



1. Introduction
A mass-action kinetics system exhibits absolute concentration robustness (ACR) if the steady-state value of at least one species is robust to fluctuations in initial concentrations of all species [Reference Shinar and Feinberg27]. Another biologically significant property is the existence of multiple steady states, that is, multistationarity. Significantly, this property has been linked to cellular decision-making and switch-like responses [Reference Kholodenko, Hancock and Kolch21, Reference Tyson, Albert, Goldbeter, Ruoff and Sible33].
As both ACR and multistationarity are important properties, it is perhaps surprising that their relationship was explored only recently, when the present authors with Joshi showed that ACR and multistationarity together – or even ACR by itself – are highly atypical in randomly generated reaction networks [Reference Joshi, Kaihnsa, Nguyen and Shiu18]. This result dovetails with the fact that the two properties are somewhat in opposition, as multiple steady states are not in general position in the presence of ACR.
The results of Joshi et al. are asymptotic in nature (as the number of species goes to infinity), and they pertain to networks that are at-most-bimolecular (which is typical of networks arising in biology) and reversible (which is not) [Reference Joshi, Kaihnsa, Nguyen and Shiu18]. This naturally leads to the following question:
Question 1.1. For multistationarity and ACR to coexist, how many species, reactions, and complexes are needed? Which networks (without the requirement of being reversible) of small to medium size allow such coexistence?
Another motivation for Question 1.1 comes from synthetic biology. In order to design reaction networks with certain dynamical properties, we need to better understand the design principles that allow for such behaviours, as well as the constraints on the size (such as the minimum numbers of species, reaction, and complexes) of such networks. Another possible measure of size is the dimension of a network, which is the difference between the number of species and the number of linearly independent linear conservation laws.
Our work focuses on answering Question 1.1. Broadly speaking, our results fall into two categories: (i) results that give lower bounds on the dimension of a network or its number of species, reactions, or complexes and (ii) results for certain classes of networks (one-dimensional, up to two species, and so on). Our primary focus is on at-most-bimolecular networks, but we also present results on general networks.
In the first category, our results are summarised in the following theorem, which gives some minimum requirements for ACR and nondegenerate multistationarity to coexist. This coexistence is typically on a nonzero-measure subset of the parameter space of reaction rate constants.
Theorem 1.2 (Main result). Let
 $G$
be an at-most-bimolecular reaction network with
$G$
be an at-most-bimolecular reaction network with
 $n$
species such that there exists a vector of positive rate constants
$n$
species such that there exists a vector of positive rate constants
 $\kappa ^*$
such that the mass-action system
$\kappa ^*$
such that the mass-action system
 $(G,\kappa ^*)$
has ACR and is nondegenerately multistationary. Then,
$(G,\kappa ^*)$
has ACR and is nondegenerately multistationary. Then,
 $G$
has:
$G$
has:
-
1. at least three species (that is,
 $n \geq 3$
),
$n \geq 3$
),
-
2. at least three reactant complexes (and hence, at least three reactions) and at least five complexes (reactant and product complexes), and
-
3. dimension at least
$2$
.


If, additionally,
 $G$
is full-dimensional (that is,
$G$
is full-dimensional (that is,
 $G$
has no linear conservation laws), then
$G$
has no linear conservation laws), then
 $G$
has:
$G$
has:
-
4. at least
$n+2$
reactant complexes (and hence, at least
$n+2$
reactions), and
-
5. dimension at least
$3$
.



For the proof of Theorem 1.2, we refer the reader to Section 3 for part (3) (Lemma 3.20); Section 4 for parts (1), (2), and (5) (Theorem 4.1); and Section 5 for part (4) (Theorem 5.1). Additionally, many of the lower bounds in Theorem 1.2 are tight. Indeed, this is shown for parts (1)–(3) through the following mass-conserving network:
 $\{A+B \to 2C \to 2B,\, C\to A \}$
(Example 4.10). As for part (4), this bound is proven for networks that need not be at-most-bimolecular, and its tightness is shown in that context (Proposition 5.3).
$\{A+B \to 2C \to 2B,\, C\to A \}$
(Example 4.10). As for part (4), this bound is proven for networks that need not be at-most-bimolecular, and its tightness is shown in that context (Proposition 5.3).
While Theorem 1.2 concerns nondegenerate multistationarity, we also investigate the capacity for ACR together with degenerate multistationarity, specifically, in networks with four-reactant complexes (Proposition 4.15). Finally, we prove two additional results in the spirit of Theorem 1.2. The first states that
 $3$
is the minimum number of pairs of reversible reactions needed (in reversible networks) for multistationarity, even without ACR (Theorem 3.21). The second concerns networks that are not full-dimensional and states the minimum number of reactant complexes needed for the coexistence of ACR and nondegenerate multistationarity is
$3$
is the minimum number of pairs of reversible reactions needed (in reversible networks) for multistationarity, even without ACR (Theorem 3.21). The second concerns networks that are not full-dimensional and states the minimum number of reactant complexes needed for the coexistence of ACR and nondegenerate multistationarity is
 $n-k+1$
, where
$n-k+1$
, where
 $1 \leq k \leq n-2$
is the number of linearly independent conservation laws (Theorem 5.5).
$1 \leq k \leq n-2$
is the number of linearly independent conservation laws (Theorem 5.5).
As for our second category of results, we start with one-dimensional networks, a class of networks for which ACR [Reference Mendoza, Talabis, Jose and Fontanil23, Reference Meshkat, Shiu and Torres24], multistationarity [Reference Joshi and Shiu20, Reference Shiu and de Wolff28], and even multistability [Reference Tang and Zhang31] are well-studied. Such networks do not allow for the coexistence of ACR and nondegenerate multistationarity (Proposition 3.10). Moreover, one-dimensional bimolecular networks can only be multistationary if they are degenerately so (Lemma 3.20). Moreover, we explicitly characterise all such degenerate networks (Lemma 3.19). Here our proofs make use of recent results of Lin, Tang, and Zhang [Reference Lin, Tang and Zhang22, Reference Tang and Zhang31].
Another class of at-most-bimolecular networks we analyse are those with exactly two species (Section 4.1). For such networks that are reversible, we characterise the property of unconditional ACR, which means that ACR occurs for all possible values of rate constants (Theorem 4.4). As for networks that need not be reversible, we show that ACR and multistationarity can coexist, but only in a degenerate way. Moreover, up to relabelling species, only two such networks allow such coexistence for a nonzero-measure subset of the space of reaction rate constants (Theorem 4.8).
Our works fit into a growing body of literature that explores the minimal conditions needed for various dynamical behaviours, including the two properties that are the focus of the current work: multistationarity [Reference Joshi and Shiu20, Reference Lin, Tang and Zhang22, Reference Shiu and de Wolff28] and ACR [Reference Mendoza, Talabis, Jose and Fontanil23, Reference Meshkat, Shiu and Torres24]. There are additional such studies on multistability [Reference Tang and Wang29] and Hopf bifurcations [Reference Banaji and Boros5, Reference Banaji, Boros and Hofbauer6, Reference Tang and Xu30, Reference Tang and Zhang31, Reference Wilhelm and Heinrich34] (which generate periodic orbits). For instance, in analogy to Theorem 1.2 above, the presence of Hopf bifurcations requires an at-most-bimolecular network to have at least three species, four reactions, and dimension
 $3$
[Reference Banaji and Boros5, Reference Wilhelm and Heinrich34].
$3$
[Reference Banaji and Boros5, Reference Wilhelm and Heinrich34].
This article is organised as follows: Section 2 introduces reaction networks, multistationarity, and ACR. Section 3 contains several results on steady states and their nondegeneracy. We use these results in Sections 4 and 5 to prove our main results. We conclude with a discussion in Section 6.
2. Background
This section recalls the basic setup and definitions involving reaction networks (Section 2.1), the dynamical systems they generate (Section 2.2), absolute concentration robustness (Section 2.3), and a concept pertaining to networks with only one species: “arrow diagrams” (Section 2.4).
2.1. Reaction networks
A reaction network
 $G$
is a (loopless) directed graph in which the vertices are non-negative-integer linear combinations of species
$G$
is a (loopless) directed graph in which the vertices are non-negative-integer linear combinations of species
 $X_1, X_2, \ldots, X_n$
. Each vertex is a complex, and we denote the complex at vertex
$X_1, X_2, \ldots, X_n$
. Each vertex is a complex, and we denote the complex at vertex
 $i$
by
$i$
by
 $y_i=\sum _{j=1}^{n}y_{ij}X_j$
(where
$y_i=\sum _{j=1}^{n}y_{ij}X_j$
(where
 $y_{ij}\in \mathbb Z_{\geq 0}$
) or
$y_{ij}\in \mathbb Z_{\geq 0}$
) or
 $y_i=(y_{i1}, y_{i2}, \dots, y_{in} )$
. Throughout, we assume that each species
$y_i=(y_{i1}, y_{i2}, \dots, y_{in} )$
. Throughout, we assume that each species
 $X_i$
, where
$X_i$
, where
 $i=1,2,\dots,n$
, appears in at least one complex.
$i=1,2,\dots,n$
, appears in at least one complex.
Edges of a network
 $G$
are reactions (in chemistry, these are ‘reaction steps’), and it is standard to represent a reaction
$G$
are reactions (in chemistry, these are ‘reaction steps’), and it is standard to represent a reaction
 $(y_i, y_j)$
by
$(y_i, y_j)$
by
 $y_i \to y_j$
. In such a reaction,
$y_i \to y_j$
. In such a reaction,
 $y_i$
is the reactant complex, and
$y_i$
is the reactant complex, and
 $y_j$
is the product complex; and
$y_j$
is the product complex; and
 $y_i \neq y_j$
, as
$y_i \neq y_j$
, as
 $G$
is loopless. A species
$G$
is loopless. A species
 $X_k$
is a catalyst-only species in reaction
$X_k$
is a catalyst-only species in reaction
 $y_i \to y_j$
if
$y_i \to y_j$
if
 $y_{ik} = y_{jk}$
. In examples, it is often convenient to write species as
$y_{ik} = y_{jk}$
. In examples, it is often convenient to write species as
 $A,B,C,\dots$
(rather than
$A,B,C,\dots$
(rather than
 $X_1,X_2,X_3,\dots$
) and also to view a network as a set of reactions, where the sets of species and complexes are implied.
$X_1,X_2,X_3,\dots$
) and also to view a network as a set of reactions, where the sets of species and complexes are implied.
Example 2.1.
The reaction network
 $\{ 0 \leftarrow A \to 2A,\, B \leftarrow A+B\}$
has two species, five complexes, and three reactions. The species
$\{ 0 \leftarrow A \to 2A,\, B \leftarrow A+B\}$
has two species, five complexes, and three reactions. The species
 $B$
is a catalyst-only species in the reaction
$B$
is a catalyst-only species in the reaction
 $B \leftarrow A+B$
.
$B \leftarrow A+B$
.
A reaction network is reversible if every edge of the graph is bidirected. A reaction network is weakly reversible if every connected component of the graph is strongly connected. Every reversible network is weakly reversible.
Example 2.2.
The following network is reversible:
 $\{ A+B{\leftrightarrows } 2A, \,2B{\leftrightarrows } A, 0 \leftrightarrows B \}$
. The network
$\{ A+B{\leftrightarrows } 2A, \,2B{\leftrightarrows } A, 0 \leftrightarrows B \}$
. The network
 $\{ A+B \to 2A \to 2B \to A+B \}$
is weakly reversible, but not reversible.
$\{ A+B \to 2A \to 2B \to A+B \}$
is weakly reversible, but not reversible.
One focus of our work is on at-most-bimolecular reaction networks (or, for short, bimolecular), which means that every complex
 $y_i$
satisfies
$y_i$
satisfies
 $y_{i1}+y_{i2} + \dots + y_{in} \leq 2$
. Equivalently, each complex has the form
$y_{i1}+y_{i2} + \dots + y_{in} \leq 2$
. Equivalently, each complex has the form
 $0$
,
$0$
,
 $X_i$
,
$X_i$
,
 $X_i+X_j$
, or
$X_i+X_j$
, or
 $2X_i$
(where
$2X_i$
(where
 $X_i$
and
$X_i$
and
 $X_j$
are species). The networks in Examples 2.1–2.2 are bimolecular.
$X_j$
are species). The networks in Examples 2.1–2.2 are bimolecular.
2.2. Mass-action systems
Let
 $r$
denote the number of reactions of
$r$
denote the number of reactions of
 $G$
. We write the
$G$
. We write the
 $i$
th reaction as
$i$
th reaction as
 $y_i \to y^{\prime}_i$
and assign to it a positive rate constant
$y_i \to y^{\prime}_i$
and assign to it a positive rate constant
 $\kappa _{i}\in{\mathbb{R}}_{\gt 0}$
. We use
$\kappa _{i}\in{\mathbb{R}}_{\gt 0}$
. We use
 $x=(x_1,x_2, \dots, x_n)$
to denote the vector of species concentrations. The mass-action system arising from a network
$x=(x_1,x_2, \dots, x_n)$
to denote the vector of species concentrations. The mass-action system arising from a network
 $G$
and a vector of positive rate constants
$G$
and a vector of positive rate constants
 $\kappa =(\kappa _1, \kappa _2, \dots, \kappa _r)$
, which we denote by
$\kappa =(\kappa _1, \kappa _2, \dots, \kappa _r)$
, which we denote by
 $(G,\kappa )$
, is the following dynamical system arising from mass-action kinetics:
$(G,\kappa )$
, is the following dynamical system arising from mass-action kinetics:
 \begin{equation} \frac{dx}{dt} = \sum _{i=1}^r \kappa _{i} x^{y_i} (y^{\prime}_i-y_i) \,=\!:\, f_{\kappa }(x), \end{equation}
\begin{equation} \frac{dx}{dt} = \sum _{i=1}^r \kappa _{i} x^{y_i} (y^{\prime}_i-y_i) \,=\!:\, f_{\kappa }(x), \end{equation}
where
 $x^{y_i} \,:\!=\, \prod _{j=1}^n x_j^{y_{ij}}$
. Observe that the right-hand side of the ODEs (1) consists of polynomials
$x^{y_i} \,:\!=\, \prod _{j=1}^n x_j^{y_{ij}}$
. Observe that the right-hand side of the ODEs (1) consists of polynomials
 $f_{\kappa,i}(x)$
, for
$f_{\kappa,i}(x)$
, for
 $i=1,\dots,n$
. For simplicity, we often write
$i=1,\dots,n$
. For simplicity, we often write
 $f_i$
instead of
$f_i$
instead of
 $f_{\kappa,i}$
. Hence,
$f_{\kappa,i}$
. Hence,
 $f_{\kappa }(x)\,:\!=\,(f_1(x), f_2(x), \ldots,f_n(x))$
is a vector-valued polynomial function.
$f_{\kappa }(x)\,:\!=\,(f_1(x), f_2(x), \ldots,f_n(x))$
is a vector-valued polynomial function.
The question of which polynomials
 $f_i$
can appear as right-hand side of mass-action ODEs is answered in the following result [Reference Hars and Tóth15, Theorem 3.2].
$f_i$
can appear as right-hand side of mass-action ODEs is answered in the following result [Reference Hars and Tóth15, Theorem 3.2].
Lemma 2.3.
Let
 $f\,:\,\mathbb{R}^n \rightarrow \mathbb{R}^n$
be a polynomial function, that is, assume that
$f\,:\,\mathbb{R}^n \rightarrow \mathbb{R}^n$
be a polynomial function, that is, assume that
 $f_i \in \mathbb{R}[x_1, x_2 \dots, x_n]$
for
$f_i \in \mathbb{R}[x_1, x_2 \dots, x_n]$
for
 $i=1,2,\dots, n$
. Then,
$i=1,2,\dots, n$
. Then,
 $f$
arises as the right-hand side of the differential equations (1) (for some choice of network
$f$
arises as the right-hand side of the differential equations (1) (for some choice of network
 $G$
and vector of positive rate constants
$G$
and vector of positive rate constants
 $\kappa$
) if and only if, for all
$\kappa$
) if and only if, for all
 $i=1,2,\dots,n$
, every monomial in
$i=1,2,\dots,n$
, every monomial in
 $f_i$
with negative coefficient is divisible by
$f_i$
with negative coefficient is divisible by
 $x_i$
.
$x_i$
.
Next, observe that the mass-action ODEs (1) are in the linear subspace of
 $\mathbb{R}^n$
spanned by all reaction vectors
$\mathbb{R}^n$
spanned by all reaction vectors
 $y^{\prime}_i - y_i$
(for
$y^{\prime}_i - y_i$
(for
 $i=1,2,\dots, r$
). We call this the stoichiometric subspace and denote it by
$i=1,2,\dots, r$
). We call this the stoichiometric subspace and denote it by
 $S$
. The dimension of a network is the dimension of its stoichiometric subspace. (This dimension is sometimes called the “rank” [Reference Banaji4, Reference Tang and Wang29].) In particular, if
$S$
. The dimension of a network is the dimension of its stoichiometric subspace. (This dimension is sometimes called the “rank” [Reference Banaji4, Reference Tang and Wang29].) In particular, if
 $\text{dim}(S)=n$
(that is,
$\text{dim}(S)=n$
(that is,
 $S= \mathbb{R}^n$
), we say that
$S= \mathbb{R}^n$
), we say that
 $G$
is full-dimensional.
$G$
is full-dimensional.
A trajectory
 $x(t)$
of (1) with initial condition
$x(t)$
of (1) with initial condition
 $x(0) = x^0 \in \mathbb R_{\gt 0}^n$
remains, for all positive time, in the following stoichiometric compatibility class of
$x(0) = x^0 \in \mathbb R_{\gt 0}^n$
remains, for all positive time, in the following stoichiometric compatibility class of
 $G$
[Reference Feinberg11]:
$G$
[Reference Feinberg11]:
 \begin{align} P_{x(0)} \,:\!=\, (x(0)+S)\cap{\mathbb{R}}_{\geq 0}^n. \end{align}
\begin{align} P_{x(0)} \,:\!=\, (x(0)+S)\cap{\mathbb{R}}_{\geq 0}^n. \end{align}
For full-dimensional networks, there is a unique stoichiometric compatibility class:
 $P={\mathbb{R}}_{\geq 0}^n$
. For networks that are not full-dimensional, every nonzero vector
$P={\mathbb{R}}_{\geq 0}^n$
. For networks that are not full-dimensional, every nonzero vector
 $w$
in
$w$
in
 $S^{\perp }$
yields a (linear) conservation law
$S^{\perp }$
yields a (linear) conservation law
 $\langle w,x \rangle = \langle w, x(0) \rangle$
that is satisfied by every
$\langle w,x \rangle = \langle w, x(0) \rangle$
that is satisfied by every
 $x \in P_{x(0)}$
, where
$x \in P_{x(0)}$
, where
 $\langle -,- \rangle$
denotes the usual inner product on
$\langle -,- \rangle$
denotes the usual inner product on
 $\mathbb R^n$
.
$\mathbb R^n$
.
Remark 2.4. A common linear conservation law is mass conservation, where
 $w = (1,1,\dots,1)$
. In fact, several examples in our paper include mass-conserving networks (i.e. networks with mass conservation). However, in general it is not required that the components of
$w = (1,1,\dots,1)$
. In fact, several examples in our paper include mass-conserving networks (i.e. networks with mass conservation). However, in general it is not required that the components of
 $w$
are non-negative.
$w$
are non-negative.
Example 2.1 (continued). The network
 $\{ 0 \overset{\kappa _1}{\leftarrow } A \overset{\kappa _2}{\to } 2A,\, B \overset{\kappa _3}{\leftarrow } A+B\}$
has a one-dimensional stoichiometric subspace (spanned by
$\{ 0 \overset{\kappa _1}{\leftarrow } A \overset{\kappa _2}{\to } 2A,\, B \overset{\kappa _3}{\leftarrow } A+B\}$
has a one-dimensional stoichiometric subspace (spanned by
 $(1,0)$
) and generates the following mass-action ODEs (1):
$(1,0)$
) and generates the following mass-action ODEs (1):
 \begin{align} &\frac{dx_1}{dt} = -\kappa _1x_1+\kappa _2x_1-\kappa _3x_1 x_2=x_1({-}\kappa _1+\kappa _2-\kappa _3 x_2)\\ &\frac{dx_2}{dt} = 0. \notag \end{align}
\begin{align} &\frac{dx_1}{dt} = -\kappa _1x_1+\kappa _2x_1-\kappa _3x_1 x_2=x_1({-}\kappa _1+\kappa _2-\kappa _3 x_2)\\ &\frac{dx_2}{dt} = 0. \notag \end{align}
Observe that the negative monomials in the first ODE are
 $-\kappa _1x_1$
and
$-\kappa _1x_1$
and
 $-\kappa _3x_1 x_2$
, and each of these is divisible by
$-\kappa _3x_1 x_2$
, and each of these is divisible by
 $x_1$
, which is consistent with Lemma 2.3
. Next, the stoichiometric compatibility classes (2) are rays of the following form (where
$x_1$
, which is consistent with Lemma 2.3
. Next, the stoichiometric compatibility classes (2) are rays of the following form (where
 $T\gt 0$
):
$T\gt 0$
):
 \begin{align} \left\{ (x_1,x_2) \in \mathbb{R}^2_{\geq 0} \mid x_2 = T\right\}. \end{align}
\begin{align} \left\{ (x_1,x_2) \in \mathbb{R}^2_{\geq 0} \mid x_2 = T\right\}. \end{align}
The equation
 $ x_2 = T$
is the unique (up to scaling) conservation law.
$ x_2 = T$
is the unique (up to scaling) conservation law.
A steady state of a mass-action system is a non-negative vector
 $x^*\in \mathbb R_{\geq 0}^n$
at which the right-hand side of the ODEs (1) vanishes:
$x^*\in \mathbb R_{\geq 0}^n$
at which the right-hand side of the ODEs (1) vanishes:
 $f_{\kappa }(x^*)=0$
. Our main interest in this work is in positive steady states
$f_{\kappa }(x^*)=0$
. Our main interest in this work is in positive steady states
 $x^*\in \mathbb R_{\gt 0}^n$
. The set of all positive steady states of a mass-action system can have positive dimension in
$x^*\in \mathbb R_{\gt 0}^n$
. The set of all positive steady states of a mass-action system can have positive dimension in
 ${\mathbb{R}}^n$
, but this set typically intersects each stoichiometric compatibility class in finitely many points [Reference Feliu, Henriksson and Pascual-Escudero14]. Finally, a steady state
${\mathbb{R}}^n$
, but this set typically intersects each stoichiometric compatibility class in finitely many points [Reference Feliu, Henriksson and Pascual-Escudero14]. Finally, a steady state
 $x^*$
is nondegenerate if
$x^*$
is nondegenerate if
 $\text{Im}(df_{\kappa }(x^*)|_S)=S$
, where
$\text{Im}(df_{\kappa }(x^*)|_S)=S$
, where
 $df_{\kappa }(x^*)$
is the Jacobian matrix of
$df_{\kappa }(x^*)$
is the Jacobian matrix of
 $f_\kappa$
evaluated at
$f_\kappa$
evaluated at
 $x^*$
.
$x^*$
.
We consider multiple steady states at two levels: systems and networks. A mass-action system
 $(G,\kappa )$
is multistationary (respectively, nondegenerately multistationary) if there exists a stoichiometric compatibility class having more than one positive steady state (respectively, nondegenerate positive steady state). A reaction network
$(G,\kappa )$
is multistationary (respectively, nondegenerately multistationary) if there exists a stoichiometric compatibility class having more than one positive steady state (respectively, nondegenerate positive steady state). A reaction network
 $G$
is multistationary if there exists a vector of positive rate constants
$G$
is multistationary if there exists a vector of positive rate constants
 $\kappa$
such that
$\kappa$
such that
 $(G,\kappa )$
is multistationary. For a reaction network
$(G,\kappa )$
is multistationary. For a reaction network
 $G$
, we let
$G$
, we let
 $\textrm{cap}_{\text{pos}}(G)$
(respectively,
$\textrm{cap}_{\text{pos}}(G)$
(respectively,
 $\textrm{cap}_{\text{nondeg}}(G)$
) denote the maximum possible number of positive steady states (respectively, nondegenerate positive steady states) in a stoichiometric compatibility class.
$\textrm{cap}_{\text{nondeg}}(G)$
) denote the maximum possible number of positive steady states (respectively, nondegenerate positive steady states) in a stoichiometric compatibility class.
Example 2.1 (continued). We return to the network
 $G=\left\{ 0 \overset{\kappa _1}{\leftarrow } A \overset{\kappa _2}{\to } 2A,\, B \overset{\kappa _3}{\leftarrow } A+B\right\}$
and its ODEs (3). A direct computation reveals that when
$G=\left\{ 0 \overset{\kappa _1}{\leftarrow } A \overset{\kappa _2}{\to } 2A,\, B \overset{\kappa _3}{\leftarrow } A+B\right\}$
and its ODEs (3). A direct computation reveals that when
 $\kappa _1 \geq \kappa _2$
, there is no positive steady state. On the other hand, when
$\kappa _1 \geq \kappa _2$
, there is no positive steady state. On the other hand, when
 $\kappa _2 \gt \kappa _1$
, the steady states form exactly one stoichiometric compatibility class (4) – namely, the one given by
$\kappa _2 \gt \kappa _1$
, the steady states form exactly one stoichiometric compatibility class (4) – namely, the one given by
 $T = (\kappa _2 - \kappa _1)/\kappa _3$
– and all such steady states are degenerate. Hence,
$T = (\kappa _2 - \kappa _1)/\kappa _3$
– and all such steady states are degenerate. Hence,
 $G$
is multistationary but not nondegenerately multistationary.
$G$
is multistationary but not nondegenerately multistationary.
Example 2.2 (continued). The following (full-dimensional) reaction network and indicated rate constants yield a mass-action system with three nondegenerate positive steady states [Reference Joshi, Kaihnsa, Nguyen and Shiu18, Remark 3.6]:
 \begin{align} & \left \{ A+B \mathop{\leftrightarrows }_{1/4}^{1/32} 2A, \quad 2B \mathop{\leftrightarrows }_{1/4}^{1} A, \quad 0 \mathop{\leftrightarrows }_{1}^{1} B \right \}. \end{align}
\begin{align} & \left \{ A+B \mathop{\leftrightarrows }_{1/4}^{1/32} 2A, \quad 2B \mathop{\leftrightarrows }_{1/4}^{1} A, \quad 0 \mathop{\leftrightarrows }_{1}^{1} B \right \}. \end{align}
Therefore, this network is nondegenerately multistationary.
2.3. Deficiency and absolute concentration robustness
The deficiency of a reaction network
 $G$
is
$G$
is
 $\delta = m - \ell - \text{dim}(S)$
, where
$\delta = m - \ell - \text{dim}(S)$
, where
 $m$
is the number of vertices (or complexes),
$m$
is the number of vertices (or complexes),
 $\ell$
is the number of connected components of
$\ell$
is the number of connected components of
 $G$
(also called linkage classes), and
$G$
(also called linkage classes), and
 $S$
is the stoichiometric subspace. The deficiency is always non-negative [Reference Feinberg11], and it plays a central role in many classical results on the dynamical properties of mass-action systems [Reference Anderson and Cotter1–Reference Anderson and Nguyen3, Reference Feinberg10, Reference Horn16, Reference Horn and Jackson17].
$S$
is the stoichiometric subspace. The deficiency is always non-negative [Reference Feinberg11], and it plays a central role in many classical results on the dynamical properties of mass-action systems [Reference Anderson and Cotter1–Reference Anderson and Nguyen3, Reference Feinberg10, Reference Horn16, Reference Horn and Jackson17].
Two such results are stated below. These results, which are due to Feinberg and Horn [Reference Feinberg12, Reference Feinberg13, Reference Horn16], are stated for weakly reversible networks (the setting in which we use these results later).
Lemma 2.5 (Deficiency-zero theorem). Deficiency-zero networks are not multistationary. Moreover, if
 $G$
is a weakly reversible network with deficiency zero, then for every vector of positive rate constants
$G$
is a weakly reversible network with deficiency zero, then for every vector of positive rate constants
 $\kappa$
, the mass-action system
$\kappa$
, the mass-action system
 $(G,\kappa )$
admits a unique positive steady state in every stoichiometric compatibility class.
$(G,\kappa )$
admits a unique positive steady state in every stoichiometric compatibility class.
Lemma 2.6 (Deficiency-one theorem). Consider a weakly reversible network
 $G$
with connected components (linkage classes)
$G$
with connected components (linkage classes)
 $G_1$
,
$G_1$
,
 $G_2$
, …,
$G_2$
, …,
 $G_{\ell }$
. Let
$G_{\ell }$
. Let
 $\delta$
denote the deficiency of
$\delta$
denote the deficiency of
 $G$
, and (for all
$G$
, and (for all
 $i=1,2,\dots, \ell$
) let
$i=1,2,\dots, \ell$
) let
 $\delta _i$
denote the deficiency of
$\delta _i$
denote the deficiency of
 $G_i$
. Assume the following:
$G_i$
. Assume the following:
-
1.
$\delta _i \leq 1$
for all
$i=1,2,\dots,\ell$
, and
-
2.
$\delta _1+\delta _2+ \dots + \delta _{\ell } = \delta$
.



Then,
 $G$
is not multistationary: for every vector of positive rate constants
$G$
is not multistationary: for every vector of positive rate constants
 $\kappa$
, the mass-action system
$\kappa$
, the mass-action system
 $(G,\kappa )$
admits a unique positive steady state in every stoichiometric compatibility class.
$(G,\kappa )$
admits a unique positive steady state in every stoichiometric compatibility class.
Our next topic, ACR, like multistationarity, is analysed at the level of systems and also networks.
Definition 2.7 (ACR). Let
 $X_i$
be a species of a reaction network
$X_i$
be a species of a reaction network
 $G$
with
$G$
with
 $r$
reactions.
$r$
reactions.
-
1. For a fixed vector of positive rate constants
$\kappa \in \mathbb{R}^r_{\gt 0}$
, the mass-action system
$(G,\kappa )$
has absolute concentration robustness (ACR) in
$X_i$
if
$(G,\kappa )$
has a positive steady state and in every positive steady state
$x \in{\mathbb{R}}_{\gt 0}^n$
of the system, the value of
$x_i$
is the same. This value of
$x_i$
is the ACR value of
$X_i$
. -
2. The reaction network
$G$
has unconditional ACR in species
$X_i$
if, for every vector of positive rate constants
$\kappa \in \mathbb{R}^r_{\gt 0}$
, the mass-action system
$(G,\kappa )$
has ACR in
$X_i$
.













