperary, Ireland (52°51’15.82” N, 7°55’16.10” W), linked to specimen TCD 22944 from the Geological Museum at Trinity College, Dublin, Ireland (Edwards & Feehan,Reference Edwards and Feehan1980; Edwards et al., Reference Edwards, Feehan and Smith1983). Sample 19CGD02 originates from the Pterinea Beds within Cwm Graig Ddu Quarry, Powys, Wales (52°6’49.50” N, 3°31’14.70” W), correlated with specimen NMW 79.17G from the National Museum of Wales (Edwards et al., Reference Edwards, Bassett and Rogerson1979; Cleal & Thomas, Reference Cleal and Thomas1995; Edwards & Richardson, Reference Edwards and Richardson2004). Sample 19CH01 is from the Upper Roman Camp Beds at Capel Horeb Quarry, Powys, Wales (51°58’40.98” N, 3°41’1.20” W), associated with specimen NMW 69.64G from the National Museum of Wales (Edwards & Rogerson, Reference Edwards and Rogerson1979; Edwards et al., Reference Edwards, Bassett and Rogerson1979; Edwards, Reference Edwards1982; Cleal & Thomas, Reference Cleal and Thomas1995; Edwards & Richardson, Reference Edwards and Richardson2004).
Traditional heavy mineral separation techniques were used following modified methods by Garza et al. (Reference Garza, Catlos, Chamberlain, Suarez, Brookfield, Stockli and Batchelor2023). All zircon grains were annealed for 50 hours at 850 °C to repair fission tracks, minor radiation damage, reduce isotopic fractionation and improve accuracy of LA-ICP-MS data (Mattinson, Reference Mattinson2005; Allen & Campbell Reference Allen and Campbell2012). A total of 787 zircon grains were mounted on double-sided tape and inspected with backscatter electron (BSE) imaging using a JEOL Scanning Microscope at the University of Texas at Austin (UT Austin), GeoMaterials Characterization and Imaging facility (GeoMatCI). Following imaging, the zircon grains were dated using Element2 High Resolution (HR)-LA-ICP-MS in the Geo-Thermochronology Laboratory at UT Austin. The instrument uses an Eximer (192 nm) laser ablation system and obtains isotopic measurements using ion counting. A dry ablated aerosol is introduced to the instrument by pure He carrier gas containing the desired isotopic analytes, which for this study consists of 238U, 235U, 232Th, 206Pb, 207Pb and 208Pb. Each analysis consists of a 2-pulse cleaning ablation, a background measurement taken with the laser off, a 30-second measurement with the laser firing and a 30-second cleaning cycle. The laser beam was 20 µm in diameter to analyses grains as small as 50 µm. Elemental isotopic fractionation of Pb and Pb/U isotopes was corrected by interspersed analyses of primary and secondary zircon standards with known ages (GJ1 and Plesovice references) (Jackson et al., Reference Jackson, Pearson, Griffin and Belousova2004; Sláma et al., Reference Sláma, Košler, Condon, Crowley, Gerdes, Hanchar, Horstwood, Morris, Nasdala, Norberg, Schaltegger, Schoene, Tubrett and Whitehouse2008) The typical ratio of unknown standards measurements was 4:1 or 5:1. Systematic uncertainties resulting from calibration corrections are usually 1-2% for 206Pb/207Pb and 206Pb/238U. Fully propagated errors utilized for U-Pb zircon dates. Pb values are reported as total Pb without any correction for potential common 204Pb due to isobaric interferences with 204Hg. Iolite 4 software was used to process and reduce data analyses, correct instrument drift and downhole fractionation (https://iolite-software.com).
Statistical values, MDAs and figures were produced by Isoplot/Ex, Densityplotter and detritalPy with a bandwidth of ten (Ludwig, Reference Ludwig2008; Vermeesch, Reference Vermeesch2012; Sharman et al., Reference Sharman, Sharman and Sylvester2018). The LA-ICP-MS U-Pb dates underwent two interpretations. One approach involved applying a 206Pb/238U vs 207Pb/235U and 206Pb/238U vs 206Pb/207Pb 10% discordance filter across the entire dataset, reducing the number of dates from 787 to 378. Conversely, the other method entailed employing the 207Pb method as the common Pb correction for the 238U-206Pb dates up to 900 Ma for a total of 553 zircon dates. The 207Pb correction method removes common Pb by utilizing the measured 207Pb/206Pb ratio to project any data point to concordia. This method assumes that the sample’s radiogenic Pb component from the U-Pb data are concordant (Andersen, Reference Andersen2002; Chew et al., Reference Chew, Sylvester and Tubrett2011; White & Ireland, Reference White and Ireland2012; Chew et al., Reference Chew, Petrus and Kamber2014).
For the determination of the youngest single grain (YSG) MDA, calculations were based on the youngest zircon date derived from the 10% discordance filter dataset (Ludwig & Mundil, Reference Ludwig and Mundil2002). Meanwhile, the youngest cluster of 3+ grains (YC2σ+3) was determined by computing the weighted mean of the youngest zircon grain cluster consisting of three or more grains, which overlapped at 2σ uncertainty from the 10% discordance filter dataset (Dickinson & Gehrels, Reference Dickinson and Gehrels2009). Moreover, the youngest mode weighted mean (YMWM) was calculated following the method outlined by Tian et al. (Reference Tian, Fan, Valencia, Chamberlain, Waite, Stern and Loocke2022). This involved using LA-ICP-MS zircon dates that constituted the youngest age mode from a kernel density estimation (KDE) peak, calculating a weighted mean of more than three grains overlapping at 2σ uncertainty, with an approximate MSWD of 1 (Garza et al., Reference Garza, Catlos, Chamberlain, Suarez, Brookfield, Stockli and Batchelor2023). In this study, the YMWM was computed from 238U-206Pb dates up to 900 Ma using two different approaches: with a 10% discordance filter, and with a common Pb correction. Additionally, the YMWM was determined with three different KDE bandwidths (8, 10 and 12), while maintaining a consistent bin size of 5.
In addition to the zircon dates, whole rock mineralogical (XRD data) were obtained from all dated samples at the GeoMatCl facility at UT Austin. Whole rock samples were manually homogenized, ground and sieved to a 250 µm mesh size. XRD analyses were performed using a Bruker D8 instrument equipped with Cu Kα radiation and a nickel filter, along with a LYNXEYE solid-state detector. The analyses were carried out at a voltage of 45 kV and a current of 40 mA, employing a 2θscan axis ranging from 3° to 70°, with step increments of .0195° (2θ) and a duration of 1 s per step. Whole rock X-ray patterns were determined through Rietveld refinement utilizing Bruker TOPAS 4.2 software.
Whole rock elemental compositions were obtained using a portable XRF instrument at the University of Texas at Austin following the method of Rowe et al. (Reference Rowe, Hughes and Robinson2012). Samples were grounded for homogenization, pressed into a pellet and analysed using a Bruker S1 Titan 800 ED-XRF (portable XRF) instrument equipped with Rh X-ray tube. Analyses consisted of 15 Kv excitation voltage for major elements for 30 seconds and 50 Kv excitation voltage for trace elements for 60 seconds. XRF analyses were calibrated modifying the Bruker MudRock Air calibration and generating our own clastic rock standards. Our references consist of five international commercially available accepted standards (SBC-1, SGR-1b, SCo-2, ShBOQ-1 and SRM 70b) and five siltstone/sandstone internal standards. Elemental tectonic setting discrimination diagrams for siliciclastic sediments plotted after Roser & Korsh (Reference Roser and Korsch1986), Verma & Armstron-Altrin (Reference Verma and Armstrong-Altrin2013), Sahraeyan et al. (Reference Sahraeyan, Seif, Haddad, Mohammadzadeh and Ramkumar2015) and Rollinson & Pease (Reference Rollinson and Pease2021).
4. Results
4. a. Capel Horeb Quarry, Powys, Wales, UK
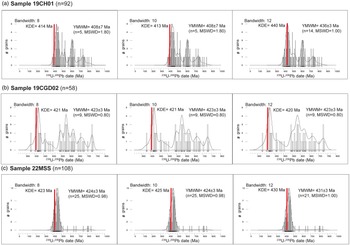
Sample 19CH01 obtained from the Upper Roman Camp Formation produced a total of 126 zircon dates post-application of a ≤ 10% discordance filter. These dates span from the Precambrian to the Devonian and serve to determine an MDA for the blue flaggy siltstone beds within the Upper Roman Camp Formation (Fig. 6a; supplementary table S1). The youngest MDA estimate is the YSG date of 385 ± 13 Ma (10% disc), while the YC2σ + 3 estimation presents a date of 402 ± 6 Ma (n = 5, MSWD = 1.80). The YMWM with various bandwidths provides dates of 408 ± 7 Ma (n = 5, MSWD = 1.80, bandwidth = 8), 408 ± 7 Ma (n = 5, MSWD = 1.80, bandwidth = 10) and 436 ± 3 Ma (n = 14, MSWD = 1.00, bandwidth = 12) (Fig. 7a).

Figure 6. Samples19CH01, 19CGD02 and 22MSS kernel density estimation (KDE) distribution plots. (a) LA-ICP-MS 238U-206Pb and 207Pb-206Pb zircon dates with (±2σ) uncertainties from 10% discordance filter dataset. Maximum depositional age (MDA) methods (youngest single grain [YSG]; youngest cluster of three or more grains overlapping within 2σ uncertainty [YC2σ+3]). (b) LA-ICP-MS 238U-206Pb dates with (±2σ) uncertainties from common Pb-corrected (207Pb method) dataset for zircon grains up to 900 Ma. Colours represent the period (pink: Precambrian; light green: Cambrian; dark green: Ordovician; teal blue: Silurian; orange: Devonian).

Figure 7. LA-ICP-MS 238U-206Pb zircon dates with (±2σ) uncertainties kernel density estimation (KDE) distribution plots up to 900 Ma from 10% discordance filter dataset with various bandwidth values (8, 10, 12) and consistent bin size of 5. Red line represents KDE peak date utilized for youngest mode weighted mean (YMWM) MDA calculation. (a) Sample 19CH01. (b) Sample 19CGD02 (c) sample 22MSS.
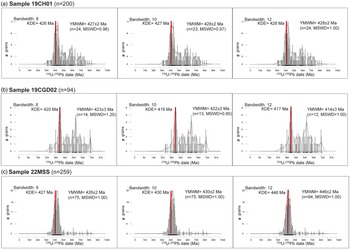
Utilizing the common Pb correction for the 238U-206Pb dates up to 900 Ma presents a total of 200 zircon dates spanning from Precambrian to Devonian (Fig. 6b). The YMWM with various bandwidths provides dates of 427 ± 2 Ma (n = 24, MSWD = 0.96, bandwidth = 8), 428 ± 2 Ma (n = 23, MSWD = 0.97, bandwidth = 10) and 428 ± 2 Ma (n = 24, MSWD = 1.00, bandwidth = 12) (Fig. 8a). The elemental (XRF) analyses of sample 19CH01 exhibit a major element composition marked by high Si, intermediate Al, and low Ca and Fe, characteristic of siltstone and sandstone lithological facies (supplementary table S2). Mineralogical (XRD) analyses support this observation, demonstrating a composition involving quartz, K-feldspar, plagioclase, chlorite, kaolinite and illite/mica. The corresponding weight percent quantification values are outlined in (Table 2 ).