Remark 2.8 (Existence of positive steady states). ACR requires the existence of a positive steady state (Definition 2.7(1)). This requirement is sometimes not included in definitions of ACR in the literature. However, this is not an extra requirement for some of the networks we consider, namely weakly reversible networks, for which positive steady states are guaranteed to exist (Boros [Reference Boros7]).
Remark 2.9. The property of unconditional ACR is often too restrictive. Thus, many of our results focus on ACR (or other properties) that hold for some full-dimensional subset of the parameter space of rate constants
 $\mathbb R^r_{\gt 0}$
(where
$\mathbb R^r_{\gt 0}$
(where
 $r$
is the number of reactions of a given network). The Lesbesgue measure of such a subset is nonzero. For simplicity, we use “measure” to mean Lebesgue measure.
$r$
is the number of reactions of a given network). The Lesbesgue measure of such a subset is nonzero. For simplicity, we use “measure” to mean Lebesgue measure.
Example 2.1 (continued). We revisit the network
 $\{ 0 \overset{\kappa _1}{\leftarrow } A \overset{\kappa _2}{\to } 2A,\, B \overset{\kappa _3}{\leftarrow } A+B\}$
. From our earlier analysis, the mass-action system has ACR in
$\{ 0 \overset{\kappa _1}{\leftarrow } A \overset{\kappa _2}{\to } 2A,\, B \overset{\kappa _3}{\leftarrow } A+B\}$
. From our earlier analysis, the mass-action system has ACR in
 $B$
when
$B$
when
 $\kappa _2 \gt \kappa _1$
(which defines a nonzero-measure subset of the rate constant space
$\kappa _2 \gt \kappa _1$
(which defines a nonzero-measure subset of the rate constant space
 $\mathbb R^3_{\gt 0}$
), but lacks ACR when
$\mathbb R^3_{\gt 0}$
), but lacks ACR when
 $\kappa _2 \leq \kappa _1$
(as there are no positive steady states).
$\kappa _2 \leq \kappa _1$
(as there are no positive steady states).
Example 2.10.
Consider the following network
 $G$
, which is bimolecular and full-dimensional:
$G$
, which is bimolecular and full-dimensional:
 \begin{align*} \left.\left\{ 2B \xleftarrow{\kappa _3} B \mathop{\rightleftarrows }_{\kappa _2}^{\kappa _1} A+B \xrightarrow{\kappa _4} A \right\}. \right. \end{align*}
\begin{align*} \left.\left\{ 2B \xleftarrow{\kappa _3} B \mathop{\rightleftarrows }_{\kappa _2}^{\kappa _1} A+B \xrightarrow{\kappa _4} A \right\}. \right. \end{align*}
The mass-action ODEs are as follows:
 \begin{align} \dot{x}_1 & = \kappa _1x_2 -\kappa _2x_1 x_2 = (\kappa _1-\kappa _2x_1)x_2 \notag \\ \dot{x}_2 & = \kappa _3x_2-\kappa _4x_1x_2 = (\kappa _3-\kappa _4x_1)x_2. \end{align}
\begin{align} \dot{x}_1 & = \kappa _1x_2 -\kappa _2x_1 x_2 = (\kappa _1-\kappa _2x_1)x_2 \notag \\ \dot{x}_2 & = \kappa _3x_2-\kappa _4x_1x_2 = (\kappa _3-\kappa _4x_1)x_2. \end{align}
When
 $\tfrac{\kappa _1}{\kappa _2} \neq \tfrac{\kappa _3}{\kappa _4}$
, there are no positive steady states and hence no ACR. Now assume
$\tfrac{\kappa _1}{\kappa _2} \neq \tfrac{\kappa _3}{\kappa _4}$
, there are no positive steady states and hence no ACR. Now assume
 $\tfrac{\kappa _1}{\kappa _2}=\tfrac{\kappa _3}{\kappa _4}$
. In this case, the positive steady states are defined by the line
$\tfrac{\kappa _1}{\kappa _2}=\tfrac{\kappa _3}{\kappa _4}$
. In this case, the positive steady states are defined by the line
 $x_1=\tfrac{\kappa _1}{\kappa _2}$
, and so the system is multistationary and has ACR in species
$x_1=\tfrac{\kappa _1}{\kappa _2}$
, and so the system is multistationary and has ACR in species
 $A$
. However, all the steady states of this system are degenerate.
$A$
. However, all the steady states of this system are degenerate.
In the next example, the steady states are nondegenerate.
Example 2.11.
Consider the following full-dimensional network [
Reference Joshi, Kaihnsa, Nguyen and Shiu18, Example 2.6], which we call
 $G$
:
$G$
:
 \begin{align*} \left \{ 2B\mathop{\rightleftarrows }_{\kappa _3}^{\kappa _4} B \mathop{\rightleftarrows }_{\kappa _2}^{\kappa _1} A + B, \quad 2A \mathop{\rightleftarrows }_{\kappa _6}^{\kappa _5} 3A \right \}. \end{align*}
\begin{align*} \left \{ 2B\mathop{\rightleftarrows }_{\kappa _3}^{\kappa _4} B \mathop{\rightleftarrows }_{\kappa _2}^{\kappa _1} A + B, \quad 2A \mathop{\rightleftarrows }_{\kappa _6}^{\kappa _5} 3A \right \}. \end{align*}
The mass-action ODEs (1) are as follows:
 \begin{align*} &\frac{dx_1}{dt}\, =\, \kappa _1x_2-\kappa _2x_1x_2+\kappa _5x_1^2-\kappa _6x_1^3\\[6pt] &\frac{dx_2}{dt} = \kappa _3x_2 - \kappa _4x_2^2. \end{align*}
\begin{align*} &\frac{dx_1}{dt}\, =\, \kappa _1x_2-\kappa _2x_1x_2+\kappa _5x_1^2-\kappa _6x_1^3\\[6pt] &\frac{dx_2}{dt} = \kappa _3x_2 - \kappa _4x_2^2. \end{align*}
The existence of positive steady states comes from the fact that
 $G$
is reversible (recall Remark 2.8). Indeed, it is straightforward to see from the ODEs that
$G$
is reversible (recall Remark 2.8). Indeed, it is straightforward to see from the ODEs that
 $G$
admits up to three positive steady states and that the steady-state value of
$G$
admits up to three positive steady states and that the steady-state value of
 $x_2$
is
$x_2$
is
 $\kappa _3/\kappa _4$
. It follows that
$\kappa _3/\kappa _4$
. It follows that
 $G$
has unconditional ACR in species
$G$
has unconditional ACR in species
 $B$
with ACR value
$B$
with ACR value
 $\kappa _3/\kappa _4$
.
$\kappa _3/\kappa _4$
.
The following result, which is [Reference Meshkat, Shiu and Torres24, Lemma 5.1], concerns ACR in one-dimensional networks.
Lemma 2.12.
Let
 $G$
be a one-dimensional network with species
$G$
be a one-dimensional network with species
 $X_1, X_2, \dots, X_n$
. If
$X_1, X_2, \dots, X_n$
. If
 $G$
has unconditional ACR in some species
$G$
has unconditional ACR in some species
 $X_{i^*}$
, then the reactant complexes of
$X_{i^*}$
, then the reactant complexes of
 $G$
differ only in species
$G$
differ only in species
 $X_{i^*}$
(more precisely, if
$X_{i^*}$
(more precisely, if
 $y$
and
$y$
and
 $\widetilde y$
are both reactant complexes of
$\widetilde y$
are both reactant complexes of
 $G$
, then
$G$
, then
 $y_i=\widetilde{y}_i$
for all
$y_i=\widetilde{y}_i$
for all
 $i \in \{1,2,\dots,n\} \smallsetminus \{i^* \})$
.
$i \in \{1,2,\dots,n\} \smallsetminus \{i^* \})$
.
2.4. Arrow diagrams
In this subsection, we recall the arrow diagrams associated with one-species networks. These diagrams are useful for stating results about such networks [Reference Joshi and Shiu20, Reference Meshkat, Shiu and Torres24, Reference Obatake, Shiu and Sofia25].
Definition 2.13 (Arrow diagram). Let
 $G$
be a reaction network with only one species
$G$
be a reaction network with only one species
 $Z$
. Let
$Z$
. Let
 $m$
denote the number of (distinct) reactant complexes of
$m$
denote the number of (distinct) reactant complexes of
 $G$
, which we list in increasing order of molecularity:
$G$
, which we list in increasing order of molecularity:
 $a_1 Z, a_2Z, \dots, a_m Z$
(so,
$a_1 Z, a_2Z, \dots, a_m Z$
(so,
 $a_1 \lt a_2 \lt \dots \lt a_m$
). For each index
$a_1 \lt a_2 \lt \dots \lt a_m$
). For each index
 $i$
(corresponding to the reactant complex
$i$
(corresponding to the reactant complex
 $a_iZ$
), we define
$a_iZ$
), we define
 $\rho _i$
as follows:
$\rho _i$
as follows:
 \begin{equation*} \rho _i \,:\!=\, \left \lbrace \begin {array}{l@{\quad}l} \to & \text {if for every reaction $a_i Z \to bZ$ in $G$, the inequality $b \gt a_i$ holds} \\ \leftarrow & \text {if for every reaction $a_i Z \to bZ$ in $G$, the inequality $b \lt a_i$ holds} \\ \leftarrow \!\!\bullet \!\!\rightarrow & \text {otherwise.} \end {array}\right. \end{equation*}
\begin{equation*} \rho _i \,:\!=\, \left \lbrace \begin {array}{l@{\quad}l} \to & \text {if for every reaction $a_i Z \to bZ$ in $G$, the inequality $b \gt a_i$ holds} \\ \leftarrow & \text {if for every reaction $a_i Z \to bZ$ in $G$, the inequality $b \lt a_i$ holds} \\ \leftarrow \!\!\bullet \!\!\rightarrow & \text {otherwise.} \end {array}\right. \end{equation*}
The vector
 $\rho = (\rho _1, \rho _2, \ldots, \rho _m) \in \{\to, \leftarrow, \leftarrow \!\!\bullet \!\!\rightarrow \}^m$
is called the arrow diagram of
$\rho = (\rho _1, \rho _2, \ldots, \rho _m) \in \{\to, \leftarrow, \leftarrow \!\!\bullet \!\!\rightarrow \}^m$
is called the arrow diagram of
 $G$
.
$G$
.
Example 2.14.
-
(1) The network
$\{0 \leftarrow A,\, 2A \to 3A\}$
has two reactant complexes
$\{A, 2A\}$
. The arrow corresponding to the reactant complex
$A$
is
$\leftarrow$
, and the arrow corresponding to the reactant complex
$2A$
is
$\rightarrow$
. Thus, the arrow diagram of the network is
$({\leftarrow}, \to)$
. -
(2) The network
$\{0 \leftarrow A,\, A \to 2A,\, 2A \to 3A, \,2A\to 4A\}$
has two reactant complexes
$\{A, 2A\}$
. The arrow corresponding to the reactant complex
$A$
is
$\leftarrow \!\!\bullet \!\!\rightarrow$
, and the arrow corresponding to the reactant complex
$2A$
is
$\rightarrow$
. Thus, the arrow diagram of the network is
$(\leftarrow \!\!\bullet \!\!\rightarrow, \to)$
.














It is often useful to consider the arrow diagrams of “embedded” one-species networks, as follows.
Definition 2.15.
Let
 $G$
be a reaction network with species
$G$
be a reaction network with species
 $X_1, X_2, \dots, X_n$
. Given a species
$X_1, X_2, \dots, X_n$
. Given a species
 $X_i$
, the embedded one-species network of
$X_i$
, the embedded one-species network of
 $G$
(with respect to
$G$
(with respect to
 $X_i$
) is obtained by deleting some (possibly empty) subset of the reactions, replacing each remaining reaction
$X_i$
) is obtained by deleting some (possibly empty) subset of the reactions, replacing each remaining reaction
 $a_1 X_1 + a_2 X_2 + \dots +a_s X_s \to b_1 X_1 + b_2 X_2 + \dots +b_s X_s$
by the reaction
$a_1 X_1 + a_2 X_2 + \dots +a_s X_s \to b_1 X_1 + b_2 X_2 + \dots +b_s X_s$
by the reaction
 $a_i X_i \to b_i X_i$
, and then deleting any trivial reactions (i.e. reactions of the form
$a_i X_i \to b_i X_i$
, and then deleting any trivial reactions (i.e. reactions of the form
 $a_i X_i \to a_i X_i$
, in which the reactant and product complexes are equal) and keeping only one copy of duplicate reactions.
$a_i X_i \to a_i X_i$
, in which the reactant and product complexes are equal) and keeping only one copy of duplicate reactions.
Example 2.16.
Consider the network
 $G=\{ 0\leftrightarrows B \to A \}$
. The following networks are embedded one-species networks of
$G=\{ 0\leftrightarrows B \to A \}$
. The following networks are embedded one-species networks of
 $G:\{0 \leftrightarrows B \}$
,
$G:\{0 \leftrightarrows B \}$
,
 $\{0 \to B\}$
,
$\{0 \to B\}$
,
 $\{0 \leftarrow B \}$
, and
$\{0 \leftarrow B \}$
, and
 $\{0 \to A\}$
.
$\{0 \to A\}$
.
3. Results on steady states and nondegeneracy
This section contains results on the steady states of mass-action systems. We use these results in later sections to prove our main results. Section 3.1 analyses the steady states of full-dimensional networks (i.e. those without conservation laws), while Section 3.2 pertains to non-full-dimensional networks. Next, Section 3.3 focuses on bimolecular networks and investigates scenarios in which the right-hand side of a mass-action ODE vanishes. Finally, Section 3.4 concerns bimolecular networks that are reversible.
3.1. Full-dimensional networks
Consider a reaction network
 $G$
with
$G$
with
 $n$
species,
$n$
species,
 $r$
reactions, and exactly
$r$
reactions, and exactly
 $j$
reactant complexesFootnote
1
and let
$j$
reactant complexesFootnote
1
and let
 $\kappa ^* \in \mathbb{R}^r_{\gt 0}$
be a vector of positive rate constants. We often rewrite the mass-action ODE system (1) for
$\kappa ^* \in \mathbb{R}^r_{\gt 0}$
be a vector of positive rate constants. We often rewrite the mass-action ODE system (1) for
 $(G,\kappa ^*)$
as follows:
$(G,\kappa ^*)$
as follows:
 \begin{align} \begin{bmatrix} dx_1/dt \\[5pt] dx_2/dt \\[5pt] \vdots \\[5pt] dx_n/dt \\[5pt] \end{bmatrix} = N \begin{bmatrix} m_1 \\[5pt] m_2 \\[5pt] \vdots \\[5pt] m_j \end{bmatrix}, \end{align}
\begin{align} \begin{bmatrix} dx_1/dt \\[5pt] dx_2/dt \\[5pt] \vdots \\[5pt] dx_n/dt \\[5pt] \end{bmatrix} = N \begin{bmatrix} m_1 \\[5pt] m_2 \\[5pt] \vdots \\[5pt] m_j \end{bmatrix}, \end{align}
where
 $N$
is an
$N$
is an
 $(n \times j)$
-matrix (with real entries) and
$(n \times j)$
-matrix (with real entries) and
 $m_1,m_2,\dots,m_j$
are the distinct monic monomials in
$m_1,m_2,\dots,m_j$
are the distinct monic monomials in
 $x_1, x_2, \ldots,x_n$
given by the reactant complexes. A monomial is monic if its coefficient is
$x_1, x_2, \ldots,x_n$
given by the reactant complexes. A monomial is monic if its coefficient is
 $1$
.
$1$
.
Example 2.10 (continued). The full-dimensional network
 $ \left\{ 2B \xleftarrow{\kappa _3} B\mathop{\rightleftarrows }\limits_{\kappa _2}^{\kappa _1} A+B \xrightarrow{\kappa _4} A \right\} $
has two reactant complexes, which yield the monomials
$ \left\{ 2B \xleftarrow{\kappa _3} B\mathop{\rightleftarrows }\limits_{\kappa _2}^{\kappa _1} A+B \xrightarrow{\kappa _4} A \right\} $
has two reactant complexes, which yield the monomials
 $m_1\,:\!=\,x_2$
and
$m_1\,:\!=\,x_2$
and
 $m_2\,:\!=\,x_1x_2$
. Consider
$m_2\,:\!=\,x_1x_2$
. Consider
 $\left(\kappa ^*_1,\kappa ^*_2,\kappa ^*_3,\kappa ^*_4\right)=(1,2,3,6)$
$\left(\kappa ^*_1,\kappa ^*_2,\kappa ^*_3,\kappa ^*_4\right)=(1,2,3,6)$
 $\big($
so,
$\big($
so,
 $\tfrac{\kappa ^*_1}{\kappa ^*_2}=\tfrac{\kappa ^*_3}{\kappa ^*_4}$
holds
$\tfrac{\kappa ^*_1}{\kappa ^*_2}=\tfrac{\kappa ^*_3}{\kappa ^*_4}$
holds
 $\big)$
. Now the matrix
$\big)$
. Now the matrix
 $N$
, as in (7), is as follows:
$N$
, as in (7), is as follows:
 \begin{align*} N \,:\!=\, \begin{bmatrix} 1&-2 \\[5pt] 3&-6 \end{bmatrix}. \end{align*}
\begin{align*} N \,:\!=\, \begin{bmatrix} 1&-2 \\[5pt] 3&-6 \end{bmatrix}. \end{align*}
This matrix N does not have full rank, and we saw earlier that all steady states of this mass-action system are degenerate. In the next result, part (1) asserts that this phenomenon holds in general.
Proposition 3.1 (Nondegenerate steady states and the matrix
 $N$
). Let
$N$
). Let
 $G$
be a full-dimensional reaction network with
$G$
be a full-dimensional reaction network with
 $n$
species, and
$n$
species, and
 $\kappa ^*$
be a vector of positive rate constants. Let
$\kappa ^*$
be a vector of positive rate constants. Let
 $N$
be a matrix defined, as in (7), by the mass-action ODE system of
$N$
be a matrix defined, as in (7), by the mass-action ODE system of
 $(G,\kappa ^*)$
.
$(G,\kappa ^*)$
.
-
1. If
$\operatorname{rank}(N) \leq n-1$
, then every positive steady state of
$(G,\kappa ^*)$
is degenerate.
-
2. If
$\operatorname{rank}(N) = n$
and
$G$
has exactly
$n+1$
reactant complexes, then the positive steady states of
$(G,\kappa ^*)$
are the positive roots of a system of binomial equations (sharing some common monomial
$m_0$
) of the following form:
where
\begin{equation*} m_i - \beta _i m_{n+1} = 0 \quad \mathrm {for}\,i=1,2,\dots, n, \end{equation*}
$\beta _1,\beta _2, \dots, \beta _{n} \in \mathbb{R}$
and
$m_1, \dots, m_{n+1}$
are distinct monic monomials in
$x_1, x_2, \ldots,x_n$
.
-
3. If
$G$
has exactly
$n+1$
reactant complexes and
$(G,\kappa ^*)$
has a nondegenerate, positive steady state, then
$(G,\kappa ^*)$
is not multistationary.















Proof. Assume
 $(G,\kappa ^*)$
is a full-dimensional mass-action system in
$(G,\kappa ^*)$
is a full-dimensional mass-action system in
 $n$
species, and let
$n$
species, and let
 $N$
be as in (7).
$N$
be as in (7).
First, we prove (1). Assume
 $\operatorname{rank}(N) \leq n-1$
, and let
$\operatorname{rank}(N) \leq n-1$
, and let
 $x^*$
be a positive steady state. It follows that the polynomials
$x^*$
be a positive steady state. It follows that the polynomials
 $f_i$
, as in (1), are linearly dependent (over
$f_i$
, as in (1), are linearly dependent (over
 $\mathbb R$
). Hence, the Jacobian matrix – even before evaluating at
$\mathbb R$
). Hence, the Jacobian matrix – even before evaluating at
 $x^*$
– has rank less than
$x^*$
– has rank less than
 $n.$
Thus, the image of the Jacobian matrix, after evaluating at
$n.$
Thus, the image of the Jacobian matrix, after evaluating at
 $x^*$
, has dimension less than
$x^*$
, has dimension less than
 $n$
, i.e.
$n$
, i.e.
 $\text{Im}(df(x^*)|_S)\neq \mathbb{R}^n =S$
. Hence,
$\text{Im}(df(x^*)|_S)\neq \mathbb{R}^n =S$
. Hence,
 $x^*$
is degenerate.
$x^*$
is degenerate.
Next, we prove (2). As in equation (7), we write the mass-action ODEs for
 $(G,\kappa ^*)$
as
$(G,\kappa ^*)$
as
 \begin{align*} \begin{bmatrix} dx_1/dt \\[5pt] dx_2/dt \\[5pt] \vdots \\[5pt] dx_n/dt \\[5pt] \end{bmatrix} = N \begin{bmatrix} m_1 \\[5pt] \vdots \\[5pt] m_n \\[5pt] m_{n+1}\\[5pt] \end{bmatrix}, \end{align*}
\begin{align*} \begin{bmatrix} dx_1/dt \\[5pt] dx_2/dt \\[5pt] \vdots \\[5pt] dx_n/dt \\[5pt] \end{bmatrix} = N \begin{bmatrix} m_1 \\[5pt] \vdots \\[5pt] m_n \\[5pt] m_{n+1}\\[5pt] \end{bmatrix}, \end{align*}
where
 $N$
is
$N$
is
 $n \times (n+1)$
and the
$n \times (n+1)$
and the
 $m_i$
’s are distinct monic monomials in
$m_i$
’s are distinct monic monomials in
 $x_1,x_2, \ldots,x_n$
.
$x_1,x_2, \ldots,x_n$
.
As
 $G$
is full-dimensional and
$G$
is full-dimensional and
 $\operatorname{rank}(N) =n$
, we can relabel the
$\operatorname{rank}(N) =n$
, we can relabel the
 $m_i$
’s, if needed, so that the square sub-matrix of
$m_i$
’s, if needed, so that the square sub-matrix of
 $N$
formed by the first
$N$
formed by the first
 $n$
columns has rank
$n$
columns has rank
 $n$
. Thus, by row-reducing
$n$
. Thus, by row-reducing
 $N$
, we obtain a matrix of the following form (where
$N$
, we obtain a matrix of the following form (where
 $\beta _1, \beta _2, \ldots,\beta _n \in \mathbb{R}$
):
$\beta _1, \beta _2, \ldots,\beta _n \in \mathbb{R}$
):
 \begin{align*} N^{\prime} \,:\!=\, \left [\begin{array}{c@{\quad}c@{\quad}c|c} & & & -\beta _1\\[5pt] &{I_n} & & -\beta _2 \\[5pt] & & & \vdots \\[5pt] & & & -\beta _n\\[5pt] \end{array}\right ]. \end{align*}
\begin{align*} N^{\prime} \,:\!=\, \left [\begin{array}{c@{\quad}c@{\quad}c|c} & & & -\beta _1\\[5pt] &{I_n} & & -\beta _2 \\[5pt] & & & \vdots \\[5pt] & & & -\beta _n\\[5pt] \end{array}\right ]. \end{align*}
We conclude from the above discussion that the positive steady states of
 $(G,\kappa ^*)$
are the positive roots of the following
$(G,\kappa ^*)$
are the positive roots of the following
 $n$
binomial equations (which are in the desired form):
$n$
binomial equations (which are in the desired form):
 \begin{align} m_i - \beta _i m_{n+1} = 0 \quad \quad \textrm{for\,}i=1,2,\ldots, n. \end{align}
\begin{align} m_i - \beta _i m_{n+1} = 0 \quad \quad \textrm{for\,}i=1,2,\ldots, n. \end{align}
Before moving on to part (3), we summarise what we know (so we can use it later). The positive steady states are the roots of the binomials (8), which we rewrite using Laurent monomials (our interest is in positive roots, so there is no issue of dividing by zero):
 \begin{align} x_1^{a_{i1}} x_2^{a_{i2}} \dots x_n^{a_{in}} \,:\!=\, \frac{m_i}{m_{n+1}} = \beta _i \quad \quad \textrm{for\,}i=1,2,\ldots, n. \end{align}
\begin{align} x_1^{a_{i1}} x_2^{a_{i2}} \dots x_n^{a_{in}} \,:\!=\, \frac{m_i}{m_{n+1}} = \beta _i \quad \quad \textrm{for\,}i=1,2,\ldots, n. \end{align}
We apply the natural
 $\log$
to (9) and obtain the following, which involves the
$\log$
to (9) and obtain the following, which involves the
 $n \times n$
matrix
$n \times n$
matrix
 $A\,:\!=\,(a_{ij})$
:
$A\,:\!=\,(a_{ij})$
:
 \begin{align} A \begin{pmatrix} \ln (x_1) \\[5pt] \ln (x_2) \\[5pt] \vdots \\[5pt] \ln (x_n) \end{pmatrix} = \begin{pmatrix} \ln (\beta _1) \\[5pt] \ln (\beta _2) \\[5pt] \vdots \\[5pt] \ln (\beta _n) \end{pmatrix} \,=\!:\, \ln (\beta ). \end{align}
\begin{align} A \begin{pmatrix} \ln (x_1) \\[5pt] \ln (x_2) \\[5pt] \vdots \\[5pt] \ln (x_n) \end{pmatrix} = \begin{pmatrix} \ln (\beta _1) \\[5pt] \ln (\beta _2) \\[5pt] \vdots \\[5pt] \ln (\beta _n) \end{pmatrix} \,=\!:\, \ln (\beta ). \end{align}
Now we prove (3). Assume
 $x^*$
is a nondegenerate, positive steady state. (We must show that no other positive steady states exist.) By part (1), the
$x^*$
is a nondegenerate, positive steady state. (We must show that no other positive steady states exist.) By part (1), the
 $n \times (n+1)$
matrix
$n \times (n+1)$
matrix
 $N$
has rank
$N$
has rank
 $n$
, so the proof of part (2) above applies. Assume for contradiction that
$n$
, so the proof of part (2) above applies. Assume for contradiction that
 $x^{**}$
is a positive steady state, with
$x^{**}$
is a positive steady state, with
 $x^{**} \neq x^*$
. Then, by (10), the linear system
$x^{**} \neq x^*$
. Then, by (10), the linear system
 $Ay=\ln (\beta )$
has more than one solution, and so
$Ay=\ln (\beta )$
has more than one solution, and so
 $\textrm{rank}(A) \leq n-1$
. It follows that the set of positive steady states,
$\textrm{rank}(A) \leq n-1$
. It follows that the set of positive steady states,
 $\left\{\left(e^{y_1}, e^{y_2}, \dots, e^{y_n}\right) \mid Ay=\ln (\beta )\right\}$
, is positive-dimensional and so (by the Inverse Function Theorem and the fact that
$\left\{\left(e^{y_1}, e^{y_2}, \dots, e^{y_n}\right) \mid Ay=\ln (\beta )\right\}$
, is positive-dimensional and so (by the Inverse Function Theorem and the fact that
 $G$
is full-dimensional) all positive steady states of
$G$
is full-dimensional) all positive steady states of
 $(G,\kappa ^*)$
are degenerate. This is a contradiction, as
$(G,\kappa ^*)$
are degenerate. This is a contradiction, as
 $x^*$
is nondegenerate.
$x^*$
is nondegenerate.
Remark 3.2. For algebraically inclined readers, observe that the equations in Proposition 3.1(2) define a toric variety. Additionally, every such variety has at most one irreducible component that intersects the positive orthant [Reference Conradi, Iosif and Kahle8, Proposition 5.2]. This fact can be used to give a more direct proof of Proposition 3.1(3).
Remark 3.3. The end of the proof of Proposition 3.1 concerns nondegenerate positive steady states and their relation to the dimension of the set of positive steady states. More ideas in this direction are explored in the recent work of Feliu, Henriksson, and Pascual-Escudero [Reference Feliu, Henriksson and Pascual-Escudero14].
Corollary 3.4 (When
 $f_i$
is zero). Let
$f_i$
is zero). Let
 $G$
be a full-dimensional reaction network with
$G$
be a full-dimensional reaction network with
 $n$
species, let
$n$
species, let
 $\kappa ^*$
be a vector of positive rate constants, and let
$\kappa ^*$
be a vector of positive rate constants, and let
 $f_1,f_2,\dots, f_n$
denote the right-hand sides of the mass-action ODEs of
$f_1,f_2,\dots, f_n$
denote the right-hand sides of the mass-action ODEs of
 $(G,\kappa ^*)$
. If
$(G,\kappa ^*)$
. If
 $f_i$
is the zero polynomial, for some
$f_i$
is the zero polynomial, for some
 $i\in \{1,\ldots,n\}$
, then every positive steady state of
$i\in \{1,\ldots,n\}$
, then every positive steady state of
 $(G,\kappa ^*)$
is degenerate.
$(G,\kappa ^*)$
is degenerate.
Proof. This result follows directly from Proposition 3.1(1) and the fact that, in this case, the rank of
 $N$
, as in (7), is strictly less than
$N$
, as in (7), is strictly less than
 $n$
.
$n$
.
The next two results pertain to networks with few reactant complexes (at most
 $n$
, where
$n$
, where
 $n$
is the number of species) and many reactant complexes (at least
$n$
is the number of species) and many reactant complexes (at least
 $n$
), respectively.
$n$
), respectively.
Proposition 3.5 (Networks with few reactants). Let
 $G$
be a reaction network with
$G$
be a reaction network with
 $n$
species.
$n$
species.
-
1. If
$G$
has exactly 1 reactant complex, then, for every vector of positive rate constants
$\kappa ^*$
, the mass-action system
$(G, \kappa ^*)$
has no positive steady states.
-
2. If
$G$
has exactly
$j$
reactant complexes, where
$2 \leq j \leq n$
(in particular,
$n \geq 2$
), and
$G$
is full-dimensional, then every positive steady state (of every mass-action system defined by
$G$
) is degenerate.









Proof. Assume
 $G$
has
$G$
has
 $n$
species, which we denote by
$n$
species, which we denote by
 $X_1, X_2, \dots, X_n$
, with exactly
$X_1, X_2, \dots, X_n$
, with exactly
 $j$
reactant complexes, for some
$j$
reactant complexes, for some
 $1\leq j \leq n$
. Let
$1\leq j \leq n$
. Let
 $\kappa ^*$
be a vector of positive rate constants. As in (7), we write the mass-action ODE system arising from
$\kappa ^*$
be a vector of positive rate constants. As in (7), we write the mass-action ODE system arising from
 $(G, \kappa ^*)$
as follows:
$(G, \kappa ^*)$
as follows:
 \begin{align} \begin{bmatrix} dx_1/dt \\[5pt] \vdots \\[5pt] dx_n/dt \\[5pt] \end{bmatrix} = N \begin{bmatrix} m_1 \\[5pt] \vdots \\[5pt] m_j \\[5pt] \end{bmatrix}\, \,=\!:\, \begin{bmatrix} f_1 \\[5pt] \vdots \\[5pt] f_n \\[5pt] \end{bmatrix}, \end{align}
\begin{align} \begin{bmatrix} dx_1/dt \\[5pt] \vdots \\[5pt] dx_n/dt \\[5pt] \end{bmatrix} = N \begin{bmatrix} m_1 \\[5pt] \vdots \\[5pt] m_j \\[5pt] \end{bmatrix}\, \,=\!:\, \begin{bmatrix} f_1 \\[5pt] \vdots \\[5pt] f_n \\[5pt] \end{bmatrix}, \end{align}
where
 $N\,:\!=\,(N_{ij})$
is an
$N\,:\!=\,(N_{ij})$
is an
 $(n \times j)$
-matrix (with entries in
$(n \times j)$
-matrix (with entries in
 $\mathbb R$
) and
$\mathbb R$
) and
 $m_1, \dots, m_j$
are distinct monic monomials in
$m_1, \dots, m_j$
are distinct monic monomials in
 $x_1, \dots, x_n$
(as
$x_1, \dots, x_n$
(as
 $G$
has
$G$
has
 $n$
species and
$n$
species and
 $j$
reactant complexes).
$j$
reactant complexes).
We first prove part (1). In this case, the right-hand sides of the ODEs have the form
 $f_i = c_i \prod _{k=1}^{n}x_k^{a_k}$
, with at least one
$f_i = c_i \prod _{k=1}^{n}x_k^{a_k}$
, with at least one
 $c_i\neq 0$
. It follows that there are no positive steady states.
$c_i\neq 0$
. It follows that there are no positive steady states.
We prove part (2). Assume that
 $G$
is full-dimensional (the stoichiometric subspace is
$G$
is full-dimensional (the stoichiometric subspace is
 $\mathbb{R}^n$
) and that
$\mathbb{R}^n$
) and that
 $2 \leq j \leq n$
. Let
$2 \leq j \leq n$
. Let
 $x^*=(x_1^*,x_2^*,\ldots,x_n^*)$
be a positive steady state. We must show
$x^*=(x_1^*,x_2^*,\ldots,x_n^*)$
be a positive steady state. We must show
 $x^*$
is degenerate.
$x^*$
is degenerate.
We first consider the subcase when the rank of the matrix
 $N$
is at most
$N$
is at most
 $(n-1)$
. By Proposition 3.1(1), every positive steady state is degenerate.
$(n-1)$
. By Proposition 3.1(1), every positive steady state is degenerate.
Now we handle the remaining subcase, when
 $N$
has rank
$N$
has rank
 $n$
(and hence,
$n$
(and hence,
 $N$
is
$N$
is
 $n\times n$
). Now, solving the steady-state equations
$n\times n$
). Now, solving the steady-state equations
 $f_1= \dots = f_n=0$
can be accomplished by multiplying the expression in (11) by
$f_1= \dots = f_n=0$
can be accomplished by multiplying the expression in (11) by
 $N^{-1}$
, which implies that every monomial
$N^{-1}$
, which implies that every monomial
 $m_1,\dots, m_n$
evaluates to zero at steady state. Hence, no positive steady states exist.
$m_1,\dots, m_n$
evaluates to zero at steady state. Hence, no positive steady states exist.
Proposition 3.6 (Networks with many reactants). If
 $G$
is a full-dimensional network with
$G$
is a full-dimensional network with
 $n$
species and exactly
$n$
species and exactly
 $j$
reactant complexes, where
$j$
reactant complexes, where
 $j \geq n$
, then:
$j \geq n$
, then:
-
1. There exists a vector of positive rate constants
$\kappa ^*$
, such that the corresponding matrix
$N$
, as in (7), has rank
$n$
. -
2. If there exists a vector of positive rate constants
$\kappa ^*$
such that the matrix
$N$
does not have rank
$n$
, then there exists a vector of positive rate constants
$\kappa ^{**}$
such that
$(G,\kappa ^{**})$
has no positive steady states.