Figure 8. LA-ICP-MS 238U-206Pb zircon dates with (±2σ) uncertainties kernel density estimation (KDE) distribution plots up to 900 Ma from common Pb-corrected (207Pb method) dataset with various bandwidth values (8, 10, 12) and consistent bin size of 5. Red line represents KDE peak date utilized for youngest mode weighted mean (YMWM) MDA calculation. (a) Sample 19CH01. (b) Sample 19CGD02 (c) sample 22MSS.
Table 2. X-ray diffraction (XRD) mineralogical analyses of this study’s samples

4. b. Cwm Graig Ddu, Powys, Wales, UK
Sample 19CGD02 obtained from the Pterinea Beds yielded a total of 65 zircon dates post-application of the ≤ 10% discordance filter. These zircon dates span from the Precambrian to the Devonian for the micaceous flaggy siltstone beds within the Pterinea Beds (Fig. 6a; supplementary table S1). The youngest estimate for this sample is the YSG date of 377 ± 8 Ma (6% disc) and the YC2σ + 3 estimation presents a date of 412 ± 6 (n = 9, MSWD = 2.20). The YMWM with various bandwidths provides dates of 423 ± 3 Ma (n = 9, MSWD = 0.80, bandwidth = 8), 423 ± 3 Ma (n = 9, MSWD = 0.80, bandwidth = 10) and 423 ± 3 Ma (n = 9, MSWD = 0.80, bandwidth = 12) (Fig. 7b).
Utilizing the common Pb correction for the 238U-206Pb dates up to 900 Ma presents a total of 94 zircon dates spanning from Precambrian to Devonian (Fig. 6b). The YMWM with various bandwidths provides dates of 423 ± 3 Ma (n = 14, MSWD = 1.20, bandwidth = 8), 422 ± 2 Ma (n = 13, MSWD = 0.80, bandwidth = 10) and 414 ± 3 Ma (n = 12, MSWD = 1.00, bandwidth = 12) (Fig. 8b).The elemental (XRF) assessments for sample 19CGD02 also depict a major element composition featuring high Si, intermediate Al, and low Ca and Fe, consistent with the siltstone and sandstone lithological facies (supplementary table S2). The mineralogical (XRD) analyses substantiate these findings, revealing a composition akin to quartz, K-feldspar, plagioclase, chlorite, kaolinite and illite/mica, with corresponding weight percent quantification values presented in (Table 2).
4. c. Borrisnoe Mountain, County Tipperary, Ireland
Sample 22MSS obtained from the Hollyford Formation yielded a total of 187 zircon dates post-application of a ≤ 10% discordance filter. These zircon dates span from the Precambrian to the Devonian age range for the micaceous siltstone bed within the Hollyford Formation (Fig. 6a; supplementary table S1). Among the estimations derived from the U-Pb LA-ICP-MS dates for sample 22MSS, the youngest estimate is the YSG date of 377 ± 13 Ma (4% disc) and the YC2σ + 3 estimation presents a date of 390 ± 13 (n = 4, MSWD= 2.00). Additionally, the YMWM with various bandwidths provides dates of 424 ± 3 Ma (n = 25, MSWD = 0.98, bandwidth = 8), 424 ± 3 Ma (n = 25, MSWD= 0.98, bandwidth= 10) and 431 ± 3 Ma (n = 21, MSWD = 1.00, bandwidth = 12) (Fig. 7c).
Utilizing the common Pb correction for the 238U-206Pb dates up to 900 Ma yields a total of 259 zircon dates spanning from Precambrian to Devonian. (Fig. 6b). The YMWM with various bandwidths provides dates of 426 ± 2 Ma (n = 75, MSWD = 1.00, bandwidth = 8), 430 ± 2 Ma (n = 75, MSWD = 1.00, bandwidth = 10) and 446 ± 2 Ma (n = 94, MSWD = 1.00, bandwidth = 12) (Fig. 8c). Elemental analyses (XRF) of sample 22MSS reveal a major element composition characterized by high Si, intermediate Al, and low Ca and Fe, indicative of siltstone and sandstone lithological facies (supplementary table S2). Mineralogical analyses (XRD) align with the elemental data, confirming a composition comprising quartz, K-feldspar, plagioclase, chlorite, kaolinite and illite/mica. Corresponding weight percent quantification values are detailed in (Table 2).
5. Discussion
5. a. Geochemistry of tectonic setting sediment provenance
The closure of the Iapetus Ocean resulted from the collision between Laurentia, Baltica and Avalonia during the Early Ordovician to the Late Silurian, leading to significant tectonism and the formation of the Caledonian Mountains (Chew & Strachan, Reference Chew and Strachan2014; Torsvik & Cocks, Reference Torsvik and Cocks2016; Golanka et al., Reference Golonka, Porębski and Waśkowska2023; Garza et al., Reference Garza, Catlos, Chamberlain, Suarez, Brookfield, Stockli and Batchelor2023). Deposits containing Cooksonia macrofossils, investigated in this study and located in Wales and Ireland, originated from island arcs in the Early Silurian, transitioning to volcanic and plutonic sources from an active continental margin by the Late Silurian (Fig. 9) (Chew & Strachan, Reference Chew and Strachan2014; Torsvik & Cocks, Reference Torsvik and Cocks2016; Golanka et al., 2023).

Figure 9. Paleogeographic reconstruction during Early to Late Silurian (after Torsvik & Cocks, Reference Torsvik and Cocks2016 and Golanka et al., 2023). During Early Silurian (435 Ma) closing of the Iapetus Ocean forms volcanic arcs (fore-arc and back-arc) near subduction margins of Laurentia, Baltica and Avalonia, producing widespread tectonic activity (Chew & Strachan, Reference Chew and Strachan2014; Garza et al., Reference Garza, Catlos, Chamberlain, Suarez, Brookfield, Stockli and Batchelor2023). By Late Silurian (425 Ma), Caledonian Mountains are formed near an active continental margin producing substantial volcanic activity (Chew & Strachan, Reference Chew and Strachan2014; Torsvik & Cocks, Reference Torsvik and Cocks2016; Golanka et al., 2023).
The XRF elemental analyses conducted in this study on three siltstone samples from distinct localities validate the paleo-continental reconstructions (Fig. 10). Our findings reveal significant island arc sediment contributions for samples 22MSS and 19CGD02, collected from Borrisnoe Mountain (Ireland) and Cwm Graig Ddu (Wales). Conversely, sample 19CH01, originating from Capel Horeb (Wales), exhibits predominant sediment input from an active continental margin.

Figure 10. Tectonic setting discrimination diagram based on XRF major element composition (after Roser & Korsch, Reference Roser and Korsch1986 and Sahraeyan et al. Reference Sahraeyan, Seif, Haddad, Mohammadzadeh and Ramkumar2015).
The XRD mineralogical analyses align with the typical siltstone mineralogy, predominantly composed of quartz and feldspars, with a noticeable clay component primarily represented by illite/mica (Table 2 ). This dataset is particularly valuable as it corresponds with previous lithological descriptions of rocks enclosing the Cooksonia macrofossils across the three different localities (Heard, Reference Heard1939; Straw, Reference Straw1952; Cope, Reference Cope1959; Potter & Price, Reference Potter and Price1965; Edwards & Davies, Reference Edwards and Davies1976; Edwards et al., Reference Edwards, Bassett and Rogerson1979; Edwards & Feehan, Reference Edwards and Feehan1980; Edwards et al., Reference Edwards, Feehan and Smith1983). The main distinctions among localities lie in the mineral composition of tectosilicates and clays. Borrisnoe Mountain (Ireland) and Cwm Graig Ddu (Wales) exhibit a very similar mineral composition in tectosilicates and clays, supporting our elemental data indicating a comparable sediment source (Table 2). In contrast, Capel Horeb (Wales) contains a higher tectosilicate content and significantly lower clay content, suggesting a distinct sediment source.
5. b. Implications for assigning MDAs
MDAs derived from detrital zircons, typically adhere to the ‘law of detrital zircon’, a principle asserting that a geological formation or rock bed cannot be older than its youngest constituent, but can be younger. This principle sets a threshold age limit for MDAs (Gehrels, Reference Gehrels2014; Herriott et al., Reference Herriott, Crowley, Schmitz, Wartes and Gillis2019; Sharman & Malkowski, Reference Sharman and Malkowski2020; Brookfield et al., Reference Brookfield, Catlos and Suarez2021). When evaluating MDAs for the samples in this study using the 10% discordance-filtered U-Pb dates, interpretations using the YSG and youngest cluster of three or more grains overlapping at two sigma (YC2σ + 3) methods reveal U-Pb dates significantly younger than their presently assessed ages for all samples (Fig. 6a). However, MDA methodologies such as the YSG and YC2σ + 3 methods might be considered less conservative, as they rely on the youngest concordant zircons or clusters, potentially yielding considerably younger dates than the true depositional age (TDA) due to potential Pb loss or systematic uncertainties (Coutts et al., Reference Coutts, Matthews and Hubbard2019; Andersen et al., Reference Andersen, Elburg and Magwaza2019; Herriott et al., Reference Herriott, Crowley, Schmitz, Wartes and Gillis2019; Garza et al., Reference Garza, Catlos, Chamberlain, Suarez, Brookfield, Stockli and Batchelor2023; Sharman and Malkowski, Reference Sharman and Malkowski2024).
The YMWM dates derived from the 10% discordance-filtered U-Pb dates generally match the biostratigraphy age within a certain range of uncertainty, except for sample 19CH01 (Fig. 7). In this particular case, the KDE plots produced YMWM dates that either skewed too young or too old compared to the biostratigraphy age. Despite the current assigned biostratigraphy age for sample 19CH01 being Ludfordian–Gorstian (∼423–426 Ma), the KDE with a bandwidth of 8 generated a younger YMWM date of 408 ± 7 Ma (n = 5, MSWD = 1.80), while the KDE with a bandwidth of 10 also yielded a YMWM date of 408 ± 7 Ma (n = 5, MSWD = 1.80) (Fig. 7a). Conversely, the KDE with a bandwidth of 12 resulted in an older YMWM date of 436 ± 3 Ma (n = 14, MSWD = 1.00). This discrepancy in YMWM dates seems to be driven by Pb loss and the variability in selected bandwidths used for KDE plots.
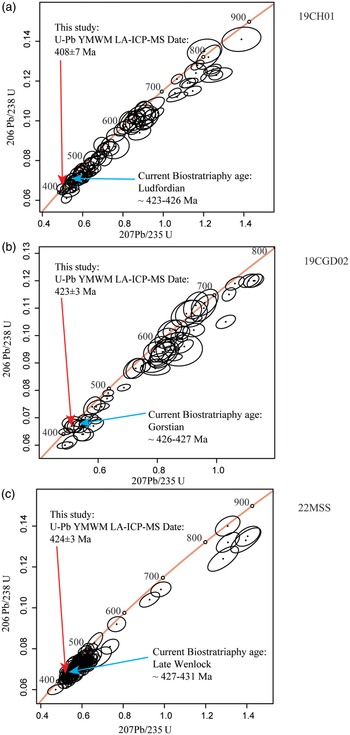
The consistently younger dates obtained through the YSG, YC2σ + 3 and YMWM methods, in comparison to their assigned biostratigraphic age, may be attributed to significant disturbances in the U-Pb system within the samples. As outlined in the methods section, only 49% of the total zircons (378 out of 787) met the criteria for the 10% discordance filter, indicating Pb loss within the zircons across all three fossil sites. The concordia diagrams illustrate Pb loss within the three samples, especially the youngest grains (Fig. 11). Figure 9 illustrates substantial tectonic activity in the three localities due to the collision of Baltica, Avalonia and Laurentia, leading to the formation of the Caledonian Mountains from the Early Ordovician to the Late Silurian. This tectonic activity likely induced zircon metamictization and subsequent Pb loss (Chew & Strachan, Reference Chew and Strachan2014; Torsvik & Cocks, Reference Torsvik and Cocks2016; Glonka et al., 2023; Garza et al., Reference Garza, Catlos, Chamberlain, Suarez, Brookfield, Stockli and Batchelor2023). Additionally, during the Late Devonian through the Early Carboniferous, the collision between Laurentia (now Laurussia) and Gondwana contributed to the formation of Pangea, potentially exacerbating the degree of metamictization in the already affected zircon grains (Blakey, Reference Blakey and Wong2003; Blakey, Reference Blakey, Fielding, Frank and Isbell2008).