Proof. Assume
 $G$
is full-dimensional, with
$G$
is full-dimensional, with
 $n$
species,
$n$
species,
 $r$
reactions (denoted by
$r$
reactions (denoted by
 $y_1 \to y^{\prime}_1, \dots y_r \to y^{\prime}_r$
), and exactly
$y_1 \to y^{\prime}_1, \dots y_r \to y^{\prime}_r$
), and exactly
 $j$
reactant complexes, where
$j$
reactant complexes, where
 $j \geq n$
.
$j \geq n$
.
We begin with part (1). Let
 $\kappa =(\kappa _1,\dots, \kappa _r)$
denote the vector of unknown rate constants (each
$\kappa =(\kappa _1,\dots, \kappa _r)$
denote the vector of unknown rate constants (each
 $\kappa _i$
is a variable). Let
$\kappa _i$
is a variable). Let
 $\widetilde N$
be the
$\widetilde N$
be the
 $(n \times j)$
matrix for
$(n \times j)$
matrix for
 $(G,\kappa )$
in the sense of
$(G,\kappa )$
in the sense of
 $N$
in (7). More precisely, the entries of
$N$
in (7). More precisely, the entries of
 $\widetilde N$
are
$\widetilde N$
are
 $\mathbb Z$
-linear combinations of the
$\mathbb Z$
-linear combinations of the
 $\kappa _i$
’s, such that, for every vector of positive rate constants
$\kappa _i$
’s, such that, for every vector of positive rate constants
 $\kappa ^* \in \mathbb R_{\gt 0}^r$
, the evaluation
$\kappa ^* \in \mathbb R_{\gt 0}^r$
, the evaluation
 $\widetilde N|_{\kappa =\kappa ^*}$
is the matrix
$\widetilde N|_{\kappa =\kappa ^*}$
is the matrix
 $N$
as in (7) for
$N$
as in (7) for
 $(G, \kappa ^*)$
.
$(G, \kappa ^*)$
.
As
 $G$
is full-dimensional, there are no
$G$
is full-dimensional, there are no
 $\mathbb{R}$
-linear relations among the
$\mathbb{R}$
-linear relations among the
 $n$
rows of
$n$
rows of
 $\widetilde N$
. Hence, the size-
$\widetilde N$
. Hence, the size-
 $n$
minors of
$n$
minors of
 $\widetilde N$
define a (possibly empty) measure-zero subset
$\widetilde N$
define a (possibly empty) measure-zero subset
 $V \subseteq \mathbb R^r_{\gt 0}$
. Thus,
$V \subseteq \mathbb R^r_{\gt 0}$
. Thus,
 $\mathbb R^r_{\gt 0} \smallsetminus V$
is nonempty, and every
$\mathbb R^r_{\gt 0} \smallsetminus V$
is nonempty, and every
 $\kappa ^* \in \mathbb R^r_{\gt 0} \smallsetminus V$
yields a matrix
$\kappa ^* \in \mathbb R^r_{\gt 0} \smallsetminus V$
yields a matrix
 $N=\widetilde N|_{\kappa =\kappa ^*}$
with rank
$N=\widetilde N|_{\kappa =\kappa ^*}$
with rank
 $n$
. This proves part (1).
$n$
. This proves part (1).
For part (2), suppose that there exists
 $\kappa ^* \in \mathbb R_{\gt 0}^r$
such that the resulting matrix
$\kappa ^* \in \mathbb R_{\gt 0}^r$
such that the resulting matrix
 $N$
has rank strictly less than
$N$
has rank strictly less than
 $n$
. It follows that there is a linear relation:
$n$
. It follows that there is a linear relation:
 \begin{align} c_1f_{\kappa ^*,1}+\dots + c_nf_{\kappa ^*,n} = 0, \end{align}
\begin{align} c_1f_{\kappa ^*,1}+\dots + c_nf_{\kappa ^*,n} = 0, \end{align}
where
 $c_1, \dots, c_n$
are real numbers – not all
$c_1, \dots, c_n$
are real numbers – not all
 $0$
– and the
$0$
– and the
 $f_{\kappa ^*,i}$
denote the right-hand sides of the mass-action ODEs for
$f_{\kappa ^*,i}$
denote the right-hand sides of the mass-action ODEs for
 $(G,\kappa ^*)$
.
$(G,\kappa ^*)$
.
On the other hand, for unknown rate constants
 $\kappa$
, as in the proof above for part (1),
$\kappa$
, as in the proof above for part (1),
 $c_1f_{\kappa,1}+\dots + c_nf_{\kappa,n}$
is not the zero polynomial. Thus, when we rewrite this expression as a sum over
$c_1f_{\kappa,1}+\dots + c_nf_{\kappa,n}$
is not the zero polynomial. Thus, when we rewrite this expression as a sum over
 $r$
reactions
$r$
reactions
 $y_i \to y^{\prime}_i$
as follows:
$y_i \to y^{\prime}_i$
as follows:
 $c_1f_{\kappa,1}+\dots + c_nf_{\kappa,n} = d_1 \kappa _1 x^{y_1} + \dots + d_r \kappa _r x^{y_r}$
, where
$c_1f_{\kappa,1}+\dots + c_nf_{\kappa,n} = d_1 \kappa _1 x^{y_1} + \dots + d_r \kappa _r x^{y_r}$
, where
 $d_i \in \mathbb{Z}$
for all
$d_i \in \mathbb{Z}$
for all
 $i$
, we conclude that
$i$
, we conclude that
 $d_i \neq 0$
for some
$d_i \neq 0$
for some
 $i$
. By relabelling reactions, if needed, we may assume that
$i$
. By relabelling reactions, if needed, we may assume that
 $i=1$
.
$i=1$
.
Now consider the following vector of positive rate constants
 $ \kappa _\epsilon ^* \,:\!=\, \left(\kappa _1^* + \epsilon, \kappa _2^*, \dots, \kappa _r^*\right)$
, for some
$ \kappa _\epsilon ^* \,:\!=\, \left(\kappa _1^* + \epsilon, \kappa _2^*, \dots, \kappa _r^*\right)$
, for some
 $\epsilon \gt 0$
. Assume for contradiction that
$\epsilon \gt 0$
. Assume for contradiction that
 $(G,\kappa _\epsilon ^*)$
has a positive steady state
$(G,\kappa _\epsilon ^*)$
has a positive steady state
 $x^*$
. At steady state,
$x^*$
. At steady state,
 $f_{\kappa ^*_\epsilon,i}$
evaluates to
$f_{\kappa ^*_\epsilon,i}$
evaluates to
 $0$
, for all
$0$
, for all
 $i$
, and this yields the first equality here:
$i$
, and this yields the first equality here:
 \begin{equation*} 0 = \left ( c_1f_{\kappa ^*_\epsilon,1}+\dots + c_nf_{\kappa ^*_\epsilon,n} \right ) |_{x=x^*} = c_1f_{\kappa ^*,1}|_{x=x^*}+\dots + c_nf_{\kappa ^*,n}|_{x=x^*} + \epsilon d_1 x^{y_1}|_{x=x^*} = \epsilon d_1 x^{y_1}|_{x=x^*}, \end{equation*}
\begin{equation*} 0 = \left ( c_1f_{\kappa ^*_\epsilon,1}+\dots + c_nf_{\kappa ^*_\epsilon,n} \right ) |_{x=x^*} = c_1f_{\kappa ^*,1}|_{x=x^*}+\dots + c_nf_{\kappa ^*,n}|_{x=x^*} + \epsilon d_1 x^{y_1}|_{x=x^*} = \epsilon d_1 x^{y_1}|_{x=x^*}, \end{equation*}
and the second and third equalities come from the fact that the mass-action ODEs are linear in the rate constants and from equation (12), respectively. We obtain
 $x^{y_1}|_{x=x^*}=0$
, which contradicts the fact that
$x^{y_1}|_{x=x^*}=0$
, which contradicts the fact that
 $x^*$
is a positive steady state. This concludes the proof.
$x^*$
is a positive steady state. This concludes the proof.
The next proposition returns to a topic from Proposition 3.1, namely, networks with
 $n$
species and
$n$
species and
 $n+1$
reactant complexes.
$n+1$
reactant complexes.
Proposition 3.7 (Networks with
 $n+1$
reactants). Assume
$n+1$
reactants). Assume
 $G$
is a full-dimensional network, with
$G$
is a full-dimensional network, with
 $n$
species and exactly
$n$
species and exactly
 $n+1$
reactant complexes, which we denote as follows:
$n+1$
reactant complexes, which we denote as follows:
 \begin{align*} y_{i1} X_1 + y_{i2} X_2 + \dots y_{in} X_n \quad \quad \textrm{for}\,i=1,2,\dots,n+1. \end{align*}
\begin{align*} y_{i1} X_1 + y_{i2} X_2 + \dots y_{in} X_n \quad \quad \textrm{for}\,i=1,2,\dots,n+1. \end{align*}
Let
 $A$
denote the
$A$
denote the
 $n \times n$
matrix obtained from the
$n \times n$
matrix obtained from the
 $(n+1) \times n$
matrix
$(n+1) \times n$
matrix
 $Y\,:\!=\,(y_{ij})$
by subtracting the last row from every row and then deleting the last row.
$Y\,:\!=\,(y_{ij})$
by subtracting the last row from every row and then deleting the last row.
-
1. If
$\operatorname{rank}(A) = n$
, then
$G$
is not nondegenerately multistationary.
-
2. If
$\operatorname{rank}(A) \leq n-1$
, then there exists a vector of positive rate constants
$\kappa ^*$
such that
$(G,\kappa ^*)$
has no positive steady states.





Proof.
Case 1:
 $\operatorname{rank}(A) = n$
. Fix an arbitrary vector of positive rate constants
$\operatorname{rank}(A) = n$
. Fix an arbitrary vector of positive rate constants
 $\kappa ^*$
. We must show that
$\kappa ^*$
. We must show that
 $(G, \kappa ^*)$
is not nondegenerately multistationary. Let
$(G, \kappa ^*)$
is not nondegenerately multistationary. Let
 $N$
denote the
$N$
denote the
 $n \times (n+1)$
matrix defined by
$n \times (n+1)$
matrix defined by
 $(G, \kappa ^*)$
, as in (7). We consider two subcases.
$(G, \kappa ^*)$
, as in (7). We consider two subcases.
Subcase:
 $\operatorname{rank}(N) \leq n-1$
. In this subcase, Proposition 3.1(1) implies that every positive steady state of
$\operatorname{rank}(N) \leq n-1$
. In this subcase, Proposition 3.1(1) implies that every positive steady state of
 $(G, \kappa ^*)$
is degenerate, and so
$(G, \kappa ^*)$
is degenerate, and so
 $(G, \kappa ^*)$
is not nondegenerately multistationary.
$(G, \kappa ^*)$
is not nondegenerately multistationary.
Subcase:
 $\operatorname{rank}(N) = n$
. Part (2) of Proposition 3.1 pertains to this setting, so we can follow that proof. In particular, equation (9) – the
$\operatorname{rank}(N) = n$
. Part (2) of Proposition 3.1 pertains to this setting, so we can follow that proof. In particular, equation (9) – the
 $(n \times n)$
matrix
$(n \times n)$
matrix
 $A$
there exactly matches the matrix
$A$
there exactly matches the matrix
 $A$
here – implies that the positive steady states are defined by a linear system of the form
$A$
here – implies that the positive steady states are defined by a linear system of the form
 $Ay=\ln (\beta )$
, where
$Ay=\ln (\beta )$
, where
 $y=(\!\ln (x_1), \dots, \ln (x_n))^{\top }$
. Hence, as
$y=(\!\ln (x_1), \dots, \ln (x_n))^{\top }$
. Hence, as
 $\operatorname{rank}(A) = n$
, we have at most one positive steady state and so
$\operatorname{rank}(A) = n$
, we have at most one positive steady state and so
 $(G, \kappa ^*)$
is not multistationary.
$(G, \kappa ^*)$
is not multistationary.
Case 2:
 $\operatorname{rank}(A) \leq n-1$
. We must show that there exists a choice of rate constants so that the resulting system has no positive steady states.
$\operatorname{rank}(A) \leq n-1$
. We must show that there exists a choice of rate constants so that the resulting system has no positive steady states.
Proposition 3.6(1) implies that there exists
 $\kappa ^*$
such that the following holds:
$\kappa ^*$
such that the following holds:
 \begin{align} \text{the matrix $N$ defined by $(G,\kappa ^*)$ has (full) rank $n$.} \end{align}
\begin{align} \text{the matrix $N$ defined by $(G,\kappa ^*)$ has (full) rank $n$.} \end{align}
Fix such a choice of
 $\kappa ^*$
. If
$\kappa ^*$
. If
 $(G, \kappa ^*)$
has no positive steady states, then we are done. Therefore, for the rest of the proof, we assume that
$(G, \kappa ^*)$
has no positive steady states, then we are done. Therefore, for the rest of the proof, we assume that
 $(G, \kappa ^*)$
admits a positive steady state.
$(G, \kappa ^*)$
admits a positive steady state.
In what follows, we need to consider additional vectors of positive rate constants (besides
 $\kappa ^*$
) and their corresponding matrices
$\kappa ^*$
) and their corresponding matrices
 $N$
, as in (7). Therefore, as in the proof of Proposition 3.6(1), let
$N$
, as in (7). Therefore, as in the proof of Proposition 3.6(1), let
 $\kappa =(\kappa _1,\dots, \kappa _r)$
(where
$\kappa =(\kappa _1,\dots, \kappa _r)$
(where
 $r$
is the number of reactions) denote the vector of unknown rate constants, and let
$r$
is the number of reactions) denote the vector of unknown rate constants, and let
 $\widetilde N$
be the
$\widetilde N$
be the
 $n \times (n+1)$
matrix for
$n \times (n+1)$
matrix for
 $(G,\kappa )$
in the sense of
$(G,\kappa )$
in the sense of
 $N$
in (7), so that for every vector of positive rate constants
$N$
in (7), so that for every vector of positive rate constants
 $\kappa ^* \in \mathbb R_{\gt 0}^r$
, the evaluation
$\kappa ^* \in \mathbb R_{\gt 0}^r$
, the evaluation
 $\widetilde N|_{\kappa =\kappa ^*}$
is the matrix
$\widetilde N|_{\kappa =\kappa ^*}$
is the matrix
 $N$
as in (7).
$N$
as in (7).
We now follow the ideas in the proof of Proposition 3.1, part (2), with the difference being that we now consider unknown rate constants
 $\kappa$
. The mass-action ODEs for
$\kappa$
. The mass-action ODEs for
 $(G,\kappa )$
are given by:
$(G,\kappa )$
are given by:
 \begin{align*} \begin{bmatrix} dx_1/dt \\[5pt] \vdots \\[5pt] dx_n/dt \\[5pt] \end{bmatrix} = \widetilde N \begin{bmatrix} m_1 \\[5pt] \vdots \\[5pt] m_{n+1}\\[5pt] \end{bmatrix}, \end{align*}
\begin{align*} \begin{bmatrix} dx_1/dt \\[5pt] \vdots \\[5pt] dx_n/dt \\[5pt] \end{bmatrix} = \widetilde N \begin{bmatrix} m_1 \\[5pt] \vdots \\[5pt] m_{n+1}\\[5pt] \end{bmatrix}, \end{align*}
where
 $m_1,\dots, m_{n+1}$
are distinct monic monomials in
$m_1,\dots, m_{n+1}$
are distinct monic monomials in
 $x_1,x_2, \ldots,x_n$
.
$x_1,x_2, \ldots,x_n$
.
Our next aim is to row-reduce
 $\widetilde N$
(over the field
$\widetilde N$
(over the field
 $\mathbb{Q}(\kappa _1,\dots, \kappa _r)$
). Accordingly, for
$\mathbb{Q}(\kappa _1,\dots, \kappa _r)$
). Accordingly, for
 $1\leq k \leq n+1$
, let
$1\leq k \leq n+1$
, let
 $[B_k]$
denote the determinant of the matrix obtained from
$[B_k]$
denote the determinant of the matrix obtained from
 $\widetilde N$
by removing the
$\widetilde N$
by removing the
 $k$
th column. By construction, each
$k$
th column. By construction, each
 $[B_k]$
is in
$[B_k]$
is in
 $\mathbb{Z}[\kappa _1,\dots, \kappa _r]$
.
$\mathbb{Z}[\kappa _1,\dots, \kappa _r]$
.
We claim that, for all
 $1 \leq k \leq n+1$
, the polynomial
$1 \leq k \leq n+1$
, the polynomial
 $[B_k]$
is nonzero. By symmetry among the monomials
$[B_k]$
is nonzero. By symmetry among the monomials
 $m_i$
, it suffices to show that
$m_i$
, it suffices to show that
 $[B_{n+1}]$
is nonzero. To show this, assume for contradiction that
$[B_{n+1}]$
is nonzero. To show this, assume for contradiction that
 $[B_{n+1}]=0$
. Then,
$[B_{n+1}]=0$
. Then,
 $\widetilde N$
can be row-reduced to a matrix in which the last row has the form
$\widetilde N$
can be row-reduced to a matrix in which the last row has the form
 $(0,0,\dots, 0, \omega )$
, where
$(0,0,\dots, 0, \omega )$
, where
 $0 \neq \omega \in \mathbb{Q}(\kappa _1,\dots, \kappa _r)$
. Now consider the evaluation at
$0 \neq \omega \in \mathbb{Q}(\kappa _1,\dots, \kappa _r)$
. Now consider the evaluation at
 $\kappa =\kappa ^*$
. By (13), the matrix
$\kappa =\kappa ^*$
. By (13), the matrix
 $N = \widetilde N|_{\kappa =\kappa ^*}$
has (full) rank
$N = \widetilde N|_{\kappa =\kappa ^*}$
has (full) rank
 $n$
, so
$n$
, so
 $\omega |_{\kappa =\kappa ^*}$
is nonzero. However, this implies that positive steady states of
$\omega |_{\kappa =\kappa ^*}$
is nonzero. However, this implies that positive steady states of
 $(G,\kappa ^*)$
satisfy
$(G,\kappa ^*)$
satisfy
 $\omega |_{\kappa =\kappa ^*} m_{n+1} = 0$
, much like in (8). Thus,
$\omega |_{\kappa =\kappa ^*} m_{n+1} = 0$
, much like in (8). Thus,
 $(G,\kappa ^*)$
has no positive steady states, which is a contradiction, and hence our claim holds.
$(G,\kappa ^*)$
has no positive steady states, which is a contradiction, and hence our claim holds.
Next, as
 $[B_{n+1}]$
is nonzero, we can apply a version of Cramer’s rule to row-reduce
$[B_{n+1}]$
is nonzero, we can apply a version of Cramer’s rule to row-reduce
 $\widetilde N$
to the following matrix (where
$\widetilde N$
to the following matrix (where
 $I_n$
denotes the size-
$I_n$
denotes the size-
 $n$
identity matrix):
$n$
identity matrix):
 \begin{align*}{\widetilde N}^{\prime} = \left [ \begin{array}{c@{\quad}c@{\quad}c|c} & & & ({-}1)^{n-1} \tfrac{[B_1]}{[B_{n+1}]} \\[9pt] & I_n & & ({-}1)^{n-2} \tfrac{[B_2]}{[B_{n+1}]} \\[5pt] & & & \vdots \\[5pt] & & & ({-}1)^{0} \tfrac{[B_n]}{[B_{n+1}]}\\[5pt] \end{array} \right ]. \end{align*}
\begin{align*}{\widetilde N}^{\prime} = \left [ \begin{array}{c@{\quad}c@{\quad}c|c} & & & ({-}1)^{n-1} \tfrac{[B_1]}{[B_{n+1}]} \\[9pt] & I_n & & ({-}1)^{n-2} \tfrac{[B_2]}{[B_{n+1}]} \\[5pt] & & & \vdots \\[5pt] & & & ({-}1)^{0} \tfrac{[B_n]}{[B_{n+1}]}\\[5pt] \end{array} \right ]. \end{align*}
Thus, as in (8), the positive steady states are the positive roots of the equations
 $m_i - \beta _i m_{n+1}=0$
(for
$m_i - \beta _i m_{n+1}=0$
(for
 $i=1,2,\dots,n$
), where:
$i=1,2,\dots,n$
), where:
 \begin{align*} \beta _i \,:\!=\, ({-}1)^{n-i+1} \frac{[B_i]}{[B_{n+1}]} \quad \textrm{for}\, i=1,2,\dots,n. \end{align*}
\begin{align*} \beta _i \,:\!=\, ({-}1)^{n-i+1} \frac{[B_i]}{[B_{n+1}]} \quad \textrm{for}\, i=1,2,\dots,n. \end{align*}
Thus,
 $\beta _i|_{\kappa = \kappa ^*} \gt 0$
(for all
$\beta _i|_{\kappa = \kappa ^*} \gt 0$
(for all
 $i=1,2,\dots,n$
), since
$i=1,2,\dots,n$
), since
 $(G,\kappa ^*)$
admits a positive steady state. We conclude from this fact, plus the claim proven earlier (namely, that
$(G,\kappa ^*)$
admits a positive steady state. We conclude from this fact, plus the claim proven earlier (namely, that
 $[B_\ell ] \neq 0$
for all
$[B_\ell ] \neq 0$
for all
 $\ell$
), that the following is an open subset of
$\ell$
), that the following is an open subset of
 $\mathbb{R}^r_{\gt 0}$
that contains
$\mathbb{R}^r_{\gt 0}$
that contains
 $\kappa ^*$
:
$\kappa ^*$
:
 \begin{align*} \Sigma \,:\!=\, \left \{ \bar \kappa \in \mathbb{R}^r_{\gt 0} \,:\, \beta _1|_{\kappa = \bar \kappa } \gt 0,\,\dots,\, \beta _n|_{\kappa = \bar \kappa } \gt 0,\, [B_1]|_{\kappa =\bar \kappa } \neq 0, \dots, [B_{n+1}]|_{\kappa = \bar \kappa } \neq 0 \right \}. \end{align*}
\begin{align*} \Sigma \,:\!=\, \left \{ \bar \kappa \in \mathbb{R}^r_{\gt 0} \,:\, \beta _1|_{\kappa = \bar \kappa } \gt 0,\,\dots,\, \beta _n|_{\kappa = \bar \kappa } \gt 0,\, [B_1]|_{\kappa =\bar \kappa } \neq 0, \dots, [B_{n+1}]|_{\kappa = \bar \kappa } \neq 0 \right \}. \end{align*}
For the rest of the proof, we restrict our attention to rate constants, like
 $\kappa ^*$
, that are in
$\kappa ^*$
, that are in
 $\Sigma$
. For such rate constants, like in (8)–(10), the positive steady states are the roots of the following equation
$\Sigma$
. For such rate constants, like in (8)–(10), the positive steady states are the roots of the following equation
 \begin{align} A \begin{pmatrix} \ln (x_1) \\[5pt] \ln (x_2) \\[5pt] \vdots \\[5pt] \ln (x_n) \end{pmatrix} = \begin{pmatrix} \ln (\beta _1) \\[5pt] \ln (\beta _2) \\[5pt] \vdots \\[5pt] \ln (\beta _n) \end{pmatrix} \,=\!:\, \ln (\beta ). \end{align}
\begin{align} A \begin{pmatrix} \ln (x_1) \\[5pt] \ln (x_2) \\[5pt] \vdots \\[5pt] \ln (x_n) \end{pmatrix} = \begin{pmatrix} \ln (\beta _1) \\[5pt] \ln (\beta _2) \\[5pt] \vdots \\[5pt] \ln (\beta _n) \end{pmatrix} \,=\!:\, \ln (\beta ). \end{align}
Next, as
 $\operatorname{rank}(A) \leq n-1$
, there exists a nonzero vector
$\operatorname{rank}(A) \leq n-1$
, there exists a nonzero vector
 $\gamma \in \mathbb{R}^n$
in the orthogonal complement of the column space of
$\gamma \in \mathbb{R}^n$
in the orthogonal complement of the column space of
 $A$
. By relabelling the
$A$
. By relabelling the
 $m_i$
’s (which permutes the columns of
$m_i$
’s (which permutes the columns of
 $\widetilde N$
), if needed, we may assume that
$\widetilde N$
), if needed, we may assume that
 $\gamma _1 \neq 0$
. By construction of
$\gamma _1 \neq 0$
. By construction of
 $\gamma$
and equation (14), we have
$\gamma$
and equation (14), we have
 $\langle \gamma,\, \ln (\beta ) \rangle = 0$
, which is readily rewritten as follows:
$\langle \gamma,\, \ln (\beta ) \rangle = 0$
, which is readily rewritten as follows:
 \begin{align} \left ( \frac{[B_1]}{[B_{n+1}]}\right )^{\gamma _1} \dots \left (({-}1)^{k+1} \frac{[B_k]}{ [B_{n+1}] }\right )^{\gamma _k} \dots \left (({-}1)^{n+1} \frac{ [B_n]}{ [B_{n+1}] }\right )^{\gamma _n} = 1. \end{align}
\begin{align} \left ( \frac{[B_1]}{[B_{n+1}]}\right )^{\gamma _1} \dots \left (({-}1)^{k+1} \frac{[B_k]}{ [B_{n+1}] }\right )^{\gamma _k} \dots \left (({-}1)^{n+1} \frac{ [B_n]}{ [B_{n+1}] }\right )^{\gamma _n} = 1. \end{align}
For
 $\varepsilon \gt 0$
, let
$\varepsilon \gt 0$
, let
 $\kappa ^*_{\epsilon }$
denote the vector of rate constants obtained from
$\kappa ^*_{\epsilon }$
denote the vector of rate constants obtained from
 $\kappa ^*$
by scaling by
$\kappa ^*$
by scaling by
 $(1+ \varepsilon )$
all rate constants of reactions in which the reactant generates the monomial
$(1+ \varepsilon )$
all rate constants of reactions in which the reactant generates the monomial
 $m_1$
. As
$m_1$
. As
 $\Sigma$
is an open set,
$\Sigma$
is an open set,
 $\kappa ^*_{\epsilon }\in \Sigma$
for
$\kappa ^*_{\epsilon }\in \Sigma$
for
 $\varepsilon$
sufficiently small. Also, by construction, the matrix
$\varepsilon$
sufficiently small. Also, by construction, the matrix
 $\widetilde N |_{\kappa = \kappa ^*_{\varepsilon }}$
is obtained from
$\widetilde N |_{\kappa = \kappa ^*_{\varepsilon }}$
is obtained from
 $\widetilde N |_{\kappa = \kappa ^*}$
by scaling the first column by
$\widetilde N |_{\kappa = \kappa ^*}$
by scaling the first column by
 $(1+ \varepsilon )$
. So, for
$(1+ \varepsilon )$
. So, for
 $2 \leq i \leq n+1$
, we have
$2 \leq i \leq n+1$
, we have
 $[B_i] |_{\kappa = \kappa ^*_{\varepsilon }} = (1+ \epsilon ) [B_i] |_{\kappa = \kappa ^*}$
.
$[B_i] |_{\kappa = \kappa ^*_{\varepsilon }} = (1+ \epsilon ) [B_i] |_{\kappa = \kappa ^*}$
.
Thus, by replacing
 $\kappa ^*$
by
$\kappa ^*$
by
 $\kappa ^*_{\varepsilon }$
, the left-hand side of equation (15) is scaled by
$\kappa ^*_{\varepsilon }$
, the left-hand side of equation (15) is scaled by
 $(1+\varepsilon )^{-\gamma _1}$
, and so there exists
$(1+\varepsilon )^{-\gamma _1}$
, and so there exists
 $\varepsilon \gt 0$
for which equation (15) does not hold (when evaluated at
$\varepsilon \gt 0$
for which equation (15) does not hold (when evaluated at
 $\kappa = \kappa ^*_{\varepsilon }$
). Hence, this vector
$\kappa = \kappa ^*_{\varepsilon }$
). Hence, this vector
 $\kappa ^*_{\varepsilon }$
yields a mass-action system
$\kappa ^*_{\varepsilon }$
yields a mass-action system
 $(G,\kappa ^*_{\varepsilon })$
with no positive steady states, as desired.
$(G,\kappa ^*_{\varepsilon })$
with no positive steady states, as desired.
Proposition 3.6 implies that for networks with at least
 $n$
reactant complexes (where
$n$
reactant complexes (where
 $n$
is the number of species), some choice of rate constants yields a matrix
$n$
is the number of species), some choice of rate constants yields a matrix
 $N$
with (full) rank
$N$
with (full) rank
 $n$
. Our next result shows that when this condition holds (even for networks with fewer reactants), every species appears in at least one reactant complex.
$n$
. Our next result shows that when this condition holds (even for networks with fewer reactants), every species appears in at least one reactant complex.
We introduce the following shorthand (which we use in several of the next results): a complex
 $y_{\ell 1}X_1+ y_{\ell 2}X_2 + \dots + y_{\ell n}X_n$
involves species
$y_{\ell 1}X_1+ y_{\ell 2}X_2 + \dots + y_{\ell n}X_n$
involves species
 $X_i$
if
$X_i$
if
 $y_{\ell i} \neq 0$
. For instance,
$y_{\ell i} \neq 0$
. For instance,
 $X_1+X_2$
involves
$X_1+X_2$
involves
 $X_2$
, but
$X_2$
, but
 $X_1+X_3$
does not.
$X_1+X_3$
does not.
Lemma 3.8 (Reactants involve all species). Let
 $G$
be a full-dimensional reaction network with
$G$
be a full-dimensional reaction network with
 $n$
species, let
$n$
species, let
 $\kappa ^*$
be a vector of positive rate constants, and let
$\kappa ^*$
be a vector of positive rate constants, and let
 $N$
be the matrix for
$N$
be the matrix for
 $(G,\kappa ^*)$
, as in (7). If
$(G,\kappa ^*)$
, as in (7). If
 $\operatorname{rank}(N) = n$
and
$\operatorname{rank}(N) = n$
and
 $(G,\kappa ^*)$
has a positive steady state, then for every species
$(G,\kappa ^*)$
has a positive steady state, then for every species
 $X_i$
, at least one reactant complex of
$X_i$
, at least one reactant complex of
 $G$
involves
$G$
involves
 $X_i$
.
$X_i$
.
Proof. We prove the contrapositive. Assume that there is a species
 $X_i$
such that for every reactant complex
$X_i$
such that for every reactant complex
 $a_1X_1+ a_2 X_2 + \dots +a_nX_n$
we have
$a_1X_1+ a_2 X_2 + \dots +a_nX_n$
we have
 $a_i = 0$
. Then, by Lemma 2.3, the right-hand side of the mass-action ODE for
$a_i = 0$
. Then, by Lemma 2.3, the right-hand side of the mass-action ODE for
 $X_i$
, which we denote by
$X_i$
, which we denote by
 $f_i$
, is a sum of monomials, all of which have positive coefficients. But
$f_i$
, is a sum of monomials, all of which have positive coefficients. But
 $(G,\kappa ^*)$
has a positive steady state, so
$(G,\kappa ^*)$
has a positive steady state, so
 $f_i$
must be
$f_i$
must be
 $0$
. We conclude that the
$0$
. We conclude that the
 $i$
th row (of the
$i$
th row (of the
 $n$
rows) of
$n$
rows) of
 $N$
is the zero row and so
$N$
is the zero row and so
 $\operatorname{rank}(N) \leq n-1$
.
$\operatorname{rank}(N) \leq n-1$
.
3.2. Networks with conservation laws
The following result is similar to several results in the prior subsection, but pertains to networks that are not full-dimensional (including, but not limited to, networks with mass conservation).
Proposition 3.9 (Networks with conservation laws and few reactants). Let
 $G$
be a reaction network with
$G$
be a reaction network with
 $n \geq 3$
species. Assume that
$n \geq 3$
species. Assume that
 $G$
is
$G$
is
 $(n-k)$
-dimensional, where
$(n-k)$
-dimensional, where
 $k \geq 1$
(so,
$k \geq 1$
(so,
 $G$
has
$G$
has
 $k$
conservation laws). If
$k$
conservation laws). If
 $G$
has exactly
$G$
has exactly
 $j$
reactant complexes, for some
$j$
reactant complexes, for some
 $j \in \{2,3,\dots, n-k\}$
, then every positive steady state (of every mass-action system defined by
$j \in \{2,3,\dots, n-k\}$
, then every positive steady state (of every mass-action system defined by
 $G$
) is degenerate.
$G$
) is degenerate.
Proof. We mimic the proofs of Propositions 3.1(1) and 3.5. Let
 $\kappa ^*$
be a vector of positive rate constants. Let
$\kappa ^*$
be a vector of positive rate constants. Let
 $N$
be an
$N$
be an
 $(n \times j)$
matrix defined, as in (7), by
$(n \times j)$
matrix defined, as in (7), by
 $(G,\kappa ^*)$
:
$(G,\kappa ^*)$
:
 \begin{align} \begin{bmatrix} dx_1/dt \\[5pt] \vdots \\[5pt] dx_n/dt \\[5pt] \end{bmatrix} = N \begin{bmatrix} m_1 \\[5pt] \vdots \\[5pt] m_j \\[5pt] \end{bmatrix}\, \,=\!:\, \begin{bmatrix} f_1 \\[5pt] \vdots \\[5pt] f_n \\[5pt] \end{bmatrix}, \end{align}
\begin{align} \begin{bmatrix} dx_1/dt \\[5pt] \vdots \\[5pt] dx_n/dt \\[5pt] \end{bmatrix} = N \begin{bmatrix} m_1 \\[5pt] \vdots \\[5pt] m_j \\[5pt] \end{bmatrix}\, \,=\!:\, \begin{bmatrix} f_1 \\[5pt] \vdots \\[5pt] f_n \\[5pt] \end{bmatrix}, \end{align}
where
 $m_1, \dots, m_j$
are distinct monic monomials in
$m_1, \dots, m_j$
are distinct monic monomials in
 $x_1, \dots, x_n$
.
$x_1, \dots, x_n$
.
We consider two cases. First assume that
 $\operatorname{rank}(N) \leq n-k-1$
. Then, the polynomials
$\operatorname{rank}(N) \leq n-k-1$
. Then, the polynomials
 $f_i$
span a subspace of dimension
$f_i$
span a subspace of dimension
 $\leq n-k-1$
and hence the Jacobian matrix – even before evaluating at a positive steady state – has rank
$\leq n-k-1$
and hence the Jacobian matrix – even before evaluating at a positive steady state – has rank
 $\leq n-k-1$
. Every positive steady state is therefore degenerate.
$\leq n-k-1$
. Every positive steady state is therefore degenerate.
Consider the remaining case:
 $\operatorname{rank}(N) = n-k$
(so,
$\operatorname{rank}(N) = n-k$
(so,
 $j=n-k$
). In this case, multiplication by
$j=n-k$
). In this case, multiplication by
 $N$
defines an injective map
$N$
defines an injective map
 $\mathbb{R}^{n-k} \to \mathbb{R}^n$
. Hence, by (16), the steady-state equations
$\mathbb{R}^{n-k} \to \mathbb{R}^n$
. Hence, by (16), the steady-state equations
 $f_1= \dots = f_{n}=0$
imply the monomial equations
$f_1= \dots = f_{n}=0$
imply the monomial equations
 $m_1=\dots = m_{j}=0$
. Thus, there are no positive steady states.
$m_1=\dots = m_{j}=0$
. Thus, there are no positive steady states.
The next result concerns networks with
 $n-1$
conservation laws, that is, one-dimensional networks.
$n-1$
conservation laws, that is, one-dimensional networks.
Proposition 3.10 (One-dimensional networks). Let
 $G$
be a one-dimensional reaction network, and let
$G$
be a one-dimensional reaction network, and let
 $\kappa ^*$
be a vector of positive rate constants. If
$\kappa ^*$
be a vector of positive rate constants. If
 $(G,\kappa ^*)$
has ACR, then
$(G,\kappa ^*)$
has ACR, then
 $(G,\kappa ^*)$
is not nondegenerately multistationary.
$(G,\kappa ^*)$
is not nondegenerately multistationary.
Proof. Assume that
 $G$
is one-dimensional, with
$G$
is one-dimensional, with
 $n$
species. Thus,
$n$
species. Thus,
 $G$
has
$G$
has
 $n-1$
linearly independent conservation laws. Let
$n-1$
linearly independent conservation laws. Let
 $\kappa ^*$
be a vector of positive rate constants for which there is ACR. We may assume that the ACR species is
$\kappa ^*$
be a vector of positive rate constants for which there is ACR. We may assume that the ACR species is
 $X_1$
(by relabelling species, if needed). Let
$X_1$
(by relabelling species, if needed). Let
 $f_1,\dots, f_n$
denote the right-hand sides of the mass-action ODEs arising from
$f_1,\dots, f_n$
denote the right-hand sides of the mass-action ODEs arising from
 $(G,\kappa ^*)$
.
$(G,\kappa ^*)$
.
Let
 $x^*=(x_1^*, \dots, x_n^*)$
denote an arbitrary positive steady state of
$x^*=(x_1^*, \dots, x_n^*)$
denote an arbitrary positive steady state of
 $(G,\kappa ^*)$
. (The ACR value is
$(G,\kappa ^*)$
. (The ACR value is
 $x^*_1$
.) Let
$x^*_1$
.) Let
 $P_{x^*}$
denote the (one-dimensional) stoichiometric compatibility class that contains
$P_{x^*}$
denote the (one-dimensional) stoichiometric compatibility class that contains
 $x^*$
. It suffices to show that (1)
$x^*$
. It suffices to show that (1)
 $x^*$
is the unique positive steady state in
$x^*$
is the unique positive steady state in
 $P_{x^*}$
or (2)
$P_{x^*}$
or (2)
 $x^*$
is degenerate.
$x^*$
is degenerate.
We consider two cases.
Case (a):
 $X_1$
is not a catalyst-only species (in some reaction of
$X_1$
is not a catalyst-only species (in some reaction of
 $G$
). This implies that
$G$
). This implies that
 $f_2,\dots, f_n$
are all scalar multiples of
$f_2,\dots, f_n$
are all scalar multiples of
 $f_1$
, and that the compatibility class
$f_1$
, and that the compatibility class
 $P_{x^*}$
is defined by
$P_{x^*}$
is defined by
 $n-1$
conservation laws of the form
$n-1$
conservation laws of the form
 $x_j=a_j x_1+b_j$
, where
$x_j=a_j x_1+b_j$
, where
 $a_j,b_j \in \mathbb{R}$
, for
$a_j,b_j \in \mathbb{R}$
, for
 $j\in \{2,3,\dots, n\}$
. By substituting these
$j\in \{2,3,\dots, n\}$
. By substituting these
 $n-1$
relations into
$n-1$
relations into
 $f_1$
, we obtain a univariate polynomial in
$f_1$
, we obtain a univariate polynomial in
 $x_1$
, which we denote by
$x_1$
, which we denote by
 $h$
. If
$h$
. If
 $h$
has multiple positive roots, then there is no ACR, which is a contradiction. If, on the other hand,
$h$
has multiple positive roots, then there is no ACR, which is a contradiction. If, on the other hand,
 $h$
does not have multiple positive roots, then
$h$
does not have multiple positive roots, then
 $P_{x^*}$
does not contain multiple positive steady states (that is,
$P_{x^*}$
does not contain multiple positive steady states (that is,
 $x^*$
is the unique positive steady state in
$x^*$
is the unique positive steady state in
 $P_{x^*}$
).
$P_{x^*}$
).
Case (b):
 $X_1$
is a catalyst-only species in all reactions of
$X_1$
is a catalyst-only species in all reactions of
 $G$
. In this case,
$G$
. In this case,
 $f_1=0$
, and
$f_1=0$
, and
 $x_1=x_1^*$
is a conservation law of
$x_1=x_1^*$
is a conservation law of
 $G$
, and it is one of the defining equations of the compatibility class
$G$
, and it is one of the defining equations of the compatibility class
 $P_{x^*}$
. By relabelling species
$P_{x^*}$
. By relabelling species
 $X_2,\dots, X_n$
, if needed, we may assume that
$X_2,\dots, X_n$
, if needed, we may assume that
 $X_2$
is not a catalyst-only species (as
$X_2$
is not a catalyst-only species (as
 $G$
is one-dimensional). Thus, we can “extend” the conservation law
$G$
is one-dimensional). Thus, we can “extend” the conservation law
 $x_1=T$
to a “basis” of
$x_1=T$
to a “basis” of
 $n-1$
conservation laws that define the compatibility class
$n-1$
conservation laws that define the compatibility class
 $P_{x^*}$
, by appending
$P_{x^*}$
, by appending
 $n-2$
conservation laws of the form
$n-2$
conservation laws of the form
 $x_j=a_jx_2+b_j$
, where
$x_j=a_jx_2+b_j$
, where
 $a_j,b_j \in \mathbb{R}$
, for
$a_j,b_j \in \mathbb{R}$
, for
 $j\in \{3,4,\dots, n\}$
.
$j\in \{3,4,\dots, n\}$
.
Next, we substitute these
 $n-2$
conservation relations into
$n-2$
conservation relations into
 $f_2$
, which yields a polynomial in
$f_2$
, which yields a polynomial in
 $x_1$
and
$x_1$
and
 $x_2$
, which we denote by
$x_2$
, which we denote by
 $g$
. Consider the following set, which is the positive variety of
$g$
. Consider the following set, which is the positive variety of
 $g$
in
$g$
in
 $\mathbb{R}^2_{\gt 0}$
(the values of
$\mathbb{R}^2_{\gt 0}$
(the values of
 $x_3,\dots, x_n$
are free, so we ignore them):
$x_3,\dots, x_n$
are free, so we ignore them):
 \begin{align} \Sigma \,:\!=\, \left\{x\in \mathbb{R}^2_{\gt 0} \mid g(x_1,x_2)=0 \right\}. \end{align}
\begin{align} \Sigma \,:\!=\, \left\{x\in \mathbb{R}^2_{\gt 0} \mid g(x_1,x_2)=0 \right\}. \end{align}
By construction and the fact that there is ACR in
 $X_1$
, the set
$X_1$
, the set
 $\Sigma$
is contained in the hyperplane (line)
$\Sigma$
is contained in the hyperplane (line)
 $x_1=x_1^*$
, and so is either one-dimensional or zero-dimensional. We consider these two subcases separately. First, assume that
$x_1=x_1^*$
, and so is either one-dimensional or zero-dimensional. We consider these two subcases separately. First, assume that
 $\Sigma$
is one-dimensional. In this subcase,
$\Sigma$
is one-dimensional. In this subcase,
 $\Sigma$
equals the subset of the hyperplane
$\Sigma$
equals the subset of the hyperplane
 $x_1=x_1^*$
in the positive quadrant
$x_1=x_1^*$
in the positive quadrant
 $\mathbb{R}^2_{\gt 0}$
, and so the compatibility class
$\mathbb{R}^2_{\gt 0}$
, and so the compatibility class
 $P_{x^*}$
consists entirely of positive steady states. The Inverse Function Theorem now implies that every positive steady state of
$P_{x^*}$
consists entirely of positive steady states. The Inverse Function Theorem now implies that every positive steady state of
 $P_{x^*}$
(in particular,
$P_{x^*}$
(in particular,
 $x^*$
) is degenerate.
$x^*$
) is degenerate.
Consider the remaining subcase, in which
 $\Sigma$
is zero-dimensional (that is,
$\Sigma$
is zero-dimensional (that is,
 $\Sigma$
consists of finitely many points). It follows that
$\Sigma$
consists of finitely many points). It follows that
 $g$
is either non-negative on
$g$
is either non-negative on
 $\mathbb{R}^2_{\gt 0}$
or non-positive on
$\mathbb{R}^2_{\gt 0}$
or non-positive on
 $\mathbb{R}^2_{\gt 0}$
, and so
$\mathbb{R}^2_{\gt 0}$
, and so
 $f_2$
is either non-negative on
$f_2$
is either non-negative on
 $P_{x^*}$
or non-positive on
$P_{x^*}$
or non-positive on
 $P_{x^*}$
. Consequently, as every
$P_{x^*}$
. Consequently, as every
 $f_i$
is a scalar multiple of
$f_i$
is a scalar multiple of
 $f_2$
, the steady state
$f_2$
, the steady state
 $x^*$
is degenerate.
$x^*$
is degenerate.
3.3. Bimolecular networks
We begin this subsection with a result that clarifies how the polynomials arising in mass-action ODEs are constrained when the network is bimolecular.
Lemma 3.11 (Bimolecular networks). Consider a bimolecular mass-action system
 $(G,\kappa ^*)$
with
$(G,\kappa ^*)$
with
 $n$
species. Let
$n$
species. Let
 $f_i$
be the right-hand side of the mass-action ODE for species
$f_i$
be the right-hand side of the mass-action ODE for species
 $X_i$
(for some
$X_i$
(for some
 $1 \leq i \leq n$
). Fix positive values
$1 \leq i \leq n$
). Fix positive values
 $a_j \gt 0$
for all
$a_j \gt 0$
for all
 $j \in \{1,2,\dots, n\} \smallsetminus \{i\}$
. Let
$j \in \{1,2,\dots, n\} \smallsetminus \{i\}$
. Let
 $g_i$
denote the univariate polynomial obtained by evaluating
$g_i$
denote the univariate polynomial obtained by evaluating
 $f_i$
at
$f_i$
at
 $x_j=a_j$
for all
$x_j=a_j$
for all
 $j \in \{1,2,\dots, n\} \smallsetminus \{i\}$
. If the polynomial
$j \in \{1,2,\dots, n\} \smallsetminus \{i\}$
. If the polynomial
 $g_i$
is nonzero, then
$g_i$
is nonzero, then
 $g_i$
has at most one sign change and hence has at most one positive root.
$g_i$
has at most one sign change and hence has at most one positive root.
Proof. Let
 $g_i$
denote the nonzero polynomial obtained by evaluating
$g_i$
denote the nonzero polynomial obtained by evaluating
 $f_i$
at
$f_i$
at
 $x_j=a_j$
for all
$x_j=a_j$
for all
 $j\neq i$
. Several properties of
$j\neq i$
. Several properties of
 $g_i$
arise from the fact that
$g_i$
arise from the fact that
 $G$
is bimolecular: (1)
$G$
is bimolecular: (1)
 $\text{deg}(g_i) \leq 2$
, (2) the coefficient of
$\text{deg}(g_i) \leq 2$
, (2) the coefficient of
 $x_i^2$
is non-positive, and (3) the constant coefficient is non-negative. Thus,
$x_i^2$
is non-positive, and (3) the constant coefficient is non-negative. Thus,
 $g_i$
has at most one sign change, and so Descartes’ rule of signs implies that
$g_i$
has at most one sign change, and so Descartes’ rule of signs implies that
 $g_i$
has at most one positive root.
$g_i$
has at most one positive root.
The next two results pertain to bimolecular mass-action systems in which the right-hand side of some ODE vanishes (Proposition 3.14) or vanishes when evaluated at an ACR value (Proposition 3.15). We motivate these results through the following example.
Example 3.12 (Enlarged Shinar-Feinberg network). A common way to construct a network with an ACR species (e.g.
 $A$
) is through the existence of an
$A$
) is through the existence of an
 $f_i$
that becomes zero when we substitute the ACR value in place of the species. We illustrate this idea through the following network:
$f_i$
that becomes zero when we substitute the ACR value in place of the species. We illustrate this idea through the following network:
 \begin{equation*} G = \left\{A+B \xrightarrow {\kappa _1} 2B,\, B\xrightarrow {\kappa _2} A,\, 0 \xleftarrow {\kappa _3} B+C \xrightarrow {\kappa _4} 2B,\, 0\xrightarrow {\kappa _5}C\right\}. \end{equation*}
\begin{equation*} G = \left\{A+B \xrightarrow {\kappa _1} 2B,\, B\xrightarrow {\kappa _2} A,\, 0 \xleftarrow {\kappa _3} B+C \xrightarrow {\kappa _4} 2B,\, 0\xrightarrow {\kappa _5}C\right\}. \end{equation*}
This network is constructed from a well-studied network first introduced by Shinar and Feinberg [Reference Shinar and Feinberg27] by adding three reactions involving a new species (
 $C$
). We examine the mass-action ODE for
$C$
). We examine the mass-action ODE for
 $B$
:
$B$
:
 \begin{align*} &\frac{dx_2}{dt} = \kappa _1x_1x_2-\kappa _2 x_2 - \kappa _3 x_2x_3+ \kappa _4 x_2x_3 = x_2(\kappa _1 x_1-\kappa _2) + x_2x_3({-}\kappa _3+\kappa _4) = g+h\, =\!:\, f_2, \end{align*}
\begin{align*} &\frac{dx_2}{dt} = \kappa _1x_1x_2-\kappa _2 x_2 - \kappa _3 x_2x_3+ \kappa _4 x_2x_3 = x_2(\kappa _1 x_1-\kappa _2) + x_2x_3({-}\kappa _3+\kappa _4) = g+h\, =\!:\, f_2, \end{align*}
where
 $g\,:\!=\, x_2(\kappa _1 x_1-\kappa _2)$
(which is the right-hand side of the ODE for
$g\,:\!=\, x_2(\kappa _1 x_1-\kappa _2)$
(which is the right-hand side of the ODE for
 $X_2$
in the original Shinar-Feinberg network) and
$X_2$
in the original Shinar-Feinberg network) and
 $h\,:\!=\, x_2x_3({-}\kappa _3+\kappa _4)$
(arising from the additional reactions, involving
$h\,:\!=\, x_2x_3({-}\kappa _3+\kappa _4)$
(arising from the additional reactions, involving
 $X_3$
).
$X_3$
).
Assume
 $\kappa _3=\kappa _4$
. It is easy to check that
$\kappa _3=\kappa _4$
. It is easy to check that
 $(G,\kappa )$
has a positive steady state and also has ACR in species
$(G,\kappa )$
has a positive steady state and also has ACR in species
 $X_1$
with ACR value
$X_1$
with ACR value
 $\alpha = \kappa _2/\kappa _1$
. Also, observe that
$\alpha = \kappa _2/\kappa _1$
. Also, observe that
 $f_2|_{x_1=\alpha }=0$
, as a result of the equalities
$f_2|_{x_1=\alpha }=0$
, as a result of the equalities
 $g|_{x_1=\alpha }=0$
and
$g|_{x_1=\alpha }=0$
and
 $h=0$
(which is due to the equality
$h=0$
(which is due to the equality
 $\kappa _3=\kappa _4$
).
$\kappa _3=\kappa _4$
).
The next two results characterise which reactions can exist in such a situation. More precisely:
-
Proposition 3.14 gives conditions that hold when a mass-action ODE is zero (effectively characterising what reactions can yield
$h=0$
in this case).
-
Proposition 3.15 gives conditions that hold when a mass-action ODE is zero when evaluated at the ACR value (effectively characterising what reactions can yield
$f_2|_{x_1=\alpha }=0$
in this case, involving a decomposition like the one we observed above:
$f_2|_{x_1=\alpha }=g|_{x_1=\alpha } +h$
).



The next result uses the following notation:
Notation 3.13 (Empty complex). We introduce the dummy variable
 $X_0\,:\!=\,0$
, so that (for instance)
$X_0\,:\!=\,0$
, so that (for instance)
 $X_0$
is the empty complex and
$X_0$
is the empty complex and
 $X_i+X_0 \,:\!=\, X_i$
for any species
$X_i+X_0 \,:\!=\, X_i$
for any species
 $X_i$
.
$X_i$
.
The following result clarifies which reactions can exist if some mass-action ODE is zero.
Proposition 3.14 (When
 $f_i$
is zero). Let
$f_i$
is zero). Let
 $G$
be a bimolecular reaction network with
$G$
be a bimolecular reaction network with
 $n$
species
$n$
species
 $X_1, X_2, \dots, X_n$
. Fix
$X_1, X_2, \dots, X_n$
. Fix
 $1 \leq i \leq n$
. Let
$1 \leq i \leq n$
. Let
 $\kappa ^*$
denote a vector of positive rate constants for
$\kappa ^*$
denote a vector of positive rate constants for
 $G$
, and let
$G$
, and let
 $f_i$
denote the right-hand side of the mass-action ODE for species
$f_i$
denote the right-hand side of the mass-action ODE for species
 $X_i$
in the system
$X_i$
in the system
 $(G, \kappa ^*)$
. If
$(G, \kappa ^*)$
. If
 $f_i$
is the zero polynomial, then the set of reactions of
$f_i$
is the zero polynomial, then the set of reactions of
 $G$
in which
$G$
in which
 $X_i$
is a non-catalyst-only species is a (possibly empty) subset of the reactions listed here (where our use of
$X_i$
is a non-catalyst-only species is a (possibly empty) subset of the reactions listed here (where our use of
 $X_0$
follows Notation 3.13):
$X_0$
follows Notation 3.13):
-
1. the reactions of the form
$X_i +X_j \to 2X_i$
(and we denote the rate constant by
$\kappa ^*_{1,j}$
), where
$j\in \{0,1,\ldots n\} \smallsetminus \{i\}$
, -
2. the reactions of the form
$X_i + X_j \to \star$
(with rate constant
$\kappa ^*_{2,j,\ell }$
, where
$\ell$
is an index for such reactions), where
$j\in \{0,1,\ldots n\} \smallsetminus \{i\}$
and
$\star$
is any complex that does not involve
$X_i$
,









and, additionally, the following relationships among the rate constants hold:
 \begin{align} \kappa ^*_{1,j} = \sum _\ell \kappa ^*_{2,j,\ell } \, \quad \quad \mathrm{for\,all\,} j\in \{0,1,\ldots n\} \smallsetminus \{i\}, \end{align}
\begin{align} \kappa ^*_{1,j} = \sum _\ell \kappa ^*_{2,j,\ell } \, \quad \quad \mathrm{for\,all\,} j\in \{0,1,\ldots n\} \smallsetminus \{i\}, \end{align}
where a rate constant is set to 0 if the corresponding reaction is not in
 $G$
.
$G$
.
Proof. Let
 $\kappa ^*$
be a vector of positive rate constants for a bimolecular network
$\kappa ^*$
be a vector of positive rate constants for a bimolecular network
 $G$
with
$G$
with
 $n$
species, and let
$n$
species, and let
 $f_i$
be the right-hand side of
$f_i$
be the right-hand side of
 $(G,\kappa ^*)$
for the species
$(G,\kappa ^*)$
for the species
 $X_i$
. Let
$X_i$
. Let
 $\Sigma$
denote the set of reactions of
$\Sigma$
denote the set of reactions of
 $G$
in which
$G$
in which
 $X_i$
is a non-catalyst-only species. Reactions not in
$X_i$
is a non-catalyst-only species. Reactions not in
 $\Sigma$
do not contribute to
$\Sigma$
do not contribute to
 $f_i$
, so we ignore them for the rest of the proof.
$f_i$
, so we ignore them for the rest of the proof.
We claim that for all reactions in
 $\Sigma$
, the reactant complex is not one of the following five types:
$\Sigma$
, the reactant complex is not one of the following five types:
 $0$
,
$0$
,
 $X_j$
,
$X_j$
,
 $X_j + X_{j^{\prime}}$
,
$X_j + X_{j^{\prime}}$
,
 $2X_j$
,
$2X_j$
,
 $2X_i$
for any
$2X_i$
for any
 $j,j^{\prime} \in \{1,2,\dots, n \} \smallsetminus \{i\}$
. Indeed, any of the first four types of complexes would yield a constant term in
$j,j^{\prime} \in \{1,2,\dots, n \} \smallsetminus \{i\}$
. Indeed, any of the first four types of complexes would yield a constant term in
 $f_i$
(when viewed as a polynomial in
$f_i$
(when viewed as a polynomial in
 $x_i$
) consisting of a sum of monomials with positive coefficients; similarly, the last type (
$x_i$
) consisting of a sum of monomials with positive coefficients; similarly, the last type (
 $2X_i$
) would yield a negative
$2X_i$
) would yield a negative
 $x_i^2$
term (the fact that
$x_i^2$
term (the fact that
 $G$
is bimolecular is used here). However,
$G$
is bimolecular is used here). However,
 $f_i$
is zero, so the claim holds.
$f_i$
is zero, so the claim holds.
It follows that, for every reaction in
 $\Sigma$
, the reactant complex either is
$\Sigma$
, the reactant complex either is
 $X_i$
or has the form
$X_i$
or has the form
 $X_i+X_j$
for some
$X_i+X_j$
for some
 $j \in \{1,2,\dots, n \} \smallsetminus \{i\}$
. It is straightforward to check that all possible such reactions (in which
$j \in \{1,2,\dots, n \} \smallsetminus \{i\}$
. It is straightforward to check that all possible such reactions (in which
 $X_i$
is a non-catalyst-only species) are listed in the proposition. Next, reactions of type (1) in the proposition contribute positively to
$X_i$
is a non-catalyst-only species) are listed in the proposition. Next, reactions of type (1) in the proposition contribute positively to
 $f_i$
, while those of type (2) contribute negatively, as follows:
$f_i$
, while those of type (2) contribute negatively, as follows:
 \begin{align} f_i = \left ( \kappa ^*_{1,0} - \sum _\ell \kappa ^*_{2, 0, \ell } \right ) x_i \,+ \sum _{j\in \{1,2,\dots, n \} \smallsetminus \{i\}} \left ( \kappa ^*_{1,j} - \sum _\ell \kappa ^*_{2,j,\ell } \right ) x_i x_j. \end{align}
\begin{align} f_i = \left ( \kappa ^*_{1,0} - \sum _\ell \kappa ^*_{2, 0, \ell } \right ) x_i \,+ \sum _{j\in \{1,2,\dots, n \} \smallsetminus \{i\}} \left ( \kappa ^*_{1,j} - \sum _\ell \kappa ^*_{2,j,\ell } \right ) x_i x_j. \end{align}
As
 $f_i=0$
, the coefficient of
$f_i=0$
, the coefficient of
 $x_i$
and the coefficient of each
$x_i$
and the coefficient of each
 $x_{ij}$
in (19) must be
$x_{ij}$
in (19) must be
 $0$
, which yields the desired equalities (18).
$0$
, which yields the desired equalities (18).
Proposition 3.14 concerns general (bimolecular) mass-action systems, and now we consider those with ACR. The next result characterises which reactions can exist if some mass-action ODE becomes zero when evaluated at the ACR value.
Proposition 3.15 (When
 $f_i$
is zero at the ACR value). Let
$f_i$
is zero at the ACR value). Let
 $G$
be a bimolecular reaction network with species
$G$
be a bimolecular reaction network with species
 $X_1, X_2, \dots, X_n$
, where
$X_1, X_2, \dots, X_n$
, where
 $n \geq 2$
. Let
$n \geq 2$
. Let
 $\kappa ^*$
denote a vector of positive rate constants. Assume that the mass-action system
$\kappa ^*$
denote a vector of positive rate constants. Assume that the mass-action system
 $(G, \kappa ^*)$
has ACR in species
$(G, \kappa ^*)$
has ACR in species
 $X_1$
with ACR value
$X_1$
with ACR value
 $\alpha \gt 0$
. Fix
$\alpha \gt 0$
. Fix
 $2 \leq i \leq n$
. Let
$2 \leq i \leq n$
. Let
 $f_i$
denote the right-hand side of the mass-action ODE for species
$f_i$
denote the right-hand side of the mass-action ODE for species
 $X_i$
in the system
$X_i$
in the system
 $(G, \kappa ^*)$
. If
$(G, \kappa ^*)$
. If
 $f_i\neq 0$
and
$f_i\neq 0$
and
 $f_i|_{x_1=\alpha }$
is the zero polynomial, then the set of reactions of
$f_i|_{x_1=\alpha }$
is the zero polynomial, then the set of reactions of
 $G$
in which
$G$
in which
 $X_i$
is a non-catalyst-only species is a nonempty subset of the following reactions (the same as the ones in Proposition 3.14):
$X_i$
is a non-catalyst-only species is a nonempty subset of the following reactions (the same as the ones in Proposition 3.14):
-
1. the reactions of the form
$X_i +X_j \to 2X_i$
(and we denote the rate constant by
$\kappa ^*_{1,j}$
), where
$j\in \{0,1,\ldots n\} \smallsetminus \{i\}$
, -
2. the reactions of the form
$X_i + X_j \to \star$
(with rate constant
$\kappa ^*_{2,j,\ell }$
, where
$\ell$
is an index for such reactions), where
$j\in \{0,1,\ldots n\} \smallsetminus \{i\}$
and
$\star$
is any complex that does not involve
$X_i$
.









Additionally, the following relationship between the ACR value
 $\alpha$
and the rate constants holds:
$\alpha$
and the rate constants holds:
 \begin{align} \alpha = \frac{ \left ( \sum _{\ell } \kappa ^*_{2,0, \ell } \right ) - \kappa ^*_{1,0}}{ \kappa ^*_{1,1} - \left ( \sum _\ell \kappa ^*_{2,1,\ell } \right ) }. \end{align}
\begin{align} \alpha = \frac{ \left ( \sum _{\ell } \kappa ^*_{2,0, \ell } \right ) - \kappa ^*_{1,0}}{ \kappa ^*_{1,1} - \left ( \sum _\ell \kappa ^*_{2,1,\ell } \right ) }. \end{align}
In particular, the numerator and denominator of (20) are nonzero. Finally, if
 $n\geq 3$
, then the following relationships among the rate constants hold:
$n\geq 3$
, then the following relationships among the rate constants hold:
 \begin{align} \kappa ^*_{1,j} = \sum _\ell \kappa ^*_{2,j,\ell } \, \quad \quad \textrm{for\,all\,} j\in \{2,3,\ldots n\} \smallsetminus \{i\}. \end{align}
\begin{align} \kappa ^*_{1,j} = \sum _\ell \kappa ^*_{2,j,\ell } \, \quad \quad \textrm{for\,all\,} j\in \{2,3,\ldots n\} \smallsetminus \{i\}. \end{align}
(In equations (20)–(21), a rate constant is set to 0 if the corresponding reaction is not in
 $G$
.)
$G$
.)
Proof. Assume that
 $f_i$
is nonzero, but
$f_i$
is nonzero, but
 $f_i|_{x_1=\alpha }$
is zero. Using properties of polynomial rings over a field, it follows that
$f_i|_{x_1=\alpha }$
is zero. Using properties of polynomial rings over a field, it follows that
 $(x_1- \alpha )$
divides
$(x_1- \alpha )$
divides
 $f_i$
. From the fact that
$f_i$
. From the fact that
 $G$
is bimolecular, we conclude that:
$G$
is bimolecular, we conclude that:
 \begin{align} \notag f_i &= (x_1 - \alpha ) \left ( \beta x_i + \gamma + \sum _{j \in [n] \smallsetminus \{i\}} \delta _j x_j \right ) \\[5pt] &=\, \beta x_1 x_i + \gamma x_1 + \left ( \sum _{j \in [n] \smallsetminus \{i\}} \delta _j x_1 x_j \right ) -\alpha \beta x_i -\alpha \gamma - \left ( \sum _{j \in [n] \smallsetminus \{i\}} \alpha \delta _j x_j \right ), \end{align}
\begin{align} \notag f_i &= (x_1 - \alpha ) \left ( \beta x_i + \gamma + \sum _{j \in [n] \smallsetminus \{i\}} \delta _j x_j \right ) \\[5pt] &=\, \beta x_1 x_i + \gamma x_1 + \left ( \sum _{j \in [n] \smallsetminus \{i\}} \delta _j x_1 x_j \right ) -\alpha \beta x_i -\alpha \gamma - \left ( \sum _{j \in [n] \smallsetminus \{i\}} \alpha \delta _j x_j \right ), \end{align}
for some real numbers
 $\beta, \gamma, \delta _j$
, at least one of which is nonzero.
$\beta, \gamma, \delta _j$
, at least one of which is nonzero.
In the right-hand side of (22), the variable
 $x_i$
does not appear in any of the following monomials (here the hypothesis
$x_i$
does not appear in any of the following monomials (here the hypothesis
 $i\neq 1$
is used):
$i\neq 1$
is used):
 \begin{equation*} \gamma x_1, \quad -\alpha \gamma, \quad \delta _j x_1 x_j, \quad -\alpha \delta _j x_j, \end{equation*}
\begin{equation*} \gamma x_1, \quad -\alpha \gamma, \quad \delta _j x_1 x_j, \quad -\alpha \delta _j x_j, \end{equation*}
for
 $ j \in \{1,2,\dots,n\} \smallsetminus \{i\}$
, so Lemma 2.3 implies that the coefficients of these monomials must be non-negative. Since
$ j \in \{1,2,\dots,n\} \smallsetminus \{i\}$
, so Lemma 2.3 implies that the coefficients of these monomials must be non-negative. Since
 $\alpha \gt 0$
, we conclude that
$\alpha \gt 0$
, we conclude that
 $\gamma =0$
and
$\gamma =0$
and
 $\delta _j=0$
(for all
$\delta _j=0$
(for all
 $j \in \{1,2,\dots,n\} \smallsetminus \{i\}$
).
$j \in \{1,2,\dots,n\} \smallsetminus \{i\}$
).
Thus, using (22), we have
 $f_i= \beta x_1 x_i - \alpha \beta x_i$
, for some
$f_i= \beta x_1 x_i - \alpha \beta x_i$
, for some
 $\beta \in \mathbb{R} \smallsetminus \{0\}$
. Next, we investigate which reactions contribute to the two monomials in
$\beta \in \mathbb{R} \smallsetminus \{0\}$
. Next, we investigate which reactions contribute to the two monomials in
 $f_i$
. For
$f_i$
. For
 $ \beta x_1 x_i$
, the contributing reactions have the form
$ \beta x_1 x_i$
, the contributing reactions have the form
 $X_1+X_i\to 2X_i$
and
$X_1+X_i\to 2X_i$
and
 $X_1 + X_i \to \star$
, where
$X_1 + X_i \to \star$
, where
 $\star$
does not involve
$\star$
does not involve
 $X_i$
. The first reaction contributes positively, while the second type contributes negatively. Let
$X_i$
. The first reaction contributes positively, while the second type contributes negatively. Let
 $\kappa ^*_{1,1}$
be the reaction rate constant for
$\kappa ^*_{1,1}$
be the reaction rate constant for
 $X_1+X_i\to 2X_i$
(as in the statement of the lemma) and
$X_1+X_i\to 2X_i$
(as in the statement of the lemma) and
 $\kappa ^*_{2,1,\ell }$
be the rate constant for reactions of type
$\kappa ^*_{2,1,\ell }$
be the rate constant for reactions of type
 $X_1 + X_i \to \star$
, where
$X_1 + X_i \to \star$
, where
 $\ell$
is an index for all the reactions of this type. We conclude that
$\ell$
is an index for all the reactions of this type. We conclude that
 \begin{align} \kappa ^*_{1,1} - \sum _\ell \kappa ^*_{2,1,\ell } \,= \, \beta. \end{align}
\begin{align} \kappa ^*_{1,1} - \sum _\ell \kappa ^*_{2,1,\ell } \,= \, \beta. \end{align}
Similarly, the monomial
 $-\alpha \beta x_i$
in
$-\alpha \beta x_i$
in
 $f_i$
comes from reactions of the form
$f_i$
comes from reactions of the form
 $X_i \to 2X_i$
, which contributes positively, and
$X_i \to 2X_i$
, which contributes positively, and
 $X_i \to \star$
, which contributes negatively, where
$X_i \to \star$
, which contributes negatively, where
 $\star$
is a complex that does not involve
$\star$
is a complex that does not involve
 $X_i$
. Hence,
$X_i$
. Hence,
 \begin{align} \kappa ^*_{1,0} - \sum _\ell \kappa ^*_{2,0,\ell } = - \alpha \beta. \end{align}
\begin{align} \kappa ^*_{1,0} - \sum _\ell \kappa ^*_{2,0,\ell } = - \alpha \beta. \end{align}
Now the equations (23) and (24) together imply the desired equality (20).
Next, let
 $\Sigma$
denote the set of reactions of
$\Sigma$
denote the set of reactions of
 $G$
in which
$G$
in which
 $X_i$
is a non-catalyst-only species. We showed above that
$X_i$
is a non-catalyst-only species. We showed above that
 $\Sigma$
contains a (nonempty) subset of reactions with rate constants labelled by
$\Sigma$
contains a (nonempty) subset of reactions with rate constants labelled by
 $\kappa ^*_{1,0}$
,
$\kappa ^*_{1,0}$
,
 $\kappa ^*_{1,1}$
,
$\kappa ^*_{1,1}$
,
 $\kappa ^*_{2,0,\ell }$
,
$\kappa ^*_{2,0,\ell }$
,
 $\kappa ^*_{2,1,\ell }$
. Let
$\kappa ^*_{2,1,\ell }$
. Let
 $\Sigma ^{\prime} \subseteq \Sigma$
denote the remaining reactions, and let
$\Sigma ^{\prime} \subseteq \Sigma$
denote the remaining reactions, and let
 $G^{\prime}$
denote the subnetwork defined by the reactions in
$G^{\prime}$
denote the subnetwork defined by the reactions in
 $\Sigma ^{\prime}$
. Let
$\Sigma ^{\prime}$
. Let
 $\kappa ^{\prime}$
be obtained from
$\kappa ^{\prime}$
be obtained from
 $\kappa ^*$
by restricting to coordinates corresponding to reactions in
$\kappa ^*$
by restricting to coordinates corresponding to reactions in
 $\Sigma ^{\prime}$
. By construction, the mass-action ODE of
$\Sigma ^{\prime}$
. By construction, the mass-action ODE of
 $(G^{\prime}, \kappa ^{\prime})$
for species
$(G^{\prime}, \kappa ^{\prime})$
for species
 $X_i$
has the right-hand side equal to
$X_i$
has the right-hand side equal to
 $0$
. So, Proposition 3.14 applies (where reactions arising from
$0$
. So, Proposition 3.14 applies (where reactions arising from
 $j=0,1$
in that proposition are absent from
$j=0,1$
in that proposition are absent from
 $G^{\prime}$
by construction), and yields two conclusions. First,
$G^{\prime}$
by construction), and yields two conclusions. First,
 $\Sigma ^{\prime}$
is a subset of the reactions listed in Proposition 3.15 (specifically, with
$\Sigma ^{\prime}$
is a subset of the reactions listed in Proposition 3.15 (specifically, with
 $j \not = 0,1$
), and so
$j \not = 0,1$
), and so
 $\Sigma$
is a subset of the full list (including
$\Sigma$
is a subset of the full list (including
 $j = 0,1$
). Second, the equations (18) hold (for
$j = 0,1$
). Second, the equations (18) hold (for
 $j \not = 0,1$
), which are the desired equalities (21).
$j \not = 0,1$
), which are the desired equalities (21).
Remark 3.16. The reactions listed in Propositions 3.14 and 3.15 (the lists are the same) are not reversible. Hence, if
 $G$
is a reversible network satisfying the hypotheses of either proposition, then
$G$
is a reversible network satisfying the hypotheses of either proposition, then
 $X_i$
is a catalyst-only species in every reaction of
$X_i$
is a catalyst-only species in every reaction of
 $G$
.
$G$
.
Example 3.12 (continued). We revisit the enlarged Shinar-Feinberg network,
 $ G= \{A+B \xrightarrow{\kappa _1} 2B,\, B\xrightarrow{\kappa _2} A,\, 0 \xleftarrow{\kappa _3} B+C \xrightarrow{\kappa _4} 2B,\, 0\xrightarrow{\kappa _5}C\}.$
Recall that, when
$ G= \{A+B \xrightarrow{\kappa _1} 2B,\, B\xrightarrow{\kappa _2} A,\, 0 \xleftarrow{\kappa _3} B+C \xrightarrow{\kappa _4} 2B,\, 0\xrightarrow{\kappa _5}C\}.$
Recall that, when
 $\kappa _3=\kappa _4$
, the mass-action system
$\kappa _3=\kappa _4$
, the mass-action system
 $(G,\kappa )$
has ACR in
$(G,\kappa )$
has ACR in
 $X_1$
with ACR value
$X_1$
with ACR value
 $\alpha = \kappa _2/\kappa _1$
, and that
$\alpha = \kappa _2/\kappa _1$
, and that
 $f_2|_{x_1=\alpha }=0$
. In the notation of Proposition 3.15
, the rate constants of reactions in which
$f_2|_{x_1=\alpha }=0$
. In the notation of Proposition 3.15
, the rate constants of reactions in which
 $X_2$
is non-catalyst-only are:
$X_2$
is non-catalyst-only are:
 \begin{equation*} \kappa ^*_{1,1}=\kappa _1, \,\quad \kappa ^*_{1,0}=\kappa _2, \,\quad \kappa ^*_{2,3,1}=\kappa _3,\,\quad \kappa ^*_{1,3}=\kappa _4. \end{equation*}
\begin{equation*} \kappa ^*_{1,1}=\kappa _1, \,\quad \kappa ^*_{1,0}=\kappa _2, \,\quad \kappa ^*_{2,3,1}=\kappa _3,\,\quad \kappa ^*_{1,3}=\kappa _4. \end{equation*}
Now the formula in Proposition 3.15 for the ACR value (20) exactly yields the ACR value computed earlier:
 $\alpha = \kappa _2/\kappa _1$
, and the relationship among rate constants (21) recapitulates
$\alpha = \kappa _2/\kappa _1$
, and the relationship among rate constants (21) recapitulates
 $\kappa _3=\kappa _4$
.
$\kappa _3=\kappa _4$
.
Remark 3.17. The formula for the ACR value, in (20), is related to the concept of “robust ratio” introduced by Johnston and Tonello [Reference Tonello and Johnston32].
3.4. Three reversible reactions are necessary for multistationarity
Recall that, in (5), we saw an instance of a (nondegenerately) multistationary, bimolecular network that consists of
 $3$
pairs of reversible reactions. In this subsection, we prove that bimolecular networks with fewer pairs of reversible reactions are non-multistationary (Theorem 3.21). Our proof of Theorem 3.21 requires several supporting lemmas on one-dimensional networks.
$3$
pairs of reversible reactions. In this subsection, we prove that bimolecular networks with fewer pairs of reversible reactions are non-multistationary (Theorem 3.21). Our proof of Theorem 3.21 requires several supporting lemmas on one-dimensional networks.
For the next lemma, recall from Section 2.2 that
 $\textrm{cap}_{\text{pos}}(G)$
(respectively,
$\textrm{cap}_{\text{pos}}(G)$
(respectively,
 $\textrm{cap}_{\text{nondeg}}(G)$
) denotes the maximum possible number of positive (respectively, nondegenerate and positive) steady states of a network
$\textrm{cap}_{\text{nondeg}}(G)$
) denotes the maximum possible number of positive (respectively, nondegenerate and positive) steady states of a network
 $G$
. In Lemma 3.18 below, part (1) was conjectured by Joshi and Shiu [Reference Joshi and Shiu20] and then proved by Lin, Tang, and Zhang [Reference Lin, Tang and Zhang22, Theorem 4.3] (see also [Reference Pantea and Voitiuk26]). Part (2) is due to Tang and Zhang [Reference Tang and Zhang31, Theorem 6.1].
$G$
. In Lemma 3.18 below, part (1) was conjectured by Joshi and Shiu [Reference Joshi and Shiu20] and then proved by Lin, Tang, and Zhang [Reference Lin, Tang and Zhang22, Theorem 4.3] (see also [Reference Pantea and Voitiuk26]). Part (2) is due to Tang and Zhang [Reference Tang and Zhang31, Theorem 6.1].
Lemma 3.18.
Let
 $G$
be a one-dimensional reaction network.
$G$
be a one-dimensional reaction network.
-
1. If
$G$
is multistationary and
$\textrm{cap}_{\text{pos}}(G) \lt \infty$
, then
$G$
has an embedded one-species network with arrow diagram
$({\leftarrow}, \to)$
and another with arrow diagram
$(\to, \leftarrow )$
. -
2. If
$\textrm{cap}_{\text{pos}}(G) \lt \infty$
, then
$\textrm{cap}_{\text{nondeg}}(G) = \textrm{cap}_{\text{pos}}(G)$
.