Figure 11. 238U-206Pb LA-ICP-MS concordia diagrams reported as total Pb with 10% discordance filter for zircons younger than 900 Ma from this study’s localities in Ireland and Wales. (a) Sample 19CH01 from Capel Horeb, Wales, accepted biostratigraphy age from Edwards & Richardson (Reference Edwards and Richardson2004), Edwards & Kenrick (Reference Edwards and Kenrick2015). (b) Sample 19CGD02 from Cwm Graig Ddu, Wales, accepted biostratigraphy age from Edwards & Richardson (Reference Edwards and Richardson2004), Edwards & Kenrick (Reference Edwards and Kenrick2015). (c) Sample 22MSS from Borrisnoe Mountain, Ireland, accepted biostratigraphy age from Edwards & Feehan (Reference Edwards and Feehan1980), Edwards et al. (Reference Edwards, Feehan and Smith1983), Edwards & Kenrick (Reference Edwards and Kenrick2015), Salamon et al. (Reference Salamon, Gerrienne, Steemans, Gorzelak, Filipiak, Le Hérissé, Florentin, Cascales- Miñana, Brachaniec, Misz-Kennan, Niedźwiedzki and Wieslaw2018). Black ovals present LA-ICP-MS dates with 2σ uncertainty, blue arrow shows accepted biostratigraphy age and red arrow displays the YMWM MDA from dataset with 10% discordance filter.
According to Vermeesch (Reference Vermeesch2012), the choice of kernel in KDE plots is less critical compared to the bandwidth. Depending on the selected bandwidth, KDE diagrams may be oversmoothed or under-smoothed, potentially biasing MDA calculations and resulting in younger or older U-Pb dates. In datasets with limited available data, a larger bandwidth is employed, resulting in a smoother KDE plot. Conversely, in datasets with abundant data, a narrower bandwidth is utilized to maintain high resolution in KDE plots, ensuring that the peaks in the KDE distributions are not oversmoothed (Vermeesch, Reference Vermeesch2012). However, discrepancy arises from the non-standardized use of different bandwidth values across the geochronology community, where bandwidth value selection can vary between laboratories or individuals, leading to potential inconsistencies in sediment provenance results or MDA calculations.
When utilizing the common Pb-corrected (207Pb method) 238U-206Pb dates up to 900 Ma to compute YMWM dates with different bandwidths, we also observe variations in the YMWM dates (Fig. 8). The advantage of employing this common Pb-corrected method is its incorporation of more U-Pb dates into statistical calculations for MDAs that might have otherwise been discarded during the filtering process. However, since the common Pb correction does not filter based on discordance but rather shifts all 238U-206Pb zircon dates including the discordant grains with Pb-loss to concordia, we attribute the variation in YMWM dates to the different bandwidth values.
For instance, in the case of sample 19CH01, with a Ludfordian–Gorstian biostratigraphy age (∼423–426 Ma), the YMWM dates generated with bandwidths of 8, 10 and 12 all fell within the uncertainty range of the biostratigraphy age (Fig. 8a). The KDE with a bandwidth of 8 produced a YMWM date of 427 ± 2 Ma (n = 24, MSWD = 0.96), while the KDE with a bandwidth of 10 yielded a YMWM date of 428 ± 2 Ma (n = 23, MSWD = 0.97), and the KDE with a bandwidth of 12 resulted in a YMWM date of 428 ± 2 Ma (n = 24, MSWD = 1.00).
Sample 19CGD02, with a Gorstian biostratigraphy age (∼426–427 Ma), exhibited the most variability, with only the YMWM date generated using a bandwidth value of 8 matching the biostratigraphy age within uncertainty (Fig. 8b). The KDE with a bandwidth of 8 yielded a YMWM date of 423 ± 3 Ma (n = 14, MSWD = 1.20), while the KDE with a bandwidth of 10 resulted in a YMWM date of 422 ± 2 Ma (n = 13, MSWD = 0.80), and the KDE with a bandwidth of 12 produced a significantly younger YMWM date of 414 ± 3 Ma (n = 12, MSWD = 1.00).
Sample 22MSS, with a Homerian–Sheinwoodian biostratigraphy age (∼427–431 Ma), displayed some variability, with the YMWM dates generated using bandwidths of 8 and 10 matching the biostratigraphy age within uncertainty, while the YMWM date with a bandwidth of 12 resulted in a significantly older date (Fig. 8c). The KDE with a bandwidth of 8 yielded a YMWM date of 426 ± 2 Ma (n = 75, MSWD = 1.00), while the KDE with a bandwidth of 10 produced a YMWM date of 430 ± 2 Ma (n = 75, MSWD = 1.00), and the KDE with a bandwidth of 12 resulted in a significantly older YMWM date of 446 ± 2 Ma (n = 94, MSWD = 1.00).