Joshi and Shiu showed that the network
 $G=\{ 0 \leftarrow A \to 2A\}$
is the only one-species, bimolecular network for which
$G=\{ 0 \leftarrow A \to 2A\}$
is the only one-species, bimolecular network for which
 $\textrm{cap}_{\text{pos}}(G) =\infty$
[Reference Joshi and Shiu19]. The following lemma generalises this result from one-species networks to one-dimensional networks.
$\textrm{cap}_{\text{pos}}(G) =\infty$
[Reference Joshi and Shiu19]. The following lemma generalises this result from one-species networks to one-dimensional networks.
Lemma 3.19 (One-dimensional bimolecular networks with infinitely many steady states). Let
 $G$
be a one-dimensional and bimolecular reaction network with
$G$
be a one-dimensional and bimolecular reaction network with
 $n$
species. The following are equivalent:
$n$
species. The following are equivalent:
-
1.
$\textrm{cap}_{\text{pos}}(G) = \infty$
. -
2. Up to relabelling species,
$G$
is one of the following networks:
-
(a)
$\{2X_1 \leftarrow X_1+X_2 \to 2X_2 \}$
, -
(b)
$\{X_1 \to 2X_1\} \cup \Sigma$
, where
$\Sigma$
consists of at least one reaction from the following set:
\begin{equation*} \{0 \leftarrow X_1 \} \cup \{X_i \leftarrow X_1 + X_i \mid i=2,3,\dots, n\}. \end{equation*}
-






Additionally, for the networks listed above in 2(a) and 2(b), every positive steady state (of every mass-action system arising from the network
 $G$
) is degenerate.
$G$
) is degenerate.
Proof. Let
 $G$
be a one-dimensional, bimolecular reaction network. Up to relabelling species, the one-dimensional stoichiometric subspace is spanned by one of the following seven vectors:
$G$
be a one-dimensional, bimolecular reaction network. Up to relabelling species, the one-dimensional stoichiometric subspace is spanned by one of the following seven vectors:
 \begin{equation} (1,0,0,\dots, 0), (1,-1,0,0,\dots, 0), \end{equation}
\begin{equation} (1,0,0,\dots, 0), (1,-1,0,0,\dots, 0), \end{equation}
 \begin{align} & (1,1,0,0,\dots, 0), (1,-2, 0,0,\dots, 0), (1,1,-1,0,0,\dots, 0), (1,1,-2, 0,0,\dots, 0),\nonumber\\& (1,1,-1,-1, 0,0,\dots, 0). \end{align}
\begin{align} & (1,1,0,0,\dots, 0), (1,-2, 0,0,\dots, 0), (1,1,-1,0,0,\dots, 0), (1,1,-2, 0,0,\dots, 0),\nonumber\\& (1,1,-1,-1, 0,0,\dots, 0). \end{align}
We first consider the case when the stoichiometric subspace is spanned by one of the five vectors listed in (26). The network
 $G$
is then a subnetwork of one of the following networks (where we use
$G$
is then a subnetwork of one of the following networks (where we use
 $A,B,C,D$
in place of
$A,B,C,D$
in place of
 $X_1,X_2,X_3,X_4$
for ease of notation);
$X_1,X_2,X_3,X_4$
for ease of notation);
 \begin{align*} \{0 \leftrightarrows A+B\},\, \{A \leftrightarrows 2B\},\, \{A+B \leftrightarrows C\},\, \{A+B \leftrightarrows 2C\},\, \{A+B \leftrightarrows C+D\}. \end{align*}
\begin{align*} \{0 \leftrightarrows A+B\},\, \{A \leftrightarrows 2B\},\, \{A+B \leftrightarrows C\},\, \{A+B \leftrightarrows 2C\},\, \{A+B \leftrightarrows C+D\}. \end{align*}
A direct calculation shows that the deficiency of
 $G$
is 0, so the deficiency-zero theorem (Lemma 2.5) implies that
$G$
is 0, so the deficiency-zero theorem (Lemma 2.5) implies that
 $G$
is not multistationary. In particular,
$G$
is not multistationary. In particular,
 $\textrm{cap}_{\text{pos}}(G) \lt \infty$
.
$\textrm{cap}_{\text{pos}}(G) \lt \infty$
.
Having shown that the case of (26) is consistent with Lemma 3.19, we now consider the remaining two cases, from (25), separately. First, assume the stoichiometric subspace of
 $G$
is spanned by
$G$
is spanned by
 $(1,0,0,\dots, 0)$
. It follows that the reactions of
$(1,0,0,\dots, 0)$
. It follows that the reactions of
 $G$
form a subset of the following
$G$
form a subset of the following
 $2n+4$
reactions:
$2n+4$
reactions:
 \begin{align*} &0 \mathop{\leftrightarrows }_{k_0}^{m_0} X_1 \mathop{\leftrightarrows }_{m_1}^{\ell _0} 2X_1 \quad \quad 0 \mathop{\leftrightarrows }_{k_1}^{\ell _1} 2X_1 \\[5pt] & X_i \mathop{\leftrightarrows }_{k_i}^{m_i} X_1 + X_i \quad \text{for } i=2,3,\dots, n. \end{align*}
\begin{align*} &0 \mathop{\leftrightarrows }_{k_0}^{m_0} X_1 \mathop{\leftrightarrows }_{m_1}^{\ell _0} 2X_1 \quad \quad 0 \mathop{\leftrightarrows }_{k_1}^{\ell _1} 2X_1 \\[5pt] & X_i \mathop{\leftrightarrows }_{k_i}^{m_i} X_1 + X_i \quad \text{for } i=2,3,\dots, n. \end{align*}
The ODEs for species
 $X_2, X_3, \dots, X_n$
are
$X_2, X_3, \dots, X_n$
are
 $\frac{dx_i}{dt}=0$
so,
$\frac{dx_i}{dt}=0$
so,
 $x_i=T_i$
(with
$x_i=T_i$
(with
 $T_i\gt 0$
) for
$T_i\gt 0$
) for
 $i=2,3,\dots,n$
are the corresponding conservation laws. We substitute these conservation laws into the ODE for
$i=2,3,\dots,n$
are the corresponding conservation laws. We substitute these conservation laws into the ODE for
 $X_1$
:
$X_1$
:
 \begin{align} \frac{d x_1}{dt}|_{x_2=T_2,\dots, x_n=T_n} = & (k_0 + 2 k_1) + (k_2T_2 + \dots + k_nT_n) +m_1 x_1\\[5pt] \notag & - (m_0+ m_2T_2 + \dots + m_nT_n) x_1 - (\ell _0 + 2 \ell _1) x_1^2. \end{align}
\begin{align} \frac{d x_1}{dt}|_{x_2=T_2,\dots, x_n=T_n} = & (k_0 + 2 k_1) + (k_2T_2 + \dots + k_nT_n) +m_1 x_1\\[5pt] \notag & - (m_0+ m_2T_2 + \dots + m_nT_n) x_1 - (\ell _0 + 2 \ell _1) x_1^2. \end{align}
When at least one
 $k_i$
is positive and all other
$k_i$
is positive and all other
 $k_j$
’s are non-negative, the right-hand side of (27), viewed as a polynomial in
$k_j$
’s are non-negative, the right-hand side of (27), viewed as a polynomial in
 $x_1$
, has a nonzero constant term and, hence, is not the zero polynomial. Similarly, if
$x_1$
, has a nonzero constant term and, hence, is not the zero polynomial. Similarly, if
 $\ell _0$
or
$\ell _0$
or
 $\ell _1$
is positive and
$\ell _1$
is positive and
 $\ell _0,\ell _1 \geq 0$
, then the right-hand side of (27) has a nonzero coefficient of
$\ell _0,\ell _1 \geq 0$
, then the right-hand side of (27) has a nonzero coefficient of
 $x_1^2$
and is again a nonzero polynomial. We conclude that if
$x_1^2$
and is again a nonzero polynomial. We conclude that if
 $G$
contains at least one of the reactions labelled by
$G$
contains at least one of the reactions labelled by
 $k_i$
or
$k_i$
or
 $\ell _i$
, then
$\ell _i$
, then
 $\textrm{cap}_{\text{pos}}(G) \lt \infty$
, which is consistent with Lemma 3.19.
$\textrm{cap}_{\text{pos}}(G) \lt \infty$
, which is consistent with Lemma 3.19.
We now consider the case when
 $G$
contains no reactions labelled by
$G$
contains no reactions labelled by
 $k_i$
or
$k_i$
or
 $\ell _i$
, that is, every reaction of
$\ell _i$
, that is, every reaction of
 $G$
is one of the following
$G$
is one of the following
 $n+1$
reactions:
$n+1$
reactions:
 \begin{align*} &0 \stackrel{m_0}{\leftarrow } X_1 \stackrel{m_1}{\to } 2X_1 \\[5pt] & X_i \stackrel{m_i}{\leftarrow } X_1 + X_i \quad \text{for } i=2,3,\dots, n. \end{align*}
\begin{align*} &0 \stackrel{m_0}{\leftarrow } X_1 \stackrel{m_1}{\to } 2X_1 \\[5pt] & X_i \stackrel{m_i}{\leftarrow } X_1 + X_i \quad \text{for } i=2,3,\dots, n. \end{align*}
The right-hand side of the ODE for
 $X_1$
, as in (27), becomes
$X_1$
, as in (27), becomes
 $x_1(m_1 - m_0 - m_2T_2 - \dots - m_nT_n)$
. In order for this polynomial in
$x_1(m_1 - m_0 - m_2T_2 - \dots - m_nT_n)$
. In order for this polynomial in
 $x_1$
to become the zero polynomial for some choice of positive rate constants of
$x_1$
to become the zero polynomial for some choice of positive rate constants of
 $G$
(equivalently,
$G$
(equivalently,
 $\textrm{cap}_{\text{pos}}(G) = \infty$
), we must have
$\textrm{cap}_{\text{pos}}(G) = \infty$
), we must have
 $m_1\gt 0$
and
$m_1\gt 0$
and
 $m_j\gt 0$
for at least one of
$m_j\gt 0$
for at least one of
 $j=0,2,3,\dots,n$
. This gives exactly the reactions listed in Lemma 3.19(2)(b). In this case, given
$j=0,2,3,\dots,n$
. This gives exactly the reactions listed in Lemma 3.19(2)(b). In this case, given
 $m_j\gt 0$
for the reactions appearing in the network, we can always choose
$m_j\gt 0$
for the reactions appearing in the network, we can always choose
 $T_j\gt 0$
, such that the right-hand side of the ODE for
$T_j\gt 0$
, such that the right-hand side of the ODE for
 $X_1$
vanishes (i.e.
$X_1$
vanishes (i.e.
 $\textrm{cap}_{\text{pos}}(G) = \infty$
). Moreover, when this right-hand side vanishes is the only situation in which there are positive steady states, and an easy calculation shows that all such positive steady states are degenerate. This concludes our analysis of networks with stoichiometric subspace spanned by the vector
$\textrm{cap}_{\text{pos}}(G) = \infty$
). Moreover, when this right-hand side vanishes is the only situation in which there are positive steady states, and an easy calculation shows that all such positive steady states are degenerate. This concludes our analysis of networks with stoichiometric subspace spanned by the vector
 $(1,0,0,\dots, 0)$
.
$(1,0,0,\dots, 0)$
.
Our final case is when the stoichiometric subspace is spanned by the vector
 $(1,-1,0,\dots, 0)$
. In this case, the reactions of
$(1,-1,0,\dots, 0)$
. In this case, the reactions of
 $G$
form a subset of the following
$G$
form a subset of the following
 $2n+4$
reactions:
$2n+4$
reactions:
 \begin{align*} & X_1 \mathop{\leftrightarrows }_{k_1}^{\ell _1} X_2 \quad \quad 2 X_1 \mathop{\leftrightarrows }_{k_2}^{\ell _2} 2X_2 \quad \quad 2 X_1 \mathop{\leftrightarrows }_{k_3}^{m_1} X_1 + X_2 \mathop{\leftrightarrows }_{m_2}^{\ell _3} 2 X_2 \\[5pt] & X_1 + X_i\mathop{\leftrightarrows }_{k_{i+1}}^{\ell _{i+1}} X_2 + X_i\quad \text{for } i=3,4,\dots, n. \end{align*}
\begin{align*} & X_1 \mathop{\leftrightarrows }_{k_1}^{\ell _1} X_2 \quad \quad 2 X_1 \mathop{\leftrightarrows }_{k_2}^{\ell _2} 2X_2 \quad \quad 2 X_1 \mathop{\leftrightarrows }_{k_3}^{m_1} X_1 + X_2 \mathop{\leftrightarrows }_{m_2}^{\ell _3} 2 X_2 \\[5pt] & X_1 + X_i\mathop{\leftrightarrows }_{k_{i+1}}^{\ell _{i+1}} X_2 + X_i\quad \text{for } i=3,4,\dots, n. \end{align*}
The conservation laws are
 $x_1+x_2=T_2$
and
$x_1+x_2=T_2$
and
 $x_i=T_i$
for
$x_i=T_i$
for
 $i=3,4,\dots, n$
. The ODE for species
$i=3,4,\dots, n$
. The ODE for species
 $X_1$
is:
$X_1$
is:
 \begin{align} \frac{dx_1}{dt} = & -(2k_2+k_3) x_1^2 -k_1x_1 -(k_4 x_3 + \dots + k_{n+1}x_n) x_1 + (m_1-m_2) x_1 x_2\\[5pt] & \notag \quad + \left(\ell _1 x_2 + 2 \ell _2 x_2^2 + \ell _3 x_2^2\right) + (\ell _4 x_3 + \dots + \ell _{n+1} x_n) x_2. \end{align}
\begin{align} \frac{dx_1}{dt} = & -(2k_2+k_3) x_1^2 -k_1x_1 -(k_4 x_3 + \dots + k_{n+1}x_n) x_1 + (m_1-m_2) x_1 x_2\\[5pt] & \notag \quad + \left(\ell _1 x_2 + 2 \ell _2 x_2^2 + \ell _3 x_2^2\right) + (\ell _4 x_3 + \dots + \ell _{n+1} x_n) x_2. \end{align}
Consider the subcase when at least one of the
 $\ell _i$
is positive and all other
$\ell _i$
is positive and all other
 $\ell _j$
’s are non-negative. After substituting the expressions arising from the conservation laws (namely,
$\ell _j$
’s are non-negative. After substituting the expressions arising from the conservation laws (namely,
 $x_2= T_2-x_1$
and
$x_2= T_2-x_1$
and
 $x_i=T_i$
for
$x_i=T_i$
for
 $i=3,4,\dots, n$
) into the right-hand side of the ODE (28), we obtain a polynomial in
$i=3,4,\dots, n$
) into the right-hand side of the ODE (28), we obtain a polynomial in
 $x_1$
that has a positive constant term (see the second line of the right-hand side of (28)). Hence, if
$x_1$
that has a positive constant term (see the second line of the right-hand side of (28)). Hence, if
 $G$
contains at least one of the reactions labelled by
$G$
contains at least one of the reactions labelled by
 $\ell _i$
, then
$\ell _i$
, then
 $\textrm{cap}_{\text{pos}}(G) \lt \infty$
.
$\textrm{cap}_{\text{pos}}(G) \lt \infty$
.
By symmetry, if
 $G$
has at least one of the reactions labelled by
$G$
has at least one of the reactions labelled by
 $k_i$
, then again
$k_i$
, then again
 $\textrm{cap}_{\text{pos}}(G) \lt \infty$
. Hence, if
$\textrm{cap}_{\text{pos}}(G) \lt \infty$
. Hence, if
 $G$
contains a reaction labelled by
$G$
contains a reaction labelled by
 $\ell _i$
or
$\ell _i$
or
 $k_i$
, then this subcase is consistent with the lemma.
$k_i$
, then this subcase is consistent with the lemma.
Consider the remaining subcase, when
 $G$
is a subnetwork of
$G$
is a subnetwork of
 $\left\{ 2 X_1 \stackrel{m_1}{\leftarrow } X_1 + X_2 \stackrel{m_2}{\to } 2 X_2 \right\}$
, and so consists of only one or two reactions. If
$\left\{ 2 X_1 \stackrel{m_1}{\leftarrow } X_1 + X_2 \stackrel{m_2}{\to } 2 X_2 \right\}$
, and so consists of only one or two reactions. If
 $G$
has only one reaction, then Proposition 3.5 implies that
$G$
has only one reaction, then Proposition 3.5 implies that
 $\textrm{cap}_{\text{pos}}(G) = 0 \lt \infty$
(which is consistent with the lemma).
$\textrm{cap}_{\text{pos}}(G) = 0 \lt \infty$
(which is consistent with the lemma).
Now assume that
 $G$
has two reactions, that is,
$G$
has two reactions, that is,
 $G=\left\{ 2 X_1 \stackrel{m_1}{\leftarrow } X_1 + X_2 \stackrel{m_2}{\to } 2 X_2 \right\}$
. If
$G=\left\{ 2 X_1 \stackrel{m_1}{\leftarrow } X_1 + X_2 \stackrel{m_2}{\to } 2 X_2 \right\}$
. If
 $m_1 \neq m_2$
, then the ODE for
$m_1 \neq m_2$
, then the ODE for
 $X_1$
is
$X_1$
is
 $\frac{dx_1}{dt}= (m_1 - m_2) x_1 x_2$
and so there are no positive steady states. When
$\frac{dx_1}{dt}= (m_1 - m_2) x_1 x_2$
and so there are no positive steady states. When
 $m_1=m_2$
, the ODE for
$m_1=m_2$
, the ODE for
 $X_1$
becomes
$X_1$
becomes
 $\frac{dx_1}{dt}=0$
and it follows that
$\frac{dx_1}{dt}=0$
and it follows that
 $\textrm{cap}_{\text{pos}}(G) = \infty$
. Moreover, a simple computation shows that all the positive steady states are degenerate. This concludes the proof.
$\textrm{cap}_{\text{pos}}(G) = \infty$
. Moreover, a simple computation shows that all the positive steady states are degenerate. This concludes the proof.
Example 2.1 (continued). The network
 $\{ 0 \leftarrow A \to 2A,\, B \leftarrow A+B\}$
is one of the networks listed in Lemma 3.19.2(b), where
$\{ 0 \leftarrow A \to 2A,\, B \leftarrow A+B\}$
is one of the networks listed in Lemma 3.19.2(b), where
 $n=2$
.
$n=2$
.
Lemma 3.20 (One-dimensional bimolecular networks). If
 $G$
is a one-dimensional, bimolecular network, then
$G$
is a one-dimensional, bimolecular network, then
 $G$
is not nondegenerately multistationary.
$G$
is not nondegenerately multistationary.
Proof. Assume that
 $G$
is a one-dimensional network that is nondegenerately multistationary. We must show that
$G$
is a one-dimensional network that is nondegenerately multistationary. We must show that
 $G$
is not bimolecular. We claim that
$G$
is not bimolecular. We claim that
 $\textrm{cap}_{\text{pos}}(G)$
is finite. Indeed, if
$\textrm{cap}_{\text{pos}}(G)$
is finite. Indeed, if
 $\textrm{cap}_{\text{pos}}(G) = \infty$
, then Lemma 3.19 implies that all positive steady states are degenerate and so
$\textrm{cap}_{\text{pos}}(G) = \infty$
, then Lemma 3.19 implies that all positive steady states are degenerate and so
 $G$
is not nondegenerately multistationary, which is a contradiction. Hence,
$G$
is not nondegenerately multistationary, which is a contradiction. Hence,
 $\textrm{cap}_{\text{pos}}(G) \lt \infty$
.
$\textrm{cap}_{\text{pos}}(G) \lt \infty$
.
The hypotheses of part (1) of Lemma 3.18 are satisfied, that is,
 $\textrm{cap}_{\text{pos}}(G) \lt \infty$
, and
$\textrm{cap}_{\text{pos}}(G) \lt \infty$
, and
 $G$
is one-dimensional and multistationary. Therefore,
$G$
is one-dimensional and multistationary. Therefore,
 $G$
has an embedded one-species network with arrow diagram
$G$
has an embedded one-species network with arrow diagram
 $({\leftarrow}, \to)$
. Such an embedded network (e.g.
$({\leftarrow}, \to)$
. Such an embedded network (e.g.
 $\{0 \leftarrow A,\, 2A \to 3A\}$
) involves at least one complex that is not bimolecular, and so
$\{0 \leftarrow A,\, 2A \to 3A\}$
) involves at least one complex that is not bimolecular, and so
 $G$
is also not bimolecular.
$G$
is also not bimolecular.
Theorem 3.21 (Networks with up to two reversible reactions). If
 $G$
is a bimolecular reaction network that consists of one or two pairs of reversible reactions, then
$G$
is a bimolecular reaction network that consists of one or two pairs of reversible reactions, then
 $G$
is not multistationary.
$G$
is not multistationary.
Proof. Assume that
 $G$
is bimolecular and consists of one or two pairs of reversible reactions. Let
$G$
is bimolecular and consists of one or two pairs of reversible reactions. Let
 $p$
denote the number of pairs of reversible reactions (so,
$p$
denote the number of pairs of reversible reactions (so,
 $p=1$
or
$p=1$
or
 $p=2$
), and
$p=2$
), and
 $\ell$
the number of linkage classes. Let
$\ell$
the number of linkage classes. Let
 $s$
be the dimension of the stoichiometric subspace (so,
$s$
be the dimension of the stoichiometric subspace (so,
 $s=1$
or
$s=1$
or
 $s=2$
).
$s=2$
).
Case 1:
 $p=1$
. The deficiency of
$p=1$
. The deficiency of
 $G$
is
$G$
is
 $\delta = 2-1-1=0$
and
$\delta = 2-1-1=0$
and
 $G$
is weakly reversible. Hence, by the deficiency-zero theorem (Lemma 2.5) the network is not multistationary.
$G$
is weakly reversible. Hence, by the deficiency-zero theorem (Lemma 2.5) the network is not multistationary.
Case 2:
 $p=s=2$
. If
$p=s=2$
. If
 $\ell =1$
, then the deficiency is
$\ell =1$
, then the deficiency is
 $\delta = 3-1-2=0$
. If
$\delta = 3-1-2=0$
. If
 $\ell =2$
, then the deficiency is
$\ell =2$
, then the deficiency is
 $\delta =4-2-2=0$
. Therefore, for either value of
$\delta =4-2-2=0$
. Therefore, for either value of
 $\ell$
, the deficiency-zero theorem (Lemma 2.5) implies that the network is not multistationary.
$\ell$
, the deficiency-zero theorem (Lemma 2.5) implies that the network is not multistationary.
Case 3:
 $p=2$
and
$p=2$
and
 $s=1$
.
$s=1$
.
 $G$
is one-dimensional, bimolecular, and reversible. So, Lemma 3.19 implies that
$G$
is one-dimensional, bimolecular, and reversible. So, Lemma 3.19 implies that
 $\textrm{cap}_{\text{pos}}(G) \lt \infty$
. Now Lemma 3.18(2) yields
$\textrm{cap}_{\text{pos}}(G) \lt \infty$
. Now Lemma 3.18(2) yields
 $\textrm{cap}_{\text{pos}}(G) = \textrm{cap}_{\text{nondeg}}(G)$
, and Lemma 3.20 implies that
$\textrm{cap}_{\text{pos}}(G) = \textrm{cap}_{\text{nondeg}}(G)$
, and Lemma 3.20 implies that
 $\textrm{cap}_{\text{nondeg}}(G) \,\leq \, 1$
. Thus,
$\textrm{cap}_{\text{nondeg}}(G) \,\leq \, 1$
. Thus,
 $\textrm{cap}_{\text{pos}}(G) \leq 1$
, or, equivalently,
$\textrm{cap}_{\text{pos}}(G) \leq 1$
, or, equivalently,
 $G$
is non-multistationary.
$G$
is non-multistationary.
4. Main results on bimolecular networks
In this section, we establish minimal conditions for a bimolecular network to admit ACR and nondegenerate multistationarity simultaneously. These minimal conditions are in terms of the numbers of species, reactions, and reactant complexes. The main result is as follows.
Theorem 4.1 (Conditions for coexistence of ACR and nondegenerate multistationarity). Let
 $G$
be a bimolecular reaction network. If there exists a vector of positive rate constants
$G$
be a bimolecular reaction network. If there exists a vector of positive rate constants
 $\kappa ^*$
such that the mass-action system
$\kappa ^*$
such that the mass-action system
 $(G,\kappa ^*)$
has ACR and also is nondegenerately multistationary, then:
$(G,\kappa ^*)$
has ACR and also is nondegenerately multistationary, then:
-
1.
$G$
has at least three species.
-
2.
$G$
has at least three reactant complexes (and hence at least three reactions) and at least five complexes (reactant and product complexes).
-
3. If
$G$
is full-dimensional, then
$G$
has at least five reactant complexes (and hence at least five reactions).




This section is structured as follows. In Subsection 4.1, we prove part
 $(1)$
of Theorem 4.1 (specifically, part
$(1)$
of Theorem 4.1 (specifically, part
 $(1)$
follows from Proposition 4.3 and Theorem 4.8). Theorem 4.8 also analyses two-species bimolecular networks with ACR and degenerate multistationarity. Additionally, we characterise unconditional ACR in two-species bimolecular networks that are reversible (Theorem 4.4).
$(1)$
follows from Proposition 4.3 and Theorem 4.8). Theorem 4.8 also analyses two-species bimolecular networks with ACR and degenerate multistationarity. Additionally, we characterise unconditional ACR in two-species bimolecular networks that are reversible (Theorem 4.4).
Subsequently, in Subsection 4.2, we prove parts
 $(2)$
and
$(2)$
and
 $(3)$
of Theorem 4.1 (Theorem 4.9 and Proposition 4.12). We also consider full-dimensional, three-species, bimolecular networks with only four reactant complexes. By Theorem 4.1, such networks do not allow for the coexistence of ACR and nondegenerate multistationary. Nevertheless, ACR and degenerate multistationarity are possible, and we characterise the possible sets of reactant complexes of such networks (Proposition 4.15).
$(3)$
of Theorem 4.1 (Theorem 4.9 and Proposition 4.12). We also consider full-dimensional, three-species, bimolecular networks with only four reactant complexes. By Theorem 4.1, such networks do not allow for the coexistence of ACR and nondegenerate multistationary. Nevertheless, ACR and degenerate multistationarity are possible, and we characterise the possible sets of reactant complexes of such networks (Proposition 4.15).
4.1. Bimolecular networks with one or two species
This subsection characterises unconditional ACR in reversible networks with only one or two species (Proposition 4.3 and Theorem 4.4). Notably, our results show that such networks with unconditional ACR are not multistationary.
Remark 4.2 (Reversible networks). Our interest in reversible networks comes from our prior work with Joshi [Reference Joshi, Kaihnsa, Nguyen and Shiu18]. In that article, our results on multistationarity in randomly generated reaction networks arise from “lifting” this property from the following (multistationary) motif:
 \begin{align} & \left \{ B \leftrightarrows 0 \leftrightarrows A \leftrightarrows B+C, \quad C{\leftrightarrows } 2C \right \}. \end{align}
\begin{align} & \left \{ B \leftrightarrows 0 \leftrightarrows A \leftrightarrows B+C, \quad C{\leftrightarrows } 2C \right \}. \end{align}
The question arises, Are there multistationary motifs with fewer species, reactions, or complexes than the one in (29)? Discovering more motifs might aid in analysing the prevalence of multistationarity in random reaction networks generated by stochastic models besides the one in [Reference Joshi, Kaihnsa, Nguyen and Shiu18].
4.1.1. Networks with one species
When there is only one species, say
 $X_1$
, and the network is bimolecular, there are only three possible complexes:
$X_1$
, and the network is bimolecular, there are only three possible complexes:
 $0, X_1, 2X_1$
. Hence, every such network is a subnetwork of the following network:
$0, X_1, 2X_1$
. Hence, every such network is a subnetwork of the following network:
 \begin{align} G_{X_1} = \{0\leftrightarrows X_1\leftrightarrows 2X_1 \leftrightarrows 0\}. \end{align}
\begin{align} G_{X_1} = \{0\leftrightarrows X_1\leftrightarrows 2X_1 \leftrightarrows 0\}. \end{align}
Therefore, the possible reversible networks, besides
 $G_{X_1}$
itself, are listed here:
$G_{X_1}$
itself, are listed here:
 \begin{align} \{0\leftrightarrows X_1\},\, \{X_1\leftrightarrows 2X_1\},\, \{0\leftrightarrows 2X_1\},\, \{0\leftrightarrows X_1\leftrightarrows 2X_1\},\, \{X_1\leftrightarrows 0\leftrightarrows 2X_1\},\, \{0\leftrightarrows 2X_1\leftrightarrows X_1\}. \end{align}
\begin{align} \{0\leftrightarrows X_1\},\, \{X_1\leftrightarrows 2X_1\},\, \{0\leftrightarrows 2X_1\},\, \{0\leftrightarrows X_1\leftrightarrows 2X_1\},\, \{X_1\leftrightarrows 0\leftrightarrows 2X_1\},\, \{0\leftrightarrows 2X_1\leftrightarrows X_1\}. \end{align}
Proposition 4.3. Every bimolecular network in only one species is not nondegenerately multistationary. Every reversible, bimolecular network in only one species has unconditional ACR.
Proof. Let
 $G$
be a bimolecular network with only one species. Then,
$G$
be a bimolecular network with only one species. Then,
 $G$
is a subnetwork of
$G$
is a subnetwork of
 $G_{X_1}$
, in (30), and the first part of the proposition now follows readily from Lemmas 3.18–3.19.
$G_{X_1}$
, in (30), and the first part of the proposition now follows readily from Lemmas 3.18–3.19.
Next, assume
 $G$
is a reversible, bimolecular network with only one species. Then,
$G$
is a reversible, bimolecular network with only one species. Then,
 $G$
is either the network
$G$
is either the network
 $G_{X_1}$
or one of the networks listed in (31). Each of these networks is weakly reversible and satisfies the conditions of either the deficiency-zero or deficiency-one theorem (Lemmas 2.5–2.6). Thus, for every choice of positive rate constants
$G_{X_1}$
or one of the networks listed in (31). Each of these networks is weakly reversible and satisfies the conditions of either the deficiency-zero or deficiency-one theorem (Lemmas 2.5–2.6). Thus, for every choice of positive rate constants
 $\kappa$
, the mass-action system
$\kappa$
, the mass-action system
 $(G, \kappa )$
has a unique positive steady state. Hence,
$(G, \kappa )$
has a unique positive steady state. Hence,
 $G$
has unconditional ACR.
$G$
has unconditional ACR.
4.1.2. Reversible networks with two species
We now consider reversible, bimolecular networks with two species. Among such networks, the ones with unconditional ACR are characterised in the following result, which is the main result of this subsection.
Theorem 4.4 (Unconditional ACR in reversible, two-species networks). Let
 $G$
be a reversible, bimolecular reaction network with exactly two species (and at least one reaction).
$G$
be a reversible, bimolecular reaction network with exactly two species (and at least one reaction).
-
1. If
$G$
is full-dimensional, then the following are equivalent:
-
(a)
$G$
has unconditional ACR;
-
(b)
$G$
is not multistationary.
-
-
2. If
$G$
is one-dimensional, then the following are equivalent:
-
(a)
$G$
has unconditional ACR;
-
(b) Up to relabelling species,
$G$
is the (non-multistationary) network
$\{X_2 \leftrightarrows X_1+X_2\}$
.
-







Theorem 4.4 encompasses Propositions 4.5 and 4.6 below.
Proposition 4.5.
Let
 $G$
be a full-dimensional, reversible, bimolecular reaction network with exactly two species. Then, the following are equivalent:
$G$
be a full-dimensional, reversible, bimolecular reaction network with exactly two species. Then, the following are equivalent:
-
(a)
$G$
has unconditional ACR;
-
(b)
$G$
is not multistationary.


Proof. Let
 $G$
be a full-dimensional, reversible, bimolecular network with exactly
$G$
be a full-dimensional, reversible, bimolecular network with exactly
 $2$
species.
$2$
species.
We first prove
 $(b) \Rightarrow (a)$
. Assume that
$(b) \Rightarrow (a)$
. Assume that
 $G$
is non-multistationary, and let
$G$
is non-multistationary, and let
 $\kappa ^*$
be a choice of positive rate constants. Then, the mass-action system
$\kappa ^*$
be a choice of positive rate constants. Then, the mass-action system
 $(G,\kappa ^*)$
admits at most one positive steady state
$(G,\kappa ^*)$
admits at most one positive steady state
 $(x^*_1, x^*_2)$
(here the assumption that
$(x^*_1, x^*_2)$
(here the assumption that
 $G$
is full-dimensional is used). However, the fact that
$G$
is full-dimensional is used). However, the fact that
 $G$
is reversible guarantees at least one positive steady state (Remark 2.8). Hence,
$G$
is reversible guarantees at least one positive steady state (Remark 2.8). Hence,
 $(G, \kappa ^*)$
has a unique positive steady state
$(G, \kappa ^*)$
has a unique positive steady state
 $(x^*_1, x^*_2)$
and therefore has ACR in both species with ACR values
$(x^*_1, x^*_2)$
and therefore has ACR in both species with ACR values
 $x_1^*$
and
$x_1^*$
and
 $x_2^*$
, respectively. So,
$x_2^*$
, respectively. So,
 $G$
has unconditional ACR.
$G$
has unconditional ACR.
Next, we prove
 $(a) \Rightarrow (b)$
. Assume that
$(a) \Rightarrow (b)$
. Assume that
 $G$
has unconditional ACR. Let
$G$
has unconditional ACR. Let
 $\kappa ^*$
be a choice of positive rate constants. By relabelling species, if necessary, we may assume that the system
$\kappa ^*$
be a choice of positive rate constants. By relabelling species, if necessary, we may assume that the system
 $(G, \kappa ^*)$
has ACR in species
$(G, \kappa ^*)$
has ACR in species
 $X_1$
with some ACR value
$X_1$
with some ACR value
 $\alpha \gt 0$
. Every positive steady state of
$\alpha \gt 0$
. Every positive steady state of
 $(G, \kappa ^*)$
, therefore, has the form
$(G, \kappa ^*)$
, therefore, has the form
 $(\alpha, x_2^*)$
, where
$(\alpha, x_2^*)$
, where
 $x^*_2 \in \mathbb{R}_{\gt 0}$
. We must show that there is at most one such steady state.
$x^*_2 \in \mathbb{R}_{\gt 0}$
. We must show that there is at most one such steady state.
Write the mass-action ODEs of
 $(G, \kappa ^*)$
as
$(G, \kappa ^*)$
as
 $\frac{dx_1}{dt} = f_1$
and
$\frac{dx_1}{dt} = f_1$
and
 $\frac{dx_2}{dt}=f_2$
. Consider the univariate polynomial
$\frac{dx_2}{dt}=f_2$
. Consider the univariate polynomial
 $f_2|_{x_1 = \alpha } \in \mathbb{R}[x_2]$
. We claim that this polynomial is not the zero polynomial. To check this claim, assume for contradiction that
$f_2|_{x_1 = \alpha } \in \mathbb{R}[x_2]$
. We claim that this polynomial is not the zero polynomial. To check this claim, assume for contradiction that
 $f_2|_{x_1 = \alpha }$
is zero. As
$f_2|_{x_1 = \alpha }$
is zero. As
 $G$
is reversible, Remark 3.16 (which relies on Propositions 3.14–3.15) implies that
$G$
is reversible, Remark 3.16 (which relies on Propositions 3.14–3.15) implies that
 $X_2$
is a catalyst-only species of every reaction of
$X_2$
is a catalyst-only species of every reaction of
 $G$
. We conclude that
$G$
. We conclude that
 $G$
is not full-dimensional, which is a contradiction. Having shown that the univariate polynomial
$G$
is not full-dimensional, which is a contradiction. Having shown that the univariate polynomial
 $f_2|_{x_1 = \alpha }$
is nonzero, we now use Lemma 3.11 to conclude that
$f_2|_{x_1 = \alpha }$
is nonzero, we now use Lemma 3.11 to conclude that
 $(G, \kappa ^*)$
has at most one positive steady state of the form
$(G, \kappa ^*)$
has at most one positive steady state of the form
 $(\alpha, x_2^*)$
.
$(\alpha, x_2^*)$
.
Proposition 4.5 fails for networks that are not reversible. Indeed, a network without positive steady states (such as
 $\{ 0 \to A,\, 0 \to B\}$
) is not multistationary and also lacks unconditional ACR.
$\{ 0 \to A,\, 0 \to B\}$
) is not multistationary and also lacks unconditional ACR.
We end this subsection by considering two-species networks that are one-dimensional. Up to relabelling species, each such network is a subnetwork of exactly one of the following networks
 $G_i$
:
$G_i$
:
 \begin{align} G_1 &\,:\!=\, \{ 0 \leftrightarrows X_1+X_2 \} \notag \\[5pt] G_2 &\,:\!=\, \{X_1\leftrightarrows 2X_2\} \\[5pt] G_3 &\,:\!=\, \{ 0 \leftrightarrows X_1 \leftrightarrows 2X_1 \leftrightarrows 0, \, X_2 \leftrightarrows X_1+X_2\} \notag \\[5pt] G_4 &\,:\!=\, \{ 2X_1 \leftrightarrows X_1+X_2 \leftrightarrows 2X_2 \leftrightarrows 2X_1, \, X_1 \leftrightarrows X_2 \} \notag \end{align}
\begin{align} G_1 &\,:\!=\, \{ 0 \leftrightarrows X_1+X_2 \} \notag \\[5pt] G_2 &\,:\!=\, \{X_1\leftrightarrows 2X_2\} \\[5pt] G_3 &\,:\!=\, \{ 0 \leftrightarrows X_1 \leftrightarrows 2X_1 \leftrightarrows 0, \, X_2 \leftrightarrows X_1+X_2\} \notag \\[5pt] G_4 &\,:\!=\, \{ 2X_1 \leftrightarrows X_1+X_2 \leftrightarrows 2X_2 \leftrightarrows 2X_1, \, X_1 \leftrightarrows X_2 \} \notag \end{align}
The next result, which is part (2) of Theorem 4.4, states that among the reversible subnetworks of the networks
 $G_i$
listed in (32), only one has unconditional ACR (namely,
$G_i$
listed in (32), only one has unconditional ACR (namely,
 $\{X_2 \leftrightarrows X_1+X_2\}$
).
$\{X_2 \leftrightarrows X_1+X_2\}$
).
Proposition 4.6.
Let
 $G$
be a one-dimensional, reversible, bimolecular reaction network with exactly two species. Then, the following are equivalent:
$G$
be a one-dimensional, reversible, bimolecular reaction network with exactly two species. Then, the following are equivalent:
-
(a)
$G$
has unconditional ACR;
-
(b) Up to relabelling species,
$G$
is the (non-multistationary) network
$\{X_2 \leftrightarrows X_1+X_2\}$
.



Proof. Let
 $G$
be a two-species, one-dimensional, reversible, bimolecular reaction network. From the list (32), we know that
$G$
be a two-species, one-dimensional, reversible, bimolecular reaction network. From the list (32), we know that
 $G$
is a subnetwork of one of
$G$
is a subnetwork of one of
 $G_1$
,
$G_1$
,
 $G_2$
,
$G_2$
,
 $G_3$
, and
$G_3$
, and
 $G_4$
.
$G_4$
.
Assume
 $G$
is a subnetwork of
$G$
is a subnetwork of
 $G_1$
,
$G_1$
,
 $G_2$
, or
$G_2$
, or
 $G_4$
. Then,
$G_4$
. Then,
 $G \neq \{X_2 \leftrightarrows X_1+X_2\}$
and
$G \neq \{X_2 \leftrightarrows X_1+X_2\}$
and
 $G$
is not a subnetwork of
$G$
is not a subnetwork of
 $\{0 \leftrightarrows X_1 \leftrightarrows 2X_1 \leftrightarrows 0\}$
. So, it suffices to show
$\{0 \leftrightarrows X_1 \leftrightarrows 2X_1 \leftrightarrows 0\}$
. So, it suffices to show
 $G$
does not have unconditional ACR.
$G$
does not have unconditional ACR.
In networks
 $G_1$
,
$G_1$
,
 $G_2,$
and
$G_2,$
and
 $G_4$
, the reactant and product complexes of every reaction differ in both species
$G_4$
, the reactant and product complexes of every reaction differ in both species
 $X_1$
and
$X_1$
and
 $X_2$
. Also, all reactions in
$X_2$
. Also, all reactions in
 $G$
are reversible, so every complex of
$G$
are reversible, so every complex of
 $G$
is a reactant complex. We conclude that
$G$
is a reactant complex. We conclude that
 $G$
has two reactant complexes that differ in both species, and hence, Lemma 2.12 implies that
$G$
has two reactant complexes that differ in both species, and hence, Lemma 2.12 implies that
 $G$
does not have unconditional ACR.
$G$
does not have unconditional ACR.
We now consider the remaining case, when
 $G$
is a subnetwork of
$G$
is a subnetwork of
 $G_3$
. We write
$G_3$
. We write
 $G_3 = N_1 \cup N_2$
, where
$G_3 = N_1 \cup N_2$
, where
 $N_1\,:\!=\,\{0 \leftrightarrows X_1 \leftrightarrows 2X_1 \leftrightarrows 0 \}$
and
$N_1\,:\!=\,\{0 \leftrightarrows X_1 \leftrightarrows 2X_1 \leftrightarrows 0 \}$
and
 $N_2\,:\!=\,\{X_2 \leftrightarrows X_1+X_2\}$
. If
$N_2\,:\!=\,\{X_2 \leftrightarrows X_1+X_2\}$
. If
 $G=N_2$
, the mass-action ODEs are
$G=N_2$
, the mass-action ODEs are
 $d{x}_1/dt =\kappa _1x_2-\kappa _2x_1x_2$
and
$d{x}_1/dt =\kappa _1x_2-\kappa _2x_1x_2$
and
 $d{x}_2/dt =0$
, and so
$d{x}_2/dt =0$
, and so
 $G$
has unconditional ACR in species
$G$
has unconditional ACR in species
 $X_1$
with ACR value
$X_1$
with ACR value
 $\tfrac{\kappa _1}{\kappa _2}$
. If
$\tfrac{\kappa _1}{\kappa _2}$
. If
 $G$
is a subnetwork of
$G$
is a subnetwork of
 $N_1$
, then
$N_1$
, then
 $G$
has only one species (recall that every species of a network must take part in at least one reaction), which is a contradiction.
$G$
has only one species (recall that every species of a network must take part in at least one reaction), which is a contradiction.
Our final subcase is when
 $G$
contains reactions from both
$G$
contains reactions from both
 $N_1$
and
$N_1$
and
 $N_2$
. Then, from
$N_2$
. Then, from
 $N_2$
, the complex
$N_2$
, the complex
 $X_2$
is a reactant complex of
$X_2$
is a reactant complex of
 $G$
. Similarly, from
$G$
. Similarly, from
 $N_1$
, at least one of
$N_1$
, at least one of
 $X_1$
and
$X_1$
and
 $2X_1$
is a reactant complex of
$2X_1$
is a reactant complex of
 $G$
. Hence,
$G$
. Hence,
 $G$
contains two reactant complexes that differ in both species,
$G$
contains two reactant complexes that differ in both species,
 $X_1$
and
$X_1$
and
 $X_2$
. Therefore, Lemma 2.12 implies that
$X_2$
. Therefore, Lemma 2.12 implies that
 $G$
does not have unconditional ACR.
$G$
does not have unconditional ACR.
Finally, the fact that the network
 $\{X_2 \leftrightarrows X_1+X_2\}$
is non-multistationary follows easily from the deficiency-zero theorem (Lemma 2.5).
$\{X_2 \leftrightarrows X_1+X_2\}$
is non-multistationary follows easily from the deficiency-zero theorem (Lemma 2.5).
4.1.3. Irreversible networks with two species
In [Reference Meshkat, Shiu and Torres24], the following network was called a “degenerate-ACR network”, because it has unconditional ACR and yet every positive steady state is degenerate:
 \begin{align} \{ A+B \to B,\,A \to 2A \}. \end{align}
\begin{align} \{ A+B \to B,\,A \to 2A \}. \end{align}
This degeneracy arises from the fact that a single (one-dimensional) stoichiometric compatibility class consists entirely of steady states [Reference Meshkat, Shiu and Torres24, Example 2.12]. The main result of this subsection, Theorem 4.8 below, shows that only one additional two-species network exhibits both ACR and multistationarity for a nonzero-measure set of rate constants; this network is obtained by adding to (33) the reaction
 $A \to 0$
. Both networks, therefore, are one-dimensional, two-species networks.
$A \to 0$
. Both networks, therefore, are one-dimensional, two-species networks.
To prove Theorem 4.8, we need the following lemma, which concerns the network in (33) (and others as well).
Lemma 4.7.
Let
 $G$
be a subnetwork of the network
$G$
be a subnetwork of the network
 $\{X_1+X_2 \to X_2,\,0 \leftarrow X_1 \to 2X_1\}$
. Then:
$\{X_1+X_2 \to X_2,\,0 \leftarrow X_1 \to 2X_1\}$
. Then:
-
1. Every positive steady state (of every mass-action system defined by
$G$
) is degenerate.
-
2. Let
$\Sigma$
denote the set of vectors of positive rate constants
$\kappa$
for which the mass-action system
$(G, \kappa )$
both has ACR and is multistationary. If
$\Sigma$
has nonzero measure, then
$G$
is one of the following networks:
$\{X_1+X_2 \to X_2,\,X_1 \to 2X_1\}$
and
$\{X_1+X_2 \to X_2,\,0 \leftarrow X_1 \to 2X_1\}$
.








Proof. This result is straightforward to check by hand, so we only outline the steps, as follows. Assume
 $G$
is a subnetwork of
$G$
is a subnetwork of
 $\left\{X_1+X_2 \overset{k}\to X_2,\,0 \overset{\ell }\leftarrow X_1 \overset{m}\to 2X_1\right\}$
. If
$\left\{X_1+X_2 \overset{k}\to X_2,\,0 \overset{\ell }\leftarrow X_1 \overset{m}\to 2X_1\right\}$
. If
 $G$
admits a positive steady state,
$G$
admits a positive steady state,
 $G$
must contain the reaction
$G$
must contain the reaction
 $X_1 \overset{m}\to 2X_1$
. Hence, there are three subnetworks to consider:
$X_1 \overset{m}\to 2X_1$
. Hence, there are three subnetworks to consider:
-
1. If
$G= \left\{0 \overset{\ell }\leftarrow X_1 \overset{m}\to 2X_1\right\}$
, then
$\Sigma$
is empty. -
2. If
$G= \left\{X_1+X_2 \overset{k}\to X_2,\, X_1 \overset{m}\to 2X_1\right\}$
, then
$\Sigma = \left\{ (k,m) \in \mathbb{R}^2_{\gt 0} \right\}$
. -
3. If
$G= \left\{X_1+X_2 \overset{k}\to X_2,\,0 \overset{\ell }\leftarrow X_1 \overset{m}\to 2X_1\right\}$
, then
$\Sigma = \left\{ (k,\ell,m) \in \mathbb{R}^3_{\gt 0} \mid m \gt \ell \right\}$
.






In cases (2) and (3), the set
 $\Sigma$
has nonzero measure. Finally, for all three of these networks, every positive steady state is degenerate (some of these networks are also covered by Lemma 3.19).
$\Sigma$
has nonzero measure. Finally, for all three of these networks, every positive steady state is degenerate (some of these networks are also covered by Lemma 3.19).
Theorem 4.8.
Let
 $G$
be a bimolecular reaction network with exactly two species,
$G$
be a bimolecular reaction network with exactly two species,
 $X_1$
and
$X_1$
and
 $X_2$
. Let
$X_2$
. Let
 $\Sigma$
denote the set of vectors of positive rate constants
$\Sigma$
denote the set of vectors of positive rate constants
 $\kappa$
for which the mass-action system
$\kappa$
for which the mass-action system
 $(G, \kappa )$
both has ACR in species
$(G, \kappa )$
both has ACR in species
 $X_2$
and is multistationary. Then:
$X_2$
and is multistationary. Then:
-
1. For every
$\kappa ^* \in \Sigma$
, every positive steady state of
$(G, \kappa ^*)$
is degenerate.
-
2. If
$\Sigma$
has nonzero measure, then
$G$
is one of the following networks:
$\{X_1+X_2 \to X_2,\,X_1 \to 2X_1\}$
and
$\{X_1+X_2 \to X_2,\,0 \leftarrow X_1 \to 2X_1\}$
.






Proof. Assume that
 $G$
is bimolecular and has exactly two species. If
$G$
is bimolecular and has exactly two species. If
 $\Sigma$
is empty (for instance, if
$\Sigma$
is empty (for instance, if
 $G$
has no reactions), then there is nothing to prove. Accordingly, assume that
$G$
has no reactions), then there is nothing to prove. Accordingly, assume that
 $\Sigma$
is nonempty (and in particular
$\Sigma$
is nonempty (and in particular
 $G$
has at least one reaction).
$G$
has at least one reaction).
We first claim that
 $G$
has a reaction in which
$G$
has a reaction in which
 $X_1$
is a non-catalyst-only species. To prove this claim, assume for contradiction that
$X_1$
is a non-catalyst-only species. To prove this claim, assume for contradiction that
 $X_1$
is a catalyst-only species. Then, the stoichiometric compatibility classes are defined by the equations
$X_1$
is a catalyst-only species. Then, the stoichiometric compatibility classes are defined by the equations
 $x_1=T$
, for
$x_1=T$
, for
 $T\gt 0$
(we are also using the fact that
$T\gt 0$
(we are also using the fact that
 $G$
has at least one reaction). But this does not allow for multistationarity and ACR in
$G$
has at least one reaction). But this does not allow for multistationarity and ACR in
 $X_2$
to coexist, because two positive steady states in the same compatibility class would have the form
$X_2$
to coexist, because two positive steady states in the same compatibility class would have the form
 $(T,y)$
and
$(T,y)$
and
 $(T,z)$
, with
$(T,z)$
, with
 $y\neq z$
, which contradicts the assumption of ACR in
$y\neq z$
, which contradicts the assumption of ACR in
 $X_2$
. So, the claim holds.
$X_2$
. So, the claim holds.
For an arbitrary vector
 $\kappa$
of positive rate constants, let
$\kappa$
of positive rate constants, let
 $f_{\kappa,1}$
and
$f_{\kappa,1}$
and
 $f_{\kappa,2}$
denote the right-hand sides (for species
$f_{\kappa,2}$
denote the right-hand sides (for species
 $X_1$
and
$X_1$
and
 $X_2$
, respectively) of the mass-action ODE system of
$X_2$
, respectively) of the mass-action ODE system of
 $(G,\kappa )$
. Consider the following partition of
$(G,\kappa )$
. Consider the following partition of
 $\Sigma$
:
$\Sigma$
:
 \begin{align*} \Sigma = \left ( \Sigma \cap \{\kappa \mid f_{\kappa,1}= 0\} \right ) \cup \left ( \Sigma \cap \{\kappa \mid f_{\kappa,1} \neq 0\} \right ) \,=\!:\, \Sigma _0 \cup \Sigma _1. \end{align*}
\begin{align*} \Sigma = \left ( \Sigma \cap \{\kappa \mid f_{\kappa,1}= 0\} \right ) \cup \left ( \Sigma \cap \{\kappa \mid f_{\kappa,1} \neq 0\} \right ) \,=\!:\, \Sigma _0 \cup \Sigma _1. \end{align*}
By construction,
 $\Sigma _0\cap \Sigma _1=\emptyset$
. We first analyse
$\Sigma _0\cap \Sigma _1=\emptyset$
. We first analyse
 $\Sigma _0$
. If
$\Sigma _0$
. If
 $\Sigma _0$
is empty, then skip ahead to our analysis of
$\Sigma _0$
is empty, then skip ahead to our analysis of
 $\Sigma _1$
. Accordingly, assume
$\Sigma _1$
. Accordingly, assume
 $\Sigma _0$
is nonempty, and let
$\Sigma _0$
is nonempty, and let
 $\kappa ^* \in \Sigma _0$
. We must show that every positive steady state of
$\kappa ^* \in \Sigma _0$
. We must show that every positive steady state of
 $(G,\kappa ^*)$
is degenerate.
$(G,\kappa ^*)$
is degenerate.
We claim that
 $G$
is two-dimensional (assuming that
$G$
is two-dimensional (assuming that
 $\Sigma _0$
is nonempty). We prove this claim as follows. We saw that
$\Sigma _0$
is nonempty). We prove this claim as follows. We saw that
 $G$
contains a reaction in which
$G$
contains a reaction in which
 $X_1$
is a non-catalyst-only species, so Proposition 3.14 implies that for
$X_1$
is a non-catalyst-only species, so Proposition 3.14 implies that for
 $j=0$
or
$j=0$
or
 $j=2$
(or both, where we are using Notation 3.13) our network
$j=2$
(or both, where we are using Notation 3.13) our network
 $G$
contains the reaction
$G$
contains the reaction
 $X_1+X_j \to 2X_1$
and at least one reaction of the form
$X_1+X_j \to 2X_1$
and at least one reaction of the form
 $X_1+X_j \to \star$
, where
$X_1+X_j \to \star$
, where
 $\star$
is a complex not involving
$\star$
is a complex not involving
 $X_1$
. Consider the subcase
$X_1$
. Consider the subcase
 $j=0$
. If some
$j=0$
. If some
 $\star$
involves
$\star$
involves
 $X_2$
, then
$X_2$
, then
 $G$
contains
$G$
contains
 $X_1 \to 2X_1$
and
$X_1 \to 2X_1$
and
 $X_1 \to \star$
, which yield linearly independent reaction vectors and so
$X_1 \to \star$
, which yield linearly independent reaction vectors and so
 $G$
is two-dimensional. If none of the complexes
$G$
is two-dimensional. If none of the complexes
 $\star$
involve
$\star$
involve
 $X_2$
, then
$X_2$
, then
 $G$
must contain additional reactions in which
$G$
must contain additional reactions in which
 $X_2$
is not a catalyst-only species (to avoid
$X_2$
is not a catalyst-only species (to avoid
 $f_2=0$
), and so again
$f_2=0$
), and so again
 $G$
is two-dimensional. The subcase
$G$
is two-dimensional. The subcase
 $j=2$
is similar.
$j=2$
is similar.
Next, as
 $G$
is two-dimensional and
$G$
is two-dimensional and
 $f_{\kappa ^*,1}= 0$
, Corollary 3.4 implies that every positive steady state of
$f_{\kappa ^*,1}= 0$
, Corollary 3.4 implies that every positive steady state of
 $(G,\kappa ^*)$
is degenerate, as desired. Additionally, as
$(G,\kappa ^*)$
is degenerate, as desired. Additionally, as
 $X_1$
is a non-catalyst-only species and (for all
$X_1$
is a non-catalyst-only species and (for all
 $\kappa \in \Sigma _0$
)
$\kappa \in \Sigma _0$
)
 $f_{\kappa,1}=0$
, Proposition 3.14 implies that there is a nontrivial linear relation that every
$f_{\kappa,1}=0$
, Proposition 3.14 implies that there is a nontrivial linear relation that every
 $\kappa \in \Sigma _0$
satisfies. Hence,
$\kappa \in \Sigma _0$
satisfies. Hence,
 $\Sigma _0$
has zero measure.
$\Sigma _0$
has zero measure.
To complete the proof, it suffices to show the following about the set
 $\Sigma _1$
: (1) For every
$\Sigma _1$
: (1) For every
 $\kappa ^* \in \Sigma _1$
, every positive steady state of
$\kappa ^* \in \Sigma _1$
, every positive steady state of
 $(G, \kappa ^*)$
is degenerate, and (2) if
$(G, \kappa ^*)$
is degenerate, and (2) if
 $\Sigma _1$
has nonzero measure, then
$\Sigma _1$
has nonzero measure, then
 $G=\{X_1+X_2 \to X_2,\,X_1 \to 2X_1\}$
or
$G=\{X_1+X_2 \to X_2,\,X_1 \to 2X_1\}$
or
 $G=\{X_1+X_2 \to X_2,\,0 \leftarrow X_1 \to 2X_1\}$
.
$G=\{X_1+X_2 \to X_2,\,0 \leftarrow X_1 \to 2X_1\}$
.
Assume
 $\Sigma _1$
is nonempty (otherwise, there is nothing to prove). We introduce the following notation: for
$\Sigma _1$
is nonempty (otherwise, there is nothing to prove). We introduce the following notation: for
 $\widetilde \kappa \in \Sigma _1$
, let
$\widetilde \kappa \in \Sigma _1$
, let
 $\beta (\widetilde \kappa )$
denote the ACR value for
$\beta (\widetilde \kappa )$
denote the ACR value for
 $X_2$
.
$X_2$
.
We now claim the following: For every
 $\widetilde \kappa \in \Sigma _1$
, the univariate polynomial
$\widetilde \kappa \in \Sigma _1$
, the univariate polynomial
 $f_{\widetilde \kappa,_1}|_{x_2=\beta (\widetilde \kappa ) }$
is the zero polynomial. To verify this claim, we first note that
$f_{\widetilde \kappa,_1}|_{x_2=\beta (\widetilde \kappa ) }$
is the zero polynomial. To verify this claim, we first note that
 $f_{\widetilde \kappa,_1}|_{x_2=\beta (\widetilde \kappa ) }$
has at least two positive roots (as
$f_{\widetilde \kappa,_1}|_{x_2=\beta (\widetilde \kappa ) }$
has at least two positive roots (as
 $(G, \widetilde \kappa )$
is multistationarity), so the polynomial
$(G, \widetilde \kappa )$
is multistationarity), so the polynomial
 $f_{\widetilde \kappa,_1}|_{x_2=\beta (\widetilde \kappa ) }$
, if nonzero, must have at least two sign changes (by Descartes’ rule of signs). However, by Lemma 3.11, the polynomial
$f_{\widetilde \kappa,_1}|_{x_2=\beta (\widetilde \kappa ) }$
, if nonzero, must have at least two sign changes (by Descartes’ rule of signs). However, by Lemma 3.11, the polynomial
 $f_{\widetilde \kappa,_1}|_{x_2=\beta (\widetilde \kappa ) }$
has at most one sign change, and so the claim holds.
$f_{\widetilde \kappa,_1}|_{x_2=\beta (\widetilde \kappa ) }$
has at most one sign change, and so the claim holds.
We now know that for every
 $\kappa ^* \in \Sigma _1$
, we have
$\kappa ^* \in \Sigma _1$
, we have
 $f_{\kappa ^*,_1} \neq 0$
, but
$f_{\kappa ^*,_1} \neq 0$
, but
 $f_{ \kappa ^*,_1}|_{x_2=\beta (\widetilde \kappa ^*) }=0$
. Hence,
$f_{ \kappa ^*,_1}|_{x_2=\beta (\widetilde \kappa ^*) }=0$
. Hence,
 $G$
has at least one reaction in which
$G$
has at least one reaction in which
 $X_1$
is a non-catalyst-only reaction and (by Proposition 3.15) every such reaction must be one of the
$X_1$
is a non-catalyst-only reaction and (by Proposition 3.15) every such reaction must be one of the
 $8$
reactions displayed here:
$8$
reactions displayed here:
 \begin{align} 0 \overset{ \kappa _{4,1} } \longleftarrow X_1 \overset{\kappa _1} \longrightarrow 2X_1 \overset{\kappa _2} \longleftarrow X_1+X_2 \overset{\kappa _{3,1} } \longrightarrow 0, \quad X_2 \overset{\kappa _{4,2}} \longleftarrow X_1 \overset{\kappa _{4,3}} \longrightarrow 2X_2, \quad X_2 \overset{\kappa _{3,2}} \longleftarrow X_1+X_2 \overset{\kappa _{3,3}} \longrightarrow 2X_2 \end{align}
\begin{align} 0 \overset{ \kappa _{4,1} } \longleftarrow X_1 \overset{\kappa _1} \longrightarrow 2X_1 \overset{\kappa _2} \longleftarrow X_1+X_2 \overset{\kappa _{3,1} } \longrightarrow 0, \quad X_2 \overset{\kappa _{4,2}} \longleftarrow X_1 \overset{\kappa _{4,3}} \longrightarrow 2X_2, \quad X_2 \overset{\kappa _{3,2}} \longleftarrow X_1+X_2 \overset{\kappa _{3,3}} \longrightarrow 2X_2 \end{align}
For every
 $\kappa ^* \in \Sigma _1$
, Proposition 3.15 yields the following ACR value formula:
$\kappa ^* \in \Sigma _1$
, Proposition 3.15 yields the following ACR value formula:
 \begin{align} \beta (\kappa ^*) = \frac{ \kappa ^*_{4 \bullet } - \kappa ^*_1}{\kappa ^*_2 - \kappa ^*_{3 \bullet } }, \end{align}
\begin{align} \beta (\kappa ^*) = \frac{ \kappa ^*_{4 \bullet } - \kappa ^*_1}{\kappa ^*_2 - \kappa ^*_{3 \bullet } }, \end{align}
where
 $\kappa ^*_{3 \bullet } \,:\!=\, \kappa ^*_{3, 1 } + \kappa ^*_{3, 2 } + \kappa ^*_{3, 3 }$
and
$\kappa ^*_{3 \bullet } \,:\!=\, \kappa ^*_{3, 1 } + \kappa ^*_{3, 2 } + \kappa ^*_{3, 3 }$
and
 $\kappa ^*_{4 \bullet }\,:\!=\,\kappa ^*_{4, 1 } + \kappa ^*_{4, 2 } + \kappa ^*_{4, 3 }$
. For reactions in (34) that are not in
$\kappa ^*_{4 \bullet }\,:\!=\,\kappa ^*_{4, 1 } + \kappa ^*_{4, 2 } + \kappa ^*_{4, 3 }$
. For reactions in (34) that are not in
 $G$
, the corresponding rate constants,
$G$
, the corresponding rate constants,
 $\kappa ^*_i$
or
$\kappa ^*_i$
or
 $\kappa ^*_{ij}$
, are set to 0.
$\kappa ^*_{ij}$
, are set to 0.
Next, the possible reactions in which
 $X_1$
is a catalyst-only species are as follows:
$X_1$
is a catalyst-only species are as follows:
 \begin{align} 0 \overset{\kappa _5}{\underset{\kappa _6} \rightleftarrows } X_2 \overset{\kappa _7}{\underset{\kappa _8} \rightleftarrows } 2X_2 \overset{\kappa _9}{\underset{\kappa _{10}} \rightleftarrows } 0, \quad \quad X_1 \overset{\kappa _{11}}{\underset{\kappa _{12}} \rightleftarrows } X_1+ X_2 \end{align}
\begin{align} 0 \overset{\kappa _5}{\underset{\kappa _6} \rightleftarrows } X_2 \overset{\kappa _7}{\underset{\kappa _8} \rightleftarrows } 2X_2 \overset{\kappa _9}{\underset{\kappa _{10}} \rightleftarrows } 0, \quad \quad X_1 \overset{\kappa _{11}}{\underset{\kappa _{12}} \rightleftarrows } X_1+ X_2 \end{align}
We proceed by considering three subcases, based in part on whether
 $f_{\kappa,2}$
(which is a polynomial in the unknowns
$f_{\kappa,2}$
(which is a polynomial in the unknowns
 $x_1$
,
$x_1$
,
 $x_2$
, and
$x_2$
, and
 $\kappa$
) is zero:
$\kappa$
) is zero:
-
(a)
$f_{\kappa,2}=0$
, and
$X_2$
is a catalyst-only species in every reaction of
$G$
, -
(b)
$f_{\kappa,2}=0$
, and
$X_2$
is a non-catalyst-only species in some reaction of
$G$
, or -
(c)
$f_{\kappa,2} \neq 0$
.