Overall, we recognized that Pb loss may be at play when not using the common Pb correction, so to account for this, we thermally annealed our grains and applied a more stringent 10% discordance filter for one of our interpretation methods. Furthermore, we use a more conservative YMWM MDA interpretation instead of relying on the YSG and YC2σ + 3, which are more prone to erroneous interpretations in samples impacted by cryptic Pb loss. Discordance alone proves insufficient for identifying Pb loss in Phanerozoic LA-ICP-MS data, as the effects of Pb loss may be cryptic or occur shortly after crystallization, leading to U-Pb analyses shifting along concordia (Andersen et al., Reference Andersen, Elburg and Magwaza2019; Garza et al., Reference Garza, Catlos, Chamberlain, Suarez, Brookfield, Stockli and Batchelor2023; Sharman and Malkowski, Reference Sharman and Malkowski2024). This issue remains undetectable in relatively low-precision LA-ICP-MS analyses (Andersen et al., Reference Andersen, Elburg and Magwaza2019).
The YMWM MDA approach, coupled with the common Pb correction (207Pb method) and a bandwidth of 8, consistently produces dates that closely approximate the current assessed biostratigraphy ages for Borrisnoe Mountain, Ireland (sample 22MSS), Cwm Graig Ddu Quarry, Wales (sample 19CGD02), and Capel Horeb, Wales (sample 19CH01) (Table 3). We opted for the YMWM MDAs with a bandwidth of 8 due to its ability to generate KDE plots with higher resolutions that avoid oversmoothing KDE distribution peaks (Vermeesch, Reference Vermeesch2012). However, the discrepancies arising from the use of different bandwidth values for KDE plots, whether with discordance-filtered or common Pb-corrected data, require careful evaluation. Bandwidth value preferences vary among labs and individuals, potentially leading to MDA interpretation biases.
Table 3. Comparison of 238U-206Pb LA-ICP-MS youngest mode weighted mean (YMWM) MDA calculations using a 10% discordance filter, a common Pb correction (207Pb method) and various KDE bandwidth values (Bw) with assigned biostratigraphy age

Ultimately, the most accurate LA-ICP-MS MDA estimates appear consistent with U-Pb Chemical Abrasion-Isotope Dilution-Thermal Ionization Mass Spectrometry (CA-ID-TIMS) dates are derived from calculations involving averages rather than solely relying on the youngest grains (Herriott et al., Reference Herriott, Crowley, Schmitz, Wartes and Gillis2019; Tian et al., Reference Tian, Fan, Valencia, Chamberlain, Waite, Stern and Loocke2022; Garza et al., Reference Garza, Catlos, Chamberlain, Suarez, Brookfield, Stockli and Batchelor2023). CA-ID-TIMS is the most precise and accurate U-Pb dating technique, boasting ≤ 0.1% 2σ age precision (Mattinson, Reference Mattinson2005; Bowring et al., Reference Bowring, Schoene, Crowley, Ramezani and Condon2006; Schaltegger et al., Reference Schaltegger, Schmitt and Horstwood2015). This approach helps minimize the effects of potential Pb loss (Herriott et al., Reference Herriott, Crowley, Schmitz, Wartes and Gillis2019; Tian et al., Reference Tian, Fan, Valencia, Chamberlain, Waite, Stern and Loocke2022; Garza et al., Reference Garza, Catlos, Chamberlain, Suarez, Brookfield, Stockli and Batchelor2023). Given these considerations, we endorse the YMWM MDA method with the common Pb correction as the most suitable option for this study, given its alignment with CA-ID-TIMS results in other studies (Tian et al., Reference Tian, Fan, Valencia, Chamberlain, Waite, Stern and Loocke2022; Garza et al., Reference Garza, Catlos, Chamberlain, Suarez, Brookfield, Stockli and Batchelor2023). Moreover, it has previously demonstrated effectiveness in yielding optimal age results when applied to Early Paleozoic zircon grains (Tian et al., Reference Tian, Fan, Valencia, Chamberlain, Waite, Stern and Loocke2022; Garza et al., Reference Garza, Catlos, Chamberlain, Suarez, Brookfield, Stockli and Batchelor2023).
5. c. Implications for the refinement of Cooksonia ages
The determination of where and when the earliest vascular plants originated is critical for understanding the emergence of land biomes and the broader development of the biosphere. This knowledge is crucial for unraveling the adaptations that shaped vascular plants and understanding the intricate ecological interactions that facilitated the terrestrial colonization by the first arthropods (Shear & Selden, Reference Shear and Selden2001; Gerrienne et al., Reference Gerrienne, Servais and Vecoli2016; Brookfield et al., Reference Brookfield, Catlos and Suarez2021; Brookfield et al., Reference Brookfield, Catlos and Suarez2022; Dahl & Arens, Reference Dahl and Arens2020). Molecular phylogenies of vascular plants suggest that Cooksonia evolved during the Middle Cambrian (Rota-Stabelli et al., Reference Rota-Stabelli, Daley and Pisani2013; Morris et al., Reference Morris, Puttick, Clark, Edwards, Kenrick, Pressel, Wellman, Yang, Schneider and Donoghue2018). However, macrofossils of the earliest vascular plants are exclusively found in the Silurian period (Shear & Selden, Reference Shear and Selden2001; Brookfield et al., 2020; Dahl & Arens, Reference Dahl and Arens2020). The Silurian is recognized as the terrestrial equivalent of the Cambrian ‘explosion’ of marine faunas, marked by an increase in phenotypic diversity and the establishment of frameworks reminiscent of modern ecosystems (Bateman et al., Reference Bateman, Crane, DiMichele, Kenrick, Rowe, Speck and Stein1998; Benton & Emerson, Reference Benton and Emerson2007; Minter et al., Reference Minter, Buatois, Mángano, Davies, Gibling, MacNaughton and Labandeira2017).