We first consider subcase (a). By inspecting reactions in (34) and (36), we conclude that
 $G$
must be a subnetwork of
$G$
must be a subnetwork of
 $\{X_1+X_2 \to X_2,\,0 \leftarrow X_1 \to 2X_1\}$
. This subcase is done by Lemma 4.7.
$\{X_1+X_2 \to X_2,\,0 \leftarrow X_1 \to 2X_1\}$
. This subcase is done by Lemma 4.7.
Next, we examine subcase (b). Let
 $G_1\,:\!=\,\{X_1+X_2 \to X_2,\,0 \leftarrow X_1 \to 2X_1\}$
,
$G_1\,:\!=\,\{X_1+X_2 \to X_2,\,0 \leftarrow X_1 \to 2X_1\}$
,
 $G_2\,:\!=\, \{0\leftarrow X_2 \to 2X_2\}$
, and
$G_2\,:\!=\, \{0\leftarrow X_2 \to 2X_2\}$
, and
 $G_3\,:\!=\,\{0\leftarrow X_1+X_2\to 2X_2,\, X_1 \leftarrow X_1+ X_2 \to 2X_1 \}$
. By Proposition 3.14 (and by inspecting reactions in (34) and (36)),
$G_3\,:\!=\,\{0\leftarrow X_1+X_2\to 2X_2,\, X_1 \leftarrow X_1+ X_2 \to 2X_1 \}$
. By Proposition 3.14 (and by inspecting reactions in (34) and (36)),
 $G$
must be a subnetwork of
$G$
must be a subnetwork of
 $G_1\cup G_2 \cup G_3$
with at least one reaction in
$G_1\cup G_2 \cup G_3$
with at least one reaction in
 $G_2 \cup G_3$
. Moreover, there is a nontrivial linear relation in the rate constants that holds for all
$G_2 \cup G_3$
. Moreover, there is a nontrivial linear relation in the rate constants that holds for all
 $\kappa \in \Sigma _1$
. It follows that
$\kappa \in \Sigma _1$
. It follows that
 $\Sigma _1$
is contained in the hyperplane defined by this linear relation and hence has zero measure.
$\Sigma _1$
is contained in the hyperplane defined by this linear relation and hence has zero measure.
Let
 $\kappa ^* \in \Sigma _1$
. By examining
$\kappa ^* \in \Sigma _1$
. By examining
 $G_1 \cup G_2 \cup G_3$
, we see that the possible reactants of
$G_1 \cup G_2 \cup G_3$
, we see that the possible reactants of
 $G$
are
$G$
are
 $X_1,\,X_2,\,X_1+X_2$
. Next,
$X_1,\,X_2,\,X_1+X_2$
. Next,
 $G$
has at least
$G$
has at least
 $2$
reactants (as otherwise, Proposition 3.5 would imply that
$2$
reactants (as otherwise, Proposition 3.5 would imply that
 $G$
admits no positive steady states). Hence, by inspection,
$G$
admits no positive steady states). Hence, by inspection,
 $G$
either is full-dimensional or is a subnetwork of
$G$
either is full-dimensional or is a subnetwork of
 $\{ 0 \leftarrow X_2 \to 2X_2,\, X_1+X_2 \to X_1\}$
, which we already saw in Example 2.1 (where
$\{ 0 \leftarrow X_2 \to 2X_2,\, X_1+X_2 \to X_1\}$
, which we already saw in Example 2.1 (where
 $A=X_2$
and
$A=X_2$
and
 $B=X_1$
) has ACR in
$B=X_1$
) has ACR in
 $X_1$
but not in
$X_1$
but not in
 $X_2$
(and the analysis of its subnetworks is similar). Hence,
$X_2$
(and the analysis of its subnetworks is similar). Hence,
 $G$
is full-dimensional, and so Corollary 3.4 (and the fact that
$G$
is full-dimensional, and so Corollary 3.4 (and the fact that
 $f_{\kappa ^*,2}=0$
) implies that every positive steady state of
$f_{\kappa ^*,2}=0$
) implies that every positive steady state of
 $(G,\kappa ^*)$
is degenerate.
$(G,\kappa ^*)$
is degenerate.
Consider subcase (c). Let
 $\kappa ^* \in \Sigma _1$
(so, in particular,
$\kappa ^* \in \Sigma _1$
(so, in particular,
 $f_{\kappa ^*,2} \neq 0$
). We claim that
$f_{\kappa ^*,2} \neq 0$
). We claim that
 $f_{\kappa ^*,2}|_{x_2=\beta (\kappa ^*)} = 0$
. To see this, observe that, in the reactions (34) and (36), the complex
$f_{\kappa ^*,2}|_{x_2=\beta (\kappa ^*)} = 0$
. To see this, observe that, in the reactions (34) and (36), the complex
 $2X_1$
appears only as a product, never as a reactant. Hence,
$2X_1$
appears only as a product, never as a reactant. Hence,
 $f_{\kappa ^*,2}|_{x_2=\beta (\kappa ^*)}$
(which is a univariate polynomial in
$f_{\kappa ^*,2}|_{x_2=\beta (\kappa ^*)}$
(which is a univariate polynomial in
 $x_1$
) has degree at most 1. However, the fact that
$x_1$
) has degree at most 1. However, the fact that
 $(G,\kappa ^*)$
is multistationary implies that
$(G,\kappa ^*)$
is multistationary implies that
 $f_{\kappa ^*,2}|_{x_2=\beta (\kappa ^*)}$
has two or more positive roots. Hence,
$f_{\kappa ^*,2}|_{x_2=\beta (\kappa ^*)}$
has two or more positive roots. Hence,
 $f_{\kappa ^*,2}|_{x_2=\beta (\kappa ^*)}$
is the zero polynomial.
$f_{\kappa ^*,2}|_{x_2=\beta (\kappa ^*)}$
is the zero polynomial.
Now we show that every positive steady state of
 $(G,\kappa ^*)$
is degenerate. Such a steady state has the form
$(G,\kappa ^*)$
is degenerate. Such a steady state has the form
 $(p,\beta )$
, and we also know that
$(p,\beta )$
, and we also know that
 $f_{\kappa ^*,1}|_{x_2=\beta (\kappa ^*)} = f_{\kappa ^*,2}|_{x_2=\beta (\kappa ^*)}=0$
. Hence,
$f_{\kappa ^*,1}|_{x_2=\beta (\kappa ^*)} = f_{\kappa ^*,2}|_{x_2=\beta (\kappa ^*)}=0$
. Hence,
 $(x_2-\beta (\kappa ^*))$
divides both
$(x_2-\beta (\kappa ^*))$
divides both
 $f_{\kappa ^*,1}$
and
$f_{\kappa ^*,1}$
and
 $f_{\kappa ^*,2}$
. Consequently, the derivatives of
$f_{\kappa ^*,2}$
. Consequently, the derivatives of
 $f_{\kappa ^*,1}$
and
$f_{\kappa ^*,1}$
and
 $f_{\kappa ^*,2}$
with respect to
$f_{\kappa ^*,2}$
with respect to
 $x_1$
at
$x_1$
at
 $(p,\beta )$
are both zero. It follows that the first column of the
$(p,\beta )$
are both zero. It follows that the first column of the
 $2 \times 2$
Jacobian matrix, when evaluated at
$2 \times 2$
Jacobian matrix, when evaluated at
 $(p,\beta )$
, is the zero column. Hence, if the stoichiometric subspace of
$(p,\beta )$
, is the zero column. Hence, if the stoichiometric subspace of
 $G$
, which we denote by
$G$
, which we denote by
 $S$
, is two-dimensional, then
$S$
, is two-dimensional, then
 $(p,\beta )$
is degenerate.
$(p,\beta )$
is degenerate.
We now assume
 $\text{dim}(S)=1$
(and aim to reach a contradiction). Recall that
$\text{dim}(S)=1$
(and aim to reach a contradiction). Recall that
 $G$
contains at least one reaction from those in (34), so in order for
$G$
contains at least one reaction from those in (34), so in order for
 $\text{dim}(S)=1$
it must be that
$\text{dim}(S)=1$
it must be that
 $G$
contains no reaction from (36). Hence, from the expression for
$G$
contains no reaction from (36). Hence, from the expression for
 $f_2$
(which we know is not zero), in (37), the only possible reactions in
$f_2$
(which we know is not zero), in (37), the only possible reactions in
 $G$
are the ones labelled by
$G$
are the ones labelled by
 $\kappa _2, \kappa _{3,3}, \kappa _{4,2},\kappa _{4,3}$
. Hence, the one-dimensional network
$\kappa _2, \kappa _{3,3}, \kappa _{4,2},\kappa _{4,3}$
. Hence, the one-dimensional network
 $G$
is either the network
$G$
is either the network
 $\{ X_1 \overset{\kappa _{4,3}}\longrightarrow 2X_2 \}$
or a subnetwork of
$\{ X_1 \overset{\kappa _{4,3}}\longrightarrow 2X_2 \}$
or a subnetwork of
 $\left\{2X_1 \overset{\kappa _2} \longleftarrow X_1+X_2 \overset{\kappa _{3,3} } \longrightarrow 2X_2, \, X_1 \overset{\kappa _{4,2} } \longrightarrow X_2 \right\}$
. Now it is straightforward to check that
$\left\{2X_1 \overset{\kappa _2} \longleftarrow X_1+X_2 \overset{\kappa _{3,3} } \longrightarrow 2X_2, \, X_1 \overset{\kappa _{4,2} } \longrightarrow X_2 \right\}$
. Now it is straightforward to check that
 $G$
is not multistationary, which is a contradiction.
$G$
is not multistationary, which is a contradiction.
To complete the proof, it suffices to show that, in subcase (c), the set
 $\Sigma _1$
has measure zero. Accordingly, let
$\Sigma _1$
has measure zero. Accordingly, let
 $\kappa \in \Sigma _1$
. As noted earlier, the ACR value of
$\kappa \in \Sigma _1$
. As noted earlier, the ACR value of
 $X_2$
in
$X_2$
in
 $(G,\kappa )$
is
$(G,\kappa )$
is
 $\beta (\kappa ) = \frac{ \kappa _{4 \bullet } - \kappa _1}{\kappa _2 - \kappa _{3 \bullet } }$
. From (34) and (36), the right-hand side of the mass-action ODE for
$\beta (\kappa ) = \frac{ \kappa _{4 \bullet } - \kappa _1}{\kappa _2 - \kappa _{3 \bullet } }$
. From (34) and (36), the right-hand side of the mass-action ODE for
 $(G,\kappa )$
has the following form (with rate constants set to
$(G,\kappa )$
has the following form (with rate constants set to
 $0$
for reactions not in
$0$
for reactions not in
 $G$
):
$G$
):
 \begin{align} f_{\kappa,2} = ( \kappa _{3,3} - \kappa _2 - \kappa _{12}) x_1 x_2 + (\kappa _{11} + \kappa _{4,2} + 2 \kappa _{4,3}) x_1 - (\kappa _8 + 2 \kappa _9) x_2^2 + (\kappa _7 - \kappa _6) x_2 + (\kappa _5 + 2 \kappa _{10}). \end{align}
\begin{align} f_{\kappa,2} = ( \kappa _{3,3} - \kappa _2 - \kappa _{12}) x_1 x_2 + (\kappa _{11} + \kappa _{4,2} + 2 \kappa _{4,3}) x_1 - (\kappa _8 + 2 \kappa _9) x_2^2 + (\kappa _7 - \kappa _6) x_2 + (\kappa _5 + 2 \kappa _{10}). \end{align}
By assumption, at least one of the rate constants (the
 $\kappa _i$
and
$\kappa _i$
and
 $\kappa _{i,j}$
) in (37) is nonzero. By our earlier arguments, at the beginning of subcase (c), we conclude that
$\kappa _{i,j}$
) in (37) is nonzero. By our earlier arguments, at the beginning of subcase (c), we conclude that
 $f_{\kappa,2}|_{x_2=\beta (\kappa )} = 0$
. Hence, the linear and constant terms of
$f_{\kappa,2}|_{x_2=\beta (\kappa )} = 0$
. Hence, the linear and constant terms of
 $f_{\kappa,2}|_{x_2=\beta (\kappa )}$
are both
$f_{\kappa,2}|_{x_2=\beta (\kappa )}$
are both
 $0$
, which, using (37), translates as follows:
$0$
, which, using (37), translates as follows:
 \begin{align} ( \kappa _{3,3} - \kappa _2 - \kappa _{12}) \frac{ \kappa _{4 \bullet } - \kappa _1}{\kappa _2 - \kappa _{3 \bullet } } + (\kappa _{11} + \kappa _{4,2} + 2 \kappa _{4,3}) &= 0 \quad \textrm{and} \notag\\[5pt] - (\kappa _8 + 2 \kappa _9) \left ( \frac{ \kappa _{4 \bullet } - \kappa _1}{\kappa _2 - \kappa _{3 \bullet } } \right )^2 + (\kappa _7 - \kappa _6) \frac{ \kappa _{4 \bullet } - \kappa _1}{\kappa _2 - \kappa _{3 \bullet } } + (\kappa _5 + 2 \kappa _{10}) &= 0. \end{align}
\begin{align} ( \kappa _{3,3} - \kappa _2 - \kappa _{12}) \frac{ \kappa _{4 \bullet } - \kappa _1}{\kappa _2 - \kappa _{3 \bullet } } + (\kappa _{11} + \kappa _{4,2} + 2 \kappa _{4,3}) &= 0 \quad \textrm{and} \notag\\[5pt] - (\kappa _8 + 2 \kappa _9) \left ( \frac{ \kappa _{4 \bullet } - \kappa _1}{\kappa _2 - \kappa _{3 \bullet } } \right )^2 + (\kappa _7 - \kappa _6) \frac{ \kappa _{4 \bullet } - \kappa _1}{\kappa _2 - \kappa _{3 \bullet } } + (\kappa _5 + 2 \kappa _{10}) &= 0. \end{align}
It follows that
 $\Sigma _1$
is constrained by the equations (38), at least one of which is nontrivial. Hence,
$\Sigma _1$
is constrained by the equations (38), at least one of which is nontrivial. Hence,
 $\Sigma _1$
is contained in a hypersurface and so has measure zero.
$\Sigma _1$
is contained in a hypersurface and so has measure zero.
4.2. Bimolecular networks with at least three species
In the previous subsection, we showed that a bimolecular network must have at least three species in order for ACR and nondegenerate multistationarity to coexist. Consequently, this subsection focuses on bimolecular networks with at least three species. We prove that the coexistence of ACR and nondegenerate multistationarity requires a minimum of three reactant complexes and a minimum of five complexes (Theorem 4.9). The remainder of this subsection focuses on a family of networks with
 $n$
species and
$n$
species and
 $n$
reactants, for which ACR and nondegenerate multistationarity coexist (Section 4.2.1), and then analyses full-dimensional networks with
$n$
reactants, for which ACR and nondegenerate multistationarity coexist (Section 4.2.1), and then analyses full-dimensional networks with
 $3$
species (Section 4.2.2).
$3$
species (Section 4.2.2).
Theorem 4.9 (Minimum number of complexes). Let
 $G$
be a bimolecular reaction network with at least 3 species. If there exists a vector of positive rate constants
$G$
be a bimolecular reaction network with at least 3 species. If there exists a vector of positive rate constants
 $\kappa ^*$
such that
$\kappa ^*$
such that
 $(G,\kappa ^*)$
has ACR and is nondegenerately multistationary, then:
$(G,\kappa ^*)$
has ACR and is nondegenerately multistationary, then:
-
1.
$G$
has at least three reactant complexes (and hence, at least three reactions), and
-
2.
$G$
has at least five complexes (reactant and product complexes).


Proof. We first prove part (1). Let
 $(G,\kappa ^*)$
be as in the statement of the theorem and let
$(G,\kappa ^*)$
be as in the statement of the theorem and let
 $n$
denote the number of species, where
$n$
denote the number of species, where
 $n \geq 3$
. By relabelling species, if needed, we may assume that
$n \geq 3$
. By relabelling species, if needed, we may assume that
 $(G,\kappa ^*)$
has ACR in species
$(G,\kappa ^*)$
has ACR in species
 $X_1$
. Let
$X_1$
. Let
 $f_1,f_2,\dots, f_n$
denote the right-hand sides of the mass-action ODEs of
$f_1,f_2,\dots, f_n$
denote the right-hand sides of the mass-action ODEs of
 $(G,\kappa ^*)$
. As
$(G,\kappa ^*)$
. As
 $(G,\kappa ^*)$
has ACR, we know that at least one of the right-hand sides is nonzero. Let
$(G,\kappa ^*)$
has ACR, we know that at least one of the right-hand sides is nonzero. Let
 $f_i$
denote one of these nonzero polynomials.
$f_i$
denote one of these nonzero polynomials.
Assume for contradiction that
 $G$
has only
$G$
has only
 $1$
or
$1$
or
 $2$
reactant complexes. Since
$2$
reactant complexes. Since
 $G$
admits a nondegenerate positive steady state, Proposition 3.5 implies that
$G$
admits a nondegenerate positive steady state, Proposition 3.5 implies that
 $G$
is not full-dimensional and
$G$
is not full-dimensional and
 $G$
has exactly
$G$
has exactly
 $2$
reactant complexes.
$2$
reactant complexes.
We claim that all the right-hand sides
 $f_{\ell }$
are scalar multiples of each other. More precisely, we claim that for all
$f_{\ell }$
are scalar multiples of each other. More precisely, we claim that for all
 $j \in \{1,2,\dots,n\} \smallsetminus \{i\}$
, there exists
$j \in \{1,2,\dots,n\} \smallsetminus \{i\}$
, there exists
 $c_j \in \mathbb{R}$
such that
$c_j \in \mathbb{R}$
such that
 $f_j=c_j f_i$
. Indeed, each
$f_j=c_j f_i$
. Indeed, each
 $f_j$
has at most two monomials (because
$f_j$
has at most two monomials (because
 $G$
has exactly two reactant complexes), so if
$G$
has exactly two reactant complexes), so if
 $f_j$
is not a constant multiple of
$f_j$
is not a constant multiple of
 $f_i$
, then some
$f_i$
, then some
 $\mathbb{R}$
-linear combination of
$\mathbb{R}$
-linear combination of
 $f_i$
and
$f_i$
and
 $f_j$
is a monomial, and hence,
$f_j$
is a monomial, and hence,
 $(G,\kappa ^*)$
has no positive steady state (which is a contradiction).
$(G,\kappa ^*)$
has no positive steady state (which is a contradiction).
Thus, the positive steady states of
 $(G,\kappa ^*)$
are precisely the positive roots of
$(G,\kappa ^*)$
are precisely the positive roots of
 $f_i=0$
and the linear equations given by the conservation laws. Since
$f_i=0$
and the linear equations given by the conservation laws. Since
 $X_1$
is the ACR species and
$X_1$
is the ACR species and
 $G$
is bimolecular, we must have
$G$
is bimolecular, we must have
 $f_i = (\alpha - x_1) (\beta _0 + \beta _1 x_1 + \dots + \beta _n x_n)$
, where
$f_i = (\alpha - x_1) (\beta _0 + \beta _1 x_1 + \dots + \beta _n x_n)$
, where
 $\alpha$
is the (positive) ACR value and
$\alpha$
is the (positive) ACR value and
 $\beta _j \in \mathbb{R}$
for all
$\beta _j \in \mathbb{R}$
for all
 $j=0,1,\dots,n$
.
$j=0,1,\dots,n$
.
We consider several cases, based on how many of the coefficients
 $\beta _2,\beta _3, \dots, \beta _n$
are nonzero. We begin by considering the case when
$\beta _2,\beta _3, \dots, \beta _n$
are nonzero. We begin by considering the case when
 $\beta _2=\beta _3=\dots = \beta _n=0$
. In this case,
$\beta _2=\beta _3=\dots = \beta _n=0$
. In this case,
 $f_i$
is a (nonzero) polynomial in
$f_i$
is a (nonzero) polynomial in
 $x_1$
only, and so has the form
$x_1$
only, and so has the form
 $f_i=\gamma _1 x_1^{m_1} + \gamma _2 x_1^{m_2}$
, where
$f_i=\gamma _1 x_1^{m_1} + \gamma _2 x_1^{m_2}$
, where
 $\gamma _1,\gamma _2 \in \mathbb{R}$
and
$\gamma _1,\gamma _2 \in \mathbb{R}$
and
 $0 \leq m_1 \lt m_2 \leq 2$
. As
$0 \leq m_1 \lt m_2 \leq 2$
. As
 $(G,\kappa ^*)$
has a positive steady state, we conclude that
$(G,\kappa ^*)$
has a positive steady state, we conclude that
 $\gamma _1$
and
$\gamma _1$
and
 $\gamma _2$
are nonzero and have opposite signs. Now Lemma 2.3 implies that
$\gamma _2$
are nonzero and have opposite signs. Now Lemma 2.3 implies that
 $i=1$
(so,
$i=1$
(so,
 $f_1=f_i \neq 0$
) and
$f_1=f_i \neq 0$
) and
 $f_2=f_3=\dots =f_n=0$
. In fact, Lemma 2.3 implies that
$f_2=f_3=\dots =f_n=0$
. In fact, Lemma 2.3 implies that
 $X_2, \dots, X_n$
are catalyst-only species of
$X_2, \dots, X_n$
are catalyst-only species of
 $G$
(equivalently, the mass-action ODE right-hand sides for
$G$
(equivalently, the mass-action ODE right-hand sides for
 $X_2,\dots, X_n$
are zero for all choices of positive rate constants). Such a system is not multistatationary, which is a contradiction.
$X_2,\dots, X_n$
are zero for all choices of positive rate constants). Such a system is not multistatationary, which is a contradiction.
Now consider the case when two or more of the
 $\beta _2,\beta _3, \dots, \beta _n$
are nonzero. In this case, there exist distinct
$\beta _2,\beta _3, \dots, \beta _n$
are nonzero. In this case, there exist distinct
 $ j_1,j_2$
(where
$ j_1,j_2$
(where
 $2 \leq j_1,j_2 \leq n$
) with
$2 \leq j_1,j_2 \leq n$
) with
 $\beta _{j_1},\beta _{j_2}\neq 0$
. Then
$\beta _{j_1},\beta _{j_2}\neq 0$
. Then
 $f_i$
contains the monomials
$f_i$
contains the monomials
 $x_{j_1},\,x_{j_2},\,x_1x_{j_1},\,x_1x_{j_2}$
which contradicts the fact that
$x_{j_1},\,x_{j_2},\,x_1x_{j_1},\,x_1x_{j_2}$
which contradicts the fact that
 $G$
has exactly two reactant complexes.
$G$
has exactly two reactant complexes.
The final case is when exactly one of the
 $\beta _2,\beta _3, \dots, \beta _n$
is nonzero. Relabel the species, if needed, so that
$\beta _2,\beta _3, \dots, \beta _n$
is nonzero. Relabel the species, if needed, so that
 $\beta _2\neq 0$
. In this case, the two reactant complexes of
$\beta _2\neq 0$
. In this case, the two reactant complexes of
 $G$
involve only species
$G$
involve only species
 $X_1$
and
$X_1$
and
 $X_2$
. By using Lemma 2.3 again, much like we did for the prior case, we conclude that
$X_2$
. By using Lemma 2.3 again, much like we did for the prior case, we conclude that
 $X_3,\dots, X_n$
are catalyst-only species of
$X_3,\dots, X_n$
are catalyst-only species of
 $G$
and so
$G$
and so
 $(G,\kappa ^*)$
is effectively the mass-action system of a (bimolecular) network with only two species,
$(G,\kappa ^*)$
is effectively the mass-action system of a (bimolecular) network with only two species,
 $X_1$
and
$X_1$
and
 $X_2$
. Now it follows from Theorem 4.8 that
$X_2$
. Now it follows from Theorem 4.8 that
 $(G,\kappa ^*)$
is not nondegenerately multistationarity, which contradicts our assumption. This completes part (1).
$(G,\kappa ^*)$
is not nondegenerately multistationarity, which contradicts our assumption. This completes part (1).
We prove part (2). Assume for contradiction that
 $G$
has at most four complexes. By Proposition 3.10, the dimension of the stoichiometric subspace of
$G$
has at most four complexes. By Proposition 3.10, the dimension of the stoichiometric subspace of
 $G$
must be at least
$G$
must be at least
 $2$
. So, the deficiency of
$2$
. So, the deficiency of
 $G$
satisfies
$G$
satisfies
 \begin{equation*} \delta = m - \ell - \text {dim}(S) \,\leq \, 4 - 1 - 2 = 1. \end{equation*}
\begin{equation*} \delta = m - \ell - \text {dim}(S) \,\leq \, 4 - 1 - 2 = 1. \end{equation*}
Hence, the deficiency of
 $G$
is
$G$
is
 $0$
or
$0$
or
 $1$
, and the latter requires
$1$
, and the latter requires
 $G$
to have exactly one linkage class. Now Lemmas 2.5–2.6 imply that
$G$
to have exactly one linkage class. Now Lemmas 2.5–2.6 imply that
 $G$
is not multistationary, which is a contradiction.
$G$
is not multistationary, which is a contradiction.
Theorem 4.9 gives a lower bound on the number of reactant complexes and the number of all complexes (reactants and products), and the next example shows that these bounds are tight. The example also shows the tightness of the lower bounds on the number of species and the dimension of the network (from Theorem 1.2).
Example 4.10. Consider the following bimolecular network with three species and three reactant complexes and five complexes:
 \begin{equation*} G = \big\{X_1+X_2 \xrightarrow {\kappa _1} 2X_3, \quad X_3\xrightarrow {\kappa _2} X_1, \quad 2X_3\xrightarrow {\kappa _3} 2X_2\big\}. \end{equation*}
\begin{equation*} G = \big\{X_1+X_2 \xrightarrow {\kappa _1} 2X_3, \quad X_3\xrightarrow {\kappa _2} X_1, \quad 2X_3\xrightarrow {\kappa _3} 2X_2\big\}. \end{equation*}
This network is two-dimensional, as the total amount of
 $X_1,X_2,X_3$
is conserved. For every vector of positive rate constants
$X_1,X_2,X_3$
is conserved. For every vector of positive rate constants
 $\kappa$
, the system
$\kappa$
, the system
 $(G,\kappa )$
is nondegenerately multistationary and also has ACR in
$(G,\kappa )$
is nondegenerately multistationary and also has ACR in
 $X_3$
with ACR value
$X_3$
with ACR value
 $\kappa _2/ (2\kappa _3)$
. Details are given in the proof of Proposition 4.11
, below, which pertains to a family of networks that includes the network
$\kappa _2/ (2\kappa _3)$
. Details are given in the proof of Proposition 4.11
, below, which pertains to a family of networks that includes the network
 $G$
.
$G$
.
4.2.1. Non-full-dimensional networks
The bimolecular network in Example 4.10 is the
 $n=3$
case of the networks
$n=3$
case of the networks
 $G_n$
that we introduce in the next result. These networks have the property that every reactant complex is bimolecular, but (when
$G_n$
that we introduce in the next result. These networks have the property that every reactant complex is bimolecular, but (when
 $n\geq 4$
) one of product complexes is not.
$n\geq 4$
) one of product complexes is not.
Proposition 4.11 (ACR and multistationarity for all rate constants). For all
 $n \geq 3$
, consider the following network with
$n \geq 3$
, consider the following network with
 $n$
species,
$n$
species,
 $n$
reactant complexes, and
$n$
reactant complexes, and
 $n$
reactions:
$n$
reactions:
 \begin{equation*} G_n = \left \{X_1+X_2 \xrightarrow {\kappa _1} 2X_3+\sum _{j=4}^nX_j, \, X_3\xrightarrow {\kappa _2} X_1, \,2X_3\xrightarrow {\kappa _3} 2X_2\right \} \bigcup \left \{ X_4\overset {\kappa _{4}}\to 0,\, \dots,\, X_n\overset {\kappa _{n}}\to 0 \right \}. \end{equation*}
\begin{equation*} G_n = \left \{X_1+X_2 \xrightarrow {\kappa _1} 2X_3+\sum _{j=4}^nX_j, \, X_3\xrightarrow {\kappa _2} X_1, \,2X_3\xrightarrow {\kappa _3} 2X_2\right \} \bigcup \left \{ X_4\overset {\kappa _{4}}\to 0,\, \dots,\, X_n\overset {\kappa _{n}}\to 0 \right \}. \end{equation*}
Each such network
 $G_n$
satisfies the following:
$G_n$
satisfies the following:
-
1. there is a unique (up to scaling) conservation law, which is given by
$x_1+x_2+x_3=T$
, where
$T$
represents the total concentration of species
$X_1,X_2,X_3$
; and
-
2. for every vector of positive rate constants
$\kappa \in \mathbb{R}^n_{\gt 0}$
, the system
$(G_n,\kappa )$
is nondegenerately multistationary and also has ACR in species
$X_3$
,
$X_4, \dots, X_n$
.