Previous studies into the appearance of vascular plants on land have employed molecular clock phylogenetic models, microfossil records (trilete spores) and macrofossil records to understand vascular plants evolution and the timing of land colonization by Cooksonia (Edwards & Kenrick, Reference Edwards and Kenrick2015; Salamon et al., Reference Salamon, Gerrienne, Steemans, Gorzelak, Filipiak, Le Hérissé, Florentin, Cascales- Miñana, Brachaniec, Misz-Kennan, Niedźwiedzki and Wieslaw2018; Donoghue et al., Reference Donoghue, Harrison, Paps and Schneider2021). However, these prior assessments encounter discrepancies in ages between the micro- and macrofossil records which rely solely on biostratigraphy for calibration in phylogenetic models (Clarke et al., Reference Clarke, Warnock and Donoghue2011; Edwards & Kenrick, Reference Edwards and Kenrick2015; Morris et al., Reference Morris, Puttick, Clark, Edwards, Kenrick, Pressel, Wellman, Yang, Schneider and Donoghue2018; Servais et al., Reference Servais, Cascales-Miñana, Cleal, Gerrienne, Harper and Neumann2019). The primary issue is the exclusive reliance on biostratigraphy for dating both micro- and macrofossils; while a decent starting point, U-Pb dates provides a numerical assessment. Second, there exists a 20-million-year gap between trilete spores dated to a Late Ordovician (Katian) age and the oldest macrofossil assigned to a Middle Silurian (Wenlock) age, creating discordance between the micro- and macrofossil records (Edwards & Kenrick, Reference Edwards and Kenrick2015; Salamon et al., Reference Salamon, Gerrienne, Steemans, Gorzelak, Filipiak, Le Hérissé, Florentin, Cascales- Miñana, Brachaniec, Misz-Kennan, Niedźwiedzki and Wieslaw2018). Third, phylogenetic models calibrated by fossils contradict both the micro- and macrofossil records, estimating an initial appearance during the Middle Cambrian (Clarke et al., Reference Clarke, Warnock and Donoghue2011; Morris et al., Reference Morris, Puttick, Clark, Edwards, Kenrick, Pressel, Wellman, Yang, Schneider and Donoghue2018; Servais et al., Reference Servais, Cascales-Miñana, Cleal, Gerrienne, Harper and Neumann2019). Our study establishes MDAs using detrital zircon 238U-206Pb and 207Pb-206Pb LA-ICP-MS dates from the three oldest sites containing Cooksonia macrofossils in Wales and Ireland.
The existing order and age assessment of the oldest sites harbouring Cooksonia macrofossils characterizes Borrisnoe Mountain, Ireland, as the oldest, assigned a Middle Silurian (Late Wenlock) age, followed by Cwm Graig Ddu Quarry, Wales, designated with a Late Silurian (Gorstian) age, and subsequently Capel Horeb, Wales, attributed to a Late Silurian (Ludfordian) age (Table 1 ) (Edwards & Davies, Reference Edwards and Davies1976; Edwards et al., Reference Edwards, Bassett and Rogerson1979; Edwards & Rogerson, Reference Edwards and Rogerson1979; Edwards & Feehan, Reference Edwards and Feehan1980; Edwards et al., Reference Edwards, Feehan and Smith1983; Edwards & Richardson, Reference Edwards and Richardson2004). However, our recent U-Pb MDA dating outcomes refine these former age assessments. Our new findings reveal that utilizing the YMWM MDA with common Pb-corrected 238U-206Pb zircon dates indicates that both Borrisnoe Mountain and Capel Horeb host the oldest Cooksonia macrofossils, with both localities exhibiting a Gorstian–Homerian age for the sediments containing these macrofossils (Fig. 12). Specifically, the Borrisnoe Mountain sediments of the Hollyford Formation yield an MDA of 426 ± 2 Ma, while the Capel Horeb sediments of the Upper Roman Camp Formation provide an MDA of 427 ± 2 Ma (Fig. 13). However, due to the uncertainty inherent in U-Pb LA-ICP-MS analyses, there remains a possibility that the Cooksonia macrofossil from Capel Horeb could indeed be the oldest. Therefore, further CA-ID-TIMS analyses are imperative to obtain more precise and accurate U-Pb dates, allowing us to differentiate between these two sites definitively. In contrast, the Cwm Graig Ddu Quarry site presents a Pridoli-Ludlow MDA age of 423 ± 3 Ma, making it the youngest locality bearing Cooksonia macrofossils among the three sites examined in this study (Fig. 13). Notably, this MDA age aligns consistently within uncertainty with its assigned Ludfordian biostratigraphy age (Edwards & Davies, Reference Edwards and Davies1976; Edwards & Rogerson, Reference Edwards and Rogerson1979; Edwards & Richardson, Reference Edwards and Richardson2004; Edwards & Kenrick, Reference Edwards and Kenrick2015).

Figure 12. Stratigraphic columns displaying previous and this study’s chronostratigraphic assessment. (a) Generalized stratigraphic column of Hollyford Formation at Borrisnoe Mountain, Ireland (Cope, Reference Cope1959; Edwards & Feehan, Reference Edwards and Feehan1980; Edwards et al., Reference Edwards, Feehan and Smith1983). LA-ICP-MS YMWM dates derive from sample 22MSS. (b) Stratigraphic column of Pterinea Shale Beds and Lingula lata Shale at Cwm Graig Ddu, Wales (Straw, Reference Straw1952; Edwards et al., Reference Edwards, Bassett and Rogerson1979). LA-ICP-MS YMWM date is from sample 19CGD02. (C) Stratigraphic column of Cwm Clyd Beds and Roman Camp Beds at Capel Horeb, Wales (Heard, Reference Heard1939; Potter & Price, Reference Potter and Price1965; Edwards & Davies, Reference Edwards and Davies1976; Edwards & Rogerson, Reference Edwards and Rogerson1979). LA-ICP-MS YMWM date derives from sample 19CH01.

Figure 13. Geological time scale with Cooksonia’s first appearance datum (FAD) micro- and macrofossil record. Current assigned biostratigraphy age (darker font) (from Edwards et al., Reference Edwards, Bassett and Rogerson1979; Edwards & Rogerson., Reference Edwards and Rogerson1979; Edwards & Feehan, Reference Edwards and Feehan1980; Edwards et al., Reference Edwards, Feehan and Smith1983; Edwards & Richardson, Reference Edwards and Richardson2004; Edwards & Kenrick, Reference Edwards and Kenrick2015; Salamon et al., Reference Salamon, Gerrienne, Steemans, Gorzelak, Filipiak, Le Hérissé, Florentin, Cascales- Miñana, Brachaniec, Misz-Kennan, Niedźwiedzki and Wieslaw2018). This study’s 238U-206 Pb (common Pb-corrected) youngest mode weighted mean (YMWM) are shown in colour font and boxes. Borrisnoe Mountain (Ireland) in red; Cwm Graig Ddu (Wales) in blue; Capel Horeb (Wales) in green.
The MDAs obtained for the localities investigated in this study are crucial in unraveling the evolution of vascular plants and providing valuable insights into broader patterns of biological evolution, including diversification and the environmental factors influencing their development (Kenrick et al., Reference Kenrick, Wellman, Schneider and Edgecombe2012; De Vries & Archibald, 2017). Understanding the age and evolution of Cooksonia plays a pivotal role in reconstructing paleoenvironments and offers valuable perspectives on ancient plant life, contributing significantly to changes in biogeochemical cycles, paleoclimates and ecosystems (Wellman, Reference Wellman2010; Clarke et al., Reference Clarke, Warnock and Donoghue2011; Lenton et al., Reference Lenton, Crouch, Johnson, Pires and Dolan2012; Yuan et al., Reference Yuan, Liu, Chen, Xing, Spicer, Chen, Them, Wang, Li, Guo and Zhang2023). Refined maximum ages are indispensable for improving phylogenetic models and reconstructing plant evolution and the tree of life (Lenton et al., Reference Lenton, Crouch, Johnson, Pires and Dolan2012). The U-Pb MDAs act as a foundational tool for more accurately calibrating molecular clocks, assisting in the estimation of divergence times for different plant lineages.
6. Conclusions
The aim of this study is to reassess the chronological placement of the Cooksonia genus by employing U-Pb dating on the oldest macrofossils discovered in Wales (UK) and Ireland. The evolution of vascular plants on land has been a dynamic process marked by numerous innovations and adaptations, playing a pivotal role in shaping terrestrial environments globally (Kenrick et al., Reference Kenrick, Wellman, Schneider and Edgecombe2012; DeVries & Archibald, 2017). Beyond transforming landscapes, these plants have exerted a profound influence on the evolution of other organisms and ecosystems, establishing themselves as a critical component of the planet’s biodiversity and ecological complexity Garwood & Edgecombe, Reference Garwood and Edgecombe2011; Kenrick et al., Reference Kenrick, Wellman, Schneider and Edgecombe2012; Dunlop et al., Reference Dunlop, Scholtz, Selden, Minelli, Boxshall and Fusco2013; Lenton et al., Reference Lenton, Dahl, Daines, Mills, Ozaki, Saltzman and Porada2016; Wallace et al., Reference Wallace, Shuster, Greig, Planavsky and Reed2017; Dahl &Arens, Reference Dahl and Arens2020).
This study presents the first 238U-206Pb and 207Pb-206Pb zircon dates for the earliest recognized Cooksonia macrofossils, adjusting previous age interpretations reliant on biostratigraphy. By leveraging detrital zircon U-Pb dating and the YMWM MDA approach, we systematically refined the age assessments of key fossil sites harbouring Cooksonia macrofossils in Ireland and Wales. Our findings present Gorstian–Homerian maximum ages for both the Borrisnoe Mountain (Ireland) and Capel Horeb (Wales) macrofossils, with maximum ages of 426 ± 2 Ma and 427 ± 2 Ma, respectively (Fig. 13). For Cwm Graig Ddu (Wales), our Cooksonia’s (Pridoli-Ludlow) MDA of 423 ± 3 Ma aligns with its assigned biostratigraphy (Gorstian) age within the margin of uncertainty. Future U-Pb CA-ID-TIMS analyses are imperative to confirm Pb loss within the zircon grains in these localities and produce more accurate and precise U-Pb dates.
This study’s U-Pb LA-ICP-MS dataset was examined in two different ways: first, with a 10% discordance filter, and second, with common Pb correction using the 207Pb method, while varying the bandwidth values. This analysis revealed biases when calculating MDAs using statistical approaches reliant on KDE distributions (Figs. 7 & 8). To mitigate these biases, we suggest using lower bandwidth values to generate KDEs with higher resolution, avoiding oversmoothing KDE distribution peaks. Furthermore, elemental and mineralogical analyses conducted on our samples validate paleo-continental reconstructions (Fig. 10). Specifically, they indicate island arc sediment contributions for Borrisnoe Mountain in Ireland and Cwm Graig Ddu in Wales. In contrast, Capel Horeb in Wales shows sediment input from an active continental margin. The findings significantly contribute to the understanding of tectonic settings during the Silurian, and the refined U-Pb MDAs can assist adjusting the tree of life by providing a more precise calibration of molecular clocks and estimating divergence times for various plant lineages.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S0016756824000384
Acknowledgements
Data supporting the conclusions can be obtained from the supplementary material (supplementary tables S1, S2) and will be placed in the Cambridge University Press Supplementary Material data archive. We appreciate funding for this work by the Geological Society of America Graduate Student Specialized Grant from the Charles A. & June R.P. Ross Research Fund, and a Student Research Award from the University of Texas at Austin (UT Austin) Center for Planetary Systems Habitability. U-Pb dates were collected at the JSG UTChron Laboratory at UT Austin. We appreciate analytical assistance by L. Stockli and comments from L. Boucher and J. Clarke. We appreciate comments from two anonymous reviewers. We thank Coillte, Ben Evans from the British Institute for Geological Conservation and Christina Byrne from Natural Resources Wales for permission to responsibly sample the localities for this study.
Competing interests
The author(s) declare none.












 Open access
Open access