Proof. Fix
 $n \geq 3$
. The mass-action ODEs for
$n \geq 3$
. The mass-action ODEs for
 $G_n$
are as follows:
$G_n$
are as follows:
 \begin{align*} &\frac{dx_1}{dt} = -\kappa _1x_1x_2 + \kappa _2 x_3\\[3pt] &\frac{dx_2}{dt} = -\kappa _1x_1x_2 + 2\kappa _3 x_3^2\\[3pt] &\frac{dx_3}{dt} = 2\kappa _1x_1x_2 - \kappa _2x_3 - 2\kappa _3x_3^2\\[3pt] &\frac{dx_j}{dt} = \kappa _1 x_1 x_2 - \kappa _{j} x_j \,\quad \text{ for } j\in \{4,\ldots, n\}. \end{align*}
\begin{align*} &\frac{dx_1}{dt} = -\kappa _1x_1x_2 + \kappa _2 x_3\\[3pt] &\frac{dx_2}{dt} = -\kappa _1x_1x_2 + 2\kappa _3 x_3^2\\[3pt] &\frac{dx_3}{dt} = 2\kappa _1x_1x_2 - \kappa _2x_3 - 2\kappa _3x_3^2\\[3pt] &\frac{dx_j}{dt} = \kappa _1 x_1 x_2 - \kappa _{j} x_j \,\quad \text{ for } j\in \{4,\ldots, n\}. \end{align*}
The network
 $G_n$
has exactly one conservation law (up to scaling), and it is given by
$G_n$
has exactly one conservation law (up to scaling), and it is given by
 $x_1+x_2+x_3=T$
. Additionally, using the first two ODEs, we compute that the value of species
$x_1+x_2+x_3=T$
. Additionally, using the first two ODEs, we compute that the value of species
 $X_3$
at all positive steady states is
$X_3$
at all positive steady states is
 $\frac{\kappa _2}{2\kappa _3}$
. Next, we use this steady-state value for
$\frac{\kappa _2}{2\kappa _3}$
. Next, we use this steady-state value for
 $X_3$
, together with the first and fourth ODEs, to obtain the expression
$X_3$
, together with the first and fourth ODEs, to obtain the expression
 $\frac{\kappa _2^2}{2\kappa _3\kappa _n}$
for the steady-state value for
$\frac{\kappa _2^2}{2\kappa _3\kappa _n}$
for the steady-state value for
 $X_j$
, for
$X_j$
, for
 $j \geq 4$
. Thus, ACR in
$j \geq 4$
. Thus, ACR in
 $X_3,X_4,\dots, X_n$
will follow once we confirm the existence of positive steady states.
$X_3,X_4,\dots, X_n$
will follow once we confirm the existence of positive steady states.
Next, we investigate the steady-state values of
 $X_1$
and
$X_1$
and
 $X_2$
. Using the steady-state value of
$X_2$
. Using the steady-state value of
 $X_3$
, the conservation law, and the first ODE, we see that the steady-state values of
$X_3$
, the conservation law, and the first ODE, we see that the steady-state values of
 $X_1$
and
$X_1$
and
 $X_2$
correspond to the intersection points of the line
$X_2$
correspond to the intersection points of the line
 $x_1+x_2 + \frac{\kappa _2}{2\kappa _3} = T$
and the curve
$x_1+x_2 + \frac{\kappa _2}{2\kappa _3} = T$
and the curve
 $x_1 x_2 = \frac{\kappa _2^2}{2\kappa _1\kappa _3}$
. This is depicted qualitatively below (by [green] dashed lines and a [red] solid curve, respectively).
$x_1 x_2 = \frac{\kappa _2^2}{2\kappa _1\kappa _3}$
. This is depicted qualitatively below (by [green] dashed lines and a [red] solid curve, respectively).
It follows that, given any vector of positive rate constants
 $\kappa \in \mathbb{R}^n_{\gt 0}$
, when
$\kappa \in \mathbb{R}^n_{\gt 0}$
, when
 $T$
is sufficiently large, there are two pairs of (nondegenerate) positive steady-state values for
$T$
is sufficiently large, there are two pairs of (nondegenerate) positive steady-state values for
 $X_1$
and
$X_1$
and
 $X_2$
, and so
$X_2$
, and so
 $(G_n, \kappa )$
is nondegenerately multistationary (and thus admits a positive steady state, and so has ACR).
$(G_n, \kappa )$
is nondegenerately multistationary (and thus admits a positive steady state, and so has ACR).
4.2.2. Full-dimensional networks with three species
Consider a bimolecular network
 $G$
that has three species. We saw that if
$G$
that has three species. We saw that if
 $G$
admits ACR and nondegenerate multistationarity simultaneously, then
$G$
admits ACR and nondegenerate multistationarity simultaneously, then
 $G$
has at least three reactant complexes (Theorem 4.9). If, however,
$G$
has at least three reactant complexes (Theorem 4.9). If, however,
 $G$
is full-dimensional, then more reactants are required, as stated in the following result.
$G$
is full-dimensional, then more reactants are required, as stated in the following result.
Proposition 4.12 (Minimum number of reactants for full-dimensional three-species networks). Let
 $G$
be a full-dimensional bimolecular reaction network with exactly three species. If there exists a vector of positive rate constants
$G$
be a full-dimensional bimolecular reaction network with exactly three species. If there exists a vector of positive rate constants
 $\kappa ^*$
such that
$\kappa ^*$
such that
 $(G,\kappa ^*)$
has ACR and is nondegenerately multistationary, then
$(G,\kappa ^*)$
has ACR and is nondegenerately multistationary, then
 $G$
has at least 5 reactant complexes (and hence at least five reactions)
$G$
has at least 5 reactant complexes (and hence at least five reactions)
Proposition 4.12 is a direct consequence of Propositions 3.1(3) and 3.5, and a stronger version of this result appears in the next section (Theorem 5.1). Proposition 4.12 implies that if a full-dimensional bimolecular network with three species and fewer than five reactions has both ACR and multistationarity, then this coexistence happens in a degenerate way. We illustrate this situation with two examples and then characterise all such networks with exactly four reactant complexes (Proposition 4.15).
Example 4.13.
Consider the following full-dimensional network with three species, four reactions, and four-reactant complexes:
 $\{2Z \to Z,\, X+Y \to Z \to Y+Z,\, 0 \to X\}$
. When all rate constants are
$\{2Z \to Z,\, X+Y \to Z \to Y+Z,\, 0 \to X\}$
. When all rate constants are
 $1$
, the mass-action ODEs are as follows:
$1$
, the mass-action ODEs are as follows:
 \begin{align*} \frac{dx}{dt} & = 1-xy\\[3pt] \frac{dy}{dt} & = z - xy\\[3pt] \frac{dz}{dt} & = -z^2 + xy. \end{align*}
\begin{align*} \frac{dx}{dt} & = 1-xy\\[3pt] \frac{dy}{dt} & = z - xy\\[3pt] \frac{dz}{dt} & = -z^2 + xy. \end{align*}
For this system, the set of positive steady states is
 $\{ (x,y,z) \in \mathbb{R}^3_{\gt 0} \mid xy=z=1\}$
, and every positive steady state is degenerate. We conclude that this system is multistationary (but degenerately so) and has ACR in species
$\{ (x,y,z) \in \mathbb{R}^3_{\gt 0} \mid xy=z=1\}$
, and every positive steady state is degenerate. We conclude that this system is multistationary (but degenerately so) and has ACR in species
 $Z$
(with ACR value
$Z$
(with ACR value
 $1$
).
$1$
).
Example 4.14.
Consider the following network:
 $\{ X+Z\to Z,\, Y+Z \leftrightarrows Y\to 0,\, 2X\leftarrow X\to X+Y \}$
. Like the network in Example 4.13
, this network is full-dimensional and has three species, four reactions, and four-reactant complexes; however, the set of reactant complexes differs. When all rate constants are 1, the mass-action ODEs are as follows:
$\{ X+Z\to Z,\, Y+Z \leftrightarrows Y\to 0,\, 2X\leftarrow X\to X+Y \}$
. Like the network in Example 4.13
, this network is full-dimensional and has three species, four reactions, and four-reactant complexes; however, the set of reactant complexes differs. When all rate constants are 1, the mass-action ODEs are as follows:
 \begin{align*} \frac{dx}{dt} & = x-xz\\[3pt] \frac{dy}{dt} & = x - y\\[3pt] \frac{dz}{dt} & = y - yz. \end{align*}
\begin{align*} \frac{dx}{dt} & = x-xz\\[3pt] \frac{dy}{dt} & = x - y\\[3pt] \frac{dz}{dt} & = y - yz. \end{align*}
For this system, the set of positive steady states is
 $\{ (x,y,z) \in \mathbb{R}^3_{\gt 0} \mid x=y,\,z=1\}$
, and every positive steady state is degenerate. Thus, this system is (degenerately) multistationary and has ACR in species
$\{ (x,y,z) \in \mathbb{R}^3_{\gt 0} \mid x=y,\,z=1\}$
, and every positive steady state is degenerate. Thus, this system is (degenerately) multistationary and has ACR in species
 $Z$
(with ACR value
$Z$
(with ACR value
 $1$
).
$1$
).
The next result shows that Examples 4.13 and 4.14 cover all cases of three-species, four-reactant networks with ACR and (degenerate) ACR occurring together, in the sense that these two networks represent the only two possibilities for the set of reactant complexes (when a certain full-rank condition is met, which we discuss below in Remark 4.16).
Proposition 4.15 (Networks with three species and four reactants). Let
 $G$
be a full-dimensional bimolecular reaction network with exactly three species – which we call
$G$
be a full-dimensional bimolecular reaction network with exactly three species – which we call
 $X,Y,Z$
– and exactly four reactant complexes. If
$X,Y,Z$
– and exactly four reactant complexes. If
 $\kappa ^*$
is a vector of positive rate constants such that:
$\kappa ^*$
is a vector of positive rate constants such that:
-
(a)
$\operatorname{rank}(N) = 3$
, where
$N$
is the matrix for
$(G,\kappa ^*)$
as in (7), -
(b)
$(G, \kappa ^*)$
has ACR in species
$Z$
, and
-
(c)
$(G,\kappa ^*)$
is multistationary (which is degenerately so, by Proposition 4.12), then the set of reactant complexes of
$G$
is either
$\{X,\,X+Z,\,Y,\,Y+Z\}$
or
$\{0,\,X+Y,\,Z,\,2Z\}$
.









Proof. Let
 $G$
,
$G$
,
 $\kappa ^*$
, and
$\kappa ^*$
, and
 $N$
be as in the statement of the proposition. In particular,
$N$
be as in the statement of the proposition. In particular,
 $G$
has three species and four reactants, and
$G$
has three species and four reactants, and
 $(G,\kappa ^*)$
admits a positive steady state, which we denote by
$(G,\kappa ^*)$
admits a positive steady state, which we denote by
 $(x^*,y^*, \alpha )$
(so
$(x^*,y^*, \alpha )$
(so
 $\alpha$
is the ACR value of
$\alpha$
is the ACR value of
 $Z$
). Also,
$Z$
). Also,
 $N$
has rank
$N$
has rank
 $3$
and so Proposition 3.1(2) and its proof imply that steady-state equations can be “row-reduced” so that the positive steady states of
$3$
and so Proposition 3.1(2) and its proof imply that steady-state equations can be “row-reduced” so that the positive steady states of
 $(G,\kappa ^*)$
are the roots of three binomial equations of the following form:
$(G,\kappa ^*)$
are the roots of three binomial equations of the following form:
 \begin{align*} h_1 &\,:\!=\,{ m}_1 - \beta _1{ m}_4 = 0 \\[5pt] h_2 &\,:\!=\,{ m}_2 - \beta _2{ m}_4 = 0 \\[5pt] h_3 &\,:\!=\,{ m}_3 - \beta _3{ m}_4 = 0, \end{align*}
\begin{align*} h_1 &\,:\!=\,{ m}_1 - \beta _1{ m}_4 = 0 \\[5pt] h_2 &\,:\!=\,{ m}_2 - \beta _2{ m}_4 = 0 \\[5pt] h_3 &\,:\!=\,{ m}_3 - \beta _3{ m}_4 = 0, \end{align*}
where
 $\beta _j \in \mathbb{R}$
(for
$\beta _j \in \mathbb{R}$
(for
 $j=1,2,3$
) and
$j=1,2,3$
) and
 $m_i=x^{a_i} y^{b_i} z^{c_i}$
(for
$m_i=x^{a_i} y^{b_i} z^{c_i}$
(for
 $i=1,2,3,4$
) are four distinct monic monomials given by the reactant complexes. Also, each
$i=1,2,3,4$
) are four distinct monic monomials given by the reactant complexes. Also, each
 $m_i$
(for
$m_i$
(for
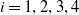 $i=1,2,3,4$
) has degree at most 2 in
$i=1,2,3,4$
) has degree at most 2 in
 $x,y,z$
(as
$x,y,z$
(as
 $G$
is bimolecular). In other words,
$G$
is bimolecular). In other words,
 $a_i,b_i,c_i$
are non-negative integers that satisfy the following:
$a_i,b_i,c_i$
are non-negative integers that satisfy the following:
 \begin{align} a_i+b_i+c_i \,\leq \, 2. \end{align}
\begin{align} a_i+b_i+c_i \,\leq \, 2. \end{align}
We infer that
 $\beta _1, \beta _2, \beta _3 \gt 0$
, because otherwise
$\beta _1, \beta _2, \beta _3 \gt 0$
, because otherwise
 $h_1=h_2=h_3=0$
would have no positive roots.
$h_1=h_2=h_3=0$
would have no positive roots.
For
 $i\in \{1,2,3\}$
, consider the following, where we recall that
$i\in \{1,2,3\}$
, consider the following, where we recall that
 $\alpha$
is the ACR value of
$\alpha$
is the ACR value of
 $Z$
:
$Z$
:
 \begin{align*} g_i \,:\!=\, h_i|_{z = \alpha } = d_i x^{a_i} y^{b_i} - d^{\prime}_i x^{a_4} y^{b_4}, \end{align*}
\begin{align*} g_i \,:\!=\, h_i|_{z = \alpha } = d_i x^{a_i} y^{b_i} - d^{\prime}_i x^{a_4} y^{b_4}, \end{align*}
where
 $d_i\,:\!=\, \alpha ^{c_i}\gt 0$
and
$d_i\,:\!=\, \alpha ^{c_i}\gt 0$
and
 $d^{\prime}_i\,:\!=\, \beta _i \alpha ^{c_4}\gt 0$
. For
$d^{\prime}_i\,:\!=\, \beta _i \alpha ^{c_4}\gt 0$
. For
 $i\in \{1,2,3\}$
, by construction,
$i\in \{1,2,3\}$
, by construction,
 $g_i(x^*,y^*)=0$
and so the subset of the positive quadrant
$g_i(x^*,y^*)=0$
and so the subset of the positive quadrant
 $\mathbb{R}^2_{\gt 0}$
defined by
$\mathbb{R}^2_{\gt 0}$
defined by
 $g_i=0$
, which we denote by
$g_i=0$
, which we denote by
 $S_i$
, is nonempty. There are four possible ‘shapes’ for each set
$S_i$
, is nonempty. There are four possible ‘shapes’ for each set
 $S_i$
:
$S_i$
:
-
1.
$S_i = \mathbb{R}^2_{\gt 0}$
, when
$(a_i,b_i)=(a_4,b_4)$
(and necessarily,
$d_i = d^{\prime}_i $
, to avoid
$S_i = \emptyset$
). -
2.
$S_i$
is the horizontal line
$y=y^*$
, when
$a_i = a_4$
and
$b_i \neq b_4$
. -
3.
$S_i$
is the vertical line
$x=x^*$
, when
$a_i \neq a_4$
and
$b_i = b_4$
. -
4.
$S_i$
is a strictly increasing curve (passing through
$(x^*,y^*)$
) defined by the following equation, when
$a_i \neq a_4$
and
$b_i \neq b_4$
:
\begin{align*} y = \left ( \frac{d_i}{d^{\prime}_i} \right ) ^{\frac{1}{b_4 - b_i } } x ^{\frac{a_i - a_4 }{b_4 - b_i } }. \end{align*}

















Any two lines/curves of the form
 $(2)$
–
$(2)$
–
 $(4)$
either coincide or intersect only at
$(4)$
either coincide or intersect only at
 $(x^*,y^*)$
. Hence, the intersection
$(x^*,y^*)$
. Hence, the intersection
 $S_1 \cap S_2 \cap S_3$
is either
$S_1 \cap S_2 \cap S_3$
is either
-
(a) the single point
$(x^*,y^*)$
, -
(b) a single line or curve of the form (2)–(4), or
-
(c) the positive quadrant
$ \mathbb{R}^2_{\gt 0}$
.


By construction and the fact that
 $\alpha$
is the ACR value, the set of all positive steady states of
$\alpha$
is the ACR value, the set of all positive steady states of
 $(G,\kappa ^*)$
is the set
$(G,\kappa ^*)$
is the set
 $\{(x,y, \alpha ) \mid (x,y) \in S_1 \cap S_2 \cap S_3\}$
. Hence, in the case of (a),
$\{(x,y, \alpha ) \mid (x,y) \in S_1 \cap S_2 \cap S_3\}$
. Hence, in the case of (a),
 $(G,\kappa ^*)$
is not multistationary, which is a contradiction.
$(G,\kappa ^*)$
is not multistationary, which is a contradiction.
Next, we show that case (c) does not occur. On the contrary, assume that it does. Then
 $S_1=S_2=S_3={\mathbb{R}}_{\gt 0}^2$
, which implies that
$S_1=S_2=S_3={\mathbb{R}}_{\gt 0}^2$
, which implies that
 $(a_1,b_1)=(a_2,b_2)=(a_3,b_3) = (a_4,b_4)$
. Since
$(a_1,b_1)=(a_2,b_2)=(a_3,b_3) = (a_4,b_4)$
. Since
 $m_1,m_2,m_3,m_4$
are four distinct monomials, it must be that
$m_1,m_2,m_3,m_4$
are four distinct monomials, it must be that
 $c_1,c_2,c_3,c_4$
are four distinct non-negative integers. However, as noted earlier,
$c_1,c_2,c_3,c_4$
are four distinct non-negative integers. However, as noted earlier,
 $c_i\in \{0,1,2\}$
for each
$c_i\in \{0,1,2\}$
for each
 $i$
, which yields a contradiction.
$i$
, which yields a contradiction.
Finally, we consider case (b). This case happens only when one of the following subcases occurs:
Subcase 1: Exactly one of the three subsets
 $S_i$
is the positive quadrant, and the other two coincide. Without loss of generality, assume
$S_i$
is the positive quadrant, and the other two coincide. Without loss of generality, assume
 $S_1 = \mathbb{R}^2_{\gt 0}$
and so
$S_1 = \mathbb{R}^2_{\gt 0}$
and so
 $S_2 = S_3 \neq \mathbb{R}^2_{\gt 0}$
. Hence,
$S_2 = S_3 \neq \mathbb{R}^2_{\gt 0}$
. Hence,
 $(a_1,b_1)=(a_4,b_4) \neq (a_2,b_2) = (a_3,b_3)$
. However,
$(a_1,b_1)=(a_4,b_4) \neq (a_2,b_2) = (a_3,b_3)$
. However,
 $m_1 \neq m_4$
and
$m_1 \neq m_4$
and
 $m_2 \neq m_3$
, and so:
$m_2 \neq m_3$
, and so:
 \begin{align} c_1\,\neq \, c_4 \quad \textrm{and} \quad c_2\,\neq \,c_3. \end{align}
\begin{align} c_1\,\neq \, c_4 \quad \textrm{and} \quad c_2\,\neq \,c_3. \end{align}
We rewrite the inequalities (39), using the equalities
 $(a_1,b_1)=(a_4,b_4)$
and
$(a_1,b_1)=(a_4,b_4)$
and
 $(a_2,b_2) = (a_3,b_3)$
:
$(a_2,b_2) = (a_3,b_3)$
:
 \begin{align} a_1+b_1+c_1 \,\leq \, 2, \quad a_1+b_1+c_4 \,\leq \, 2, \quad a_2+b_2+c_2 \,\leq \, 2, \quad a_2+b_2+c_3 \,\leq \, 2. \end{align}
\begin{align} a_1+b_1+c_1 \,\leq \, 2, \quad a_1+b_1+c_4 \,\leq \, 2, \quad a_2+b_2+c_2 \,\leq \, 2, \quad a_2+b_2+c_3 \,\leq \, 2. \end{align}
Finally, Lemma 3.8 implies that each of species
 $X$
and
$X$
and
 $Y$
takes part in some reactant complex, so we obtain the following (again using
$Y$
takes part in some reactant complex, so we obtain the following (again using
 $(a_1,b_1)=(a_4,b_4)$
and
$(a_1,b_1)=(a_4,b_4)$
and
 $(a_2,b_2) = (a_3,b_3)$
):
$(a_2,b_2) = (a_3,b_3)$
):
 \begin{align} a_1 + a_2 \,\geq \, 1 \quad \textrm{and} \quad b_1 + b_2 \,\geq \, 1. \end{align}
\begin{align} a_1 + a_2 \,\geq \, 1 \quad \textrm{and} \quad b_1 + b_2 \,\geq \, 1. \end{align}
The only non-negative solutions to the conditions in (40), (41), and (42) are as follows:
-
$a_1=a_3=1$
,
$a_2=a_3=0$
,
$b_1=b_4=0$
,
$b_2=b_3=1$
,
$\{c_1,c_4\} = \{c_2,c_3\} = \{0,1\}$
;
-
$a_1=a_3=0$
,
$a_2=a_3=1$
,
$b_1=b_4=1$
,
$b_2=b_3=0$
,
$\{c_1,c_4\} = \{c_2,c_3\} = \{0,1\}$
.










In all of these solutions, the set of reactant complexes is
 $\{X, \,X+Z, \,Y, \,Y+Z\}$
.
$\{X, \,X+Z, \,Y, \,Y+Z\}$
.
Subcase 2: Exactly two of the three subsets
 $S_i$
are the positive quadrant. Without loss of generality, assume that
$S_i$
are the positive quadrant. Without loss of generality, assume that
 $S_1 = S_2 = \mathbb{R}^2_{\gt 0} \neq S_3$
. This implies the following:
$S_1 = S_2 = \mathbb{R}^2_{\gt 0} \neq S_3$
. This implies the following:
 \begin{align} (a_1,b_1) = (a_2,b_2) = (a_4,b_4) \,\neq \, (a_3,b_3). \end{align}
\begin{align} (a_1,b_1) = (a_2,b_2) = (a_4,b_4) \,\neq \, (a_3,b_3). \end{align}
However,
 $m_1,m_2,m_4$
are three distinct monomials, so
$m_1,m_2,m_4$
are three distinct monomials, so
 $c_1, c_2,c_4$
are three distinct non-negative integers. Now inequality (39) implies that
$c_1, c_2,c_4$
are three distinct non-negative integers. Now inequality (39) implies that
 $\{c_1, c_2,c_4\} = \{0,1,2\}$
. Let
$\{c_1, c_2,c_4\} = \{0,1,2\}$
. Let
 $i^* \in \{1,2,4\}$
be such that
$i^* \in \{1,2,4\}$
be such that
 $c_{i^*}=2$
. Next, the equalities in (43) and the inequality (39) for
$c_{i^*}=2$
. Next, the equalities in (43) and the inequality (39) for
 $i=i^*$
together imply that
$i=i^*$
together imply that
 $(a_1,b_1)=(a_2,b_2)=(a_4,b_4)=(0,0)$
. Therefore, the set of reactant complexes corresponding to
$(a_1,b_1)=(a_2,b_2)=(a_4,b_4)=(0,0)$
. Therefore, the set of reactant complexes corresponding to
 $m_1,m_2,m_4$
is
$m_1,m_2,m_4$
is
 $\{0, \,Z, \,2Z\}$
. Finally, Lemma 3.8 implies that the fourth reactant complex must involve both
$\{0, \,Z, \,2Z\}$
. Finally, Lemma 3.8 implies that the fourth reactant complex must involve both
 $X$
and
$X$
and
 $Y$
and so (by bimolecularity) is
$Y$
and so (by bimolecularity) is
 $X+Y$
. Therefore, the set of reactant complexes is
$X+Y$
. Therefore, the set of reactant complexes is
 $\{0,\, X+Y, \,Z, \,2Z\}$
.
$\{0,\, X+Y, \,Z, \,2Z\}$
.
Subcase 3: None of the subsets
 $S_i$
are positive quadrants, and the three sets coincide. This implies that
$S_i$
are positive quadrants, and the three sets coincide. This implies that
 $(a_1,b_1)=(a_2,b_2)=(a_3,b_3) \neq (a_4,b_4)$
. These conditions are symmetric to those in subcase 2, and so the reactant complexes are
$(a_1,b_1)=(a_2,b_2)=(a_3,b_3) \neq (a_4,b_4)$
. These conditions are symmetric to those in subcase 2, and so the reactant complexes are
 $\{0,\, Z, \,2Z, \,X+Y\}.$
This completes subcase 3 (and case (b)).
$\{0,\, Z, \,2Z, \,X+Y\}.$
This completes subcase 3 (and case (b)).
Remark 4.16 (Rank condition). Proposition 4.15 includes the hypothesis that the matrix
 $N$
for the system
$N$
for the system
 $(G,\kappa ^*)$
has (full) rank
$(G,\kappa ^*)$
has (full) rank
 $3$
. If we remove this hypothesis, we can obtain more full-dimensional networks with three species and four reactants that allow ACR and (degenerate) multistationarity to occur together. We present one such network in Example 4.17.
$3$
. If we remove this hypothesis, we can obtain more full-dimensional networks with three species and four reactants that allow ACR and (degenerate) multistationarity to occur together. We present one such network in Example 4.17.
Example 4.17. Consider the following full-dimensional network with three species and four reactants:
 \begin{equation*}G\,:\!=\,\{0\to X\to Y \to 2 Y,\,\, Y\leftarrow Y+Z \to 2 Z\}. \end{equation*}
\begin{equation*}G\,:\!=\,\{0\to X\to Y \to 2 Y,\,\, Y\leftarrow Y+Z \to 2 Z\}. \end{equation*}
The system
 $(G,\kappa ^*)$
obtained by setting all the reaction rates to 1 has the following ODEs:
$(G,\kappa ^*)$
obtained by setting all the reaction rates to 1 has the following ODEs:
 \begin{align*} \begin{bmatrix} dx/dt\\[5pt] dy/dt\\[5pt] dz/dt \end{bmatrix} = \begin{bmatrix} -1& \quad 0 & \quad 0 & \quad 1 \\[5pt] 1 & \quad 1 & \quad -1 & \quad 0 \\[5pt] 0& \quad 0 & \quad 0 & \quad 0 \\[5pt] \end{bmatrix} \begin{bmatrix} x\\[5pt] y\\[5pt] yz\\[5pt] 1 \end{bmatrix} = N \begin{bmatrix} x\\[5pt] y\\[5pt] yz\\[5pt] 1 \end{bmatrix}. \end{align*}
\begin{align*} \begin{bmatrix} dx/dt\\[5pt] dy/dt\\[5pt] dz/dt \end{bmatrix} = \begin{bmatrix} -1& \quad 0 & \quad 0 & \quad 1 \\[5pt] 1 & \quad 1 & \quad -1 & \quad 0 \\[5pt] 0& \quad 0 & \quad 0 & \quad 0 \\[5pt] \end{bmatrix} \begin{bmatrix} x\\[5pt] y\\[5pt] yz\\[5pt] 1 \end{bmatrix} = N \begin{bmatrix} x\\[5pt] y\\[5pt] yz\\[5pt] 1 \end{bmatrix}. \end{align*}
The matrix
 $N$
(defined above) has rank
$N$
(defined above) has rank
 $2$
, the set of positive steady states is
$2$
, the set of positive steady states is
 $\{ (x,y,z) \in \mathbb{R}^3_{\gt 0} \mid x=1,$
$\{ (x,y,z) \in \mathbb{R}^3_{\gt 0} \mid x=1,$
 $y(z-1)=1\}$
, and every positive steady state is degenerate. Thus, this system is (degenerately) multistationary and has ACR in species
$y(z-1)=1\}$
, and every positive steady state is degenerate. Thus, this system is (degenerately) multistationary and has ACR in species
 $X$
(with ACR value
$X$
(with ACR value
 $1$
).
$1$
).
In the next section, we see that the exceptional networks in Proposition 4.15 – namely, full-dimensional, three-species networks with reactant complex set
 $ \{0,Z,2Z,X+Y\}$
or
$ \{0,Z,2Z,X+Y\}$
or
 $\{X,Y,X+Z,Y+Z\}$
– do not have unconditional ACR. Indeed, this fact is a direct consequence of a more general result concerning networks with
$\{X,Y,X+Z,Y+Z\}$
– do not have unconditional ACR. Indeed, this fact is a direct consequence of a more general result concerning networks with
 $n$
species and
$n$
species and
 $n+1$
reactants (Theorem 5.4).
$n+1$
reactants (Theorem 5.4).
5. Main results on general networks
The results in the prior section pertain to networks that are bimolecular, while here we analyse networks that need not be bimolecular. We consider full-dimensional networks (Section 5.1) and non-full-dimensional networks (Section 5.2) separately.
5.1. Full-dimensional networks
In Proposition 4.11, we saw a family of networks that admit ACR and nondegenerate multistationarity together. These networks have
 $n$
reactants (where
$n$
reactants (where
 $n$
is the number of species), but are not full-dimensional. In this subsection, we show that for full-dimensional networks, the coexistence of ACR and nondegenerate multistationarity requires at least
$n$
is the number of species), but are not full-dimensional. In this subsection, we show that for full-dimensional networks, the coexistence of ACR and nondegenerate multistationarity requires at least
 $n+2$
reactants (Theorem 5.1). We also show that this lower bound is tight (Proposition 5.3). Additionally, we consider full-dimensional networks with only
$n+2$
reactants (Theorem 5.1). We also show that this lower bound is tight (Proposition 5.3). Additionally, we consider full-dimensional networks with only
 $n+1$
reactants and show that if such a network is multistationary (even if only degenerately so), then the network cannot have unconditional ACR (Theorem 5.4).
$n+1$
reactants and show that if such a network is multistationary (even if only degenerately so), then the network cannot have unconditional ACR (Theorem 5.4).
Theorem 5.1 (Minimum number of reactants for full-dimensional networks). Let
 $G$
be a full-dimensional reaction network with
$G$
be a full-dimensional reaction network with
 $n$
species. If there exists a vector of positive rate constants
$n$
species. If there exists a vector of positive rate constants
 $\kappa ^*$
such that the mass-action system
$\kappa ^*$
such that the mass-action system
 $(G,\kappa ^*)$
has ACR and also is nondegenerately multistationary, then
$(G,\kappa ^*)$
has ACR and also is nondegenerately multistationary, then
 $n \geq 2$
and
$n \geq 2$
and
 $G$
has at least
$G$
has at least
 $n+2$
reactant complexes and hence, at least
$n+2$
reactant complexes and hence, at least
 $n+2$
reactions.
$n+2$
reactions.
Proof. It follows readily from definitions that ACR and multistationarity do not coexist in networks with only one species, so
 $n \geq 2$
. We proceed by contrapositive. We consider two cases. If
$n \geq 2$
. We proceed by contrapositive. We consider two cases. If
 $G$
has at most
$G$
has at most
 $n$
reactant complexes, then Proposition 3.5 (which requires
$n$
reactant complexes, then Proposition 3.5 (which requires
 $n \geq 2$
) implies that every positive steady state of
$n \geq 2$
) implies that every positive steady state of
 $(G,\kappa ^*)$
is degenerate and so
$(G,\kappa ^*)$
is degenerate and so
 $(G,\kappa ^*)$
is not nondegenerately multistationary. In the remaining case, when
$(G,\kappa ^*)$
is not nondegenerately multistationary. In the remaining case, when
 $G$
has
$G$
has
 $n+1$
reactant complexes, Proposition 3.1(3) implies that
$n+1$
reactant complexes, Proposition 3.1(3) implies that
 $(G,\kappa ^*)$
is not nondegenerately multistationary.
$(G,\kappa ^*)$
is not nondegenerately multistationary.
The next example shows that the bound in Theorem 5.1 is tight for
 $n=2$
.
$n=2$
.
Example 5.2. The following network is full-dimensional and has two species, four-reactant complexes, and four reactions (the out-of-order labelling of the rate constants is to be consistent with Proposition 5.3 , which appears later):
 \begin{equation*} \left\{ A+B \overset {\kappa _1 } \to 2B \overset {\kappa _3 } \to 2B+A, \, B \overset {\kappa _2 } \to 0 \overset {\kappa _4 } \to A \right\}. \end{equation*}
\begin{equation*} \left\{ A+B \overset {\kappa _1 } \to 2B \overset {\kappa _3 } \to 2B+A, \, B \overset {\kappa _2 } \to 0 \overset {\kappa _4 } \to A \right\}. \end{equation*}
Observe that all reactant complexes are bimolecular, but one of the product complexes is not. In the next result, we show that this network exhibits ACR (in species
 $A$
with ACR value
$A$
with ACR value
 $\kappa _2/\kappa _1$
) and nondegenerate multistationarity when
$\kappa _2/\kappa _1$
) and nondegenerate multistationarity when
 $\kappa _2^2 \gt 4 \kappa _3 \kappa _4.$
(Proposition 5.3). Among full-dimensional networks for which ACR and nondegenerate multistationarity coexist, this network is optimal in the sense that it has the fewest possible species, reactant complexes, and reactions (by Theorem
5.1
).
$\kappa _2^2 \gt 4 \kappa _3 \kappa _4.$
(Proposition 5.3). Among full-dimensional networks for which ACR and nondegenerate multistationarity coexist, this network is optimal in the sense that it has the fewest possible species, reactant complexes, and reactions (by Theorem
5.1
).
In the next result, we generalise the network in Example 5.2 to a family of networks that show that the lower bound on the number of reactions in Theorem 5.1 is tight for all
 $n$
. The networks in the following result are also optimal in terms of the molecularity of the reactant complexes (they are bimolecular), although two of the product complexes have high molecularity.
$n$
. The networks in the following result are also optimal in terms of the molecularity of the reactant complexes (they are bimolecular), although two of the product complexes have high molecularity.
Proposition 5.3.
For all
 $n \geq 2$
, consider the following full-dimensional network with
$n \geq 2$
, consider the following full-dimensional network with
 $n$
species,
$n$
species,
 $n+2$
reactions, and
$n+2$
reactions, and
 $n+2$
reactant complexes:
$n+2$
reactant complexes:
 \begin{align*} G_n = \left \{ X_1+X_2 \overset{\kappa _1 } \to 2X_2 + X_3 + \dots + X_n,\, X_2 \overset{\kappa _2 } \to 0,\, 2X_2 \overset{\kappa _3 } \to 2X_2+X_1,\, 0 \overset{\kappa _4 } \to X_1 \right \} \bigcup \left\{ X_j\overset{\kappa _{j+2}}\to 0 \mid 3 \leq j \leq n\right\}. \end{align*}
\begin{align*} G_n = \left \{ X_1+X_2 \overset{\kappa _1 } \to 2X_2 + X_3 + \dots + X_n,\, X_2 \overset{\kappa _2 } \to 0,\, 2X_2 \overset{\kappa _3 } \to 2X_2+X_1,\, 0 \overset{\kappa _4 } \to X_1 \right \} \bigcup \left\{ X_j\overset{\kappa _{j+2}}\to 0 \mid 3 \leq j \leq n\right\}. \end{align*}
For every vector of positive rate constants
 $\kappa ^*$
for which
$\kappa ^*$
for which
 $(\kappa ^*_2)^2 \gt 4 \kappa ^*_3 \kappa ^*_4$
, the system
$(\kappa ^*_2)^2 \gt 4 \kappa ^*_3 \kappa ^*_4$
, the system
 $(G_n, \kappa ^*)$
has nondegenerate multistationarity and has ACR in species
$(G_n, \kappa ^*)$
has nondegenerate multistationarity and has ACR in species
 $X_1$
.
$X_1$
.
Proof. The mass-action ODEs are given by:
 \begin{align*}{\frac{dx_1}{dt}} & = \kappa _3 x_2^2 - \kappa _1 x_1 x_2 + \kappa _4\\[5pt]{\frac{dx_2}{dt}} & = \kappa _1 x_1 x_2-\kappa _2x_2\\[5pt]{\frac{dx_j}{dt}} & = \kappa _1 x_1 x_2 - \kappa _{j+2} x_j \,\quad \text{ for } j\in{3,\ldots, n.} \end{align*}
\begin{align*}{\frac{dx_1}{dt}} & = \kappa _3 x_2^2 - \kappa _1 x_1 x_2 + \kappa _4\\[5pt]{\frac{dx_2}{dt}} & = \kappa _1 x_1 x_2-\kappa _2x_2\\[5pt]{\frac{dx_j}{dt}} & = \kappa _1 x_1 x_2 - \kappa _{j+2} x_j \,\quad \text{ for } j\in{3,\ldots, n.} \end{align*}
The steady-state equation for
 $X_2$
implies that
$X_2$
implies that
 $x_1=\kappa _2/\kappa _1$
at all positive steady states, so there is ACR in
$x_1=\kappa _2/\kappa _1$
at all positive steady states, so there is ACR in
 $X_1$
(whenever positive steady states exist). Next, the steady-state equations for
$X_1$
(whenever positive steady states exist). Next, the steady-state equations for
 $X_1$
and
$X_1$
and
 $X_2$
imply that the steady-state values of
$X_2$
imply that the steady-state values of
 $X_2$
are
$X_2$
are
 $x_2^{\pm } \,:\!=\, \frac{k_2 \pm \sqrt{k_2^2 - 4k_3k_4}}{2 k_3}$
. Both of these steady-state values are positive precisely when the discriminant
$x_2^{\pm } \,:\!=\, \frac{k_2 \pm \sqrt{k_2^2 - 4k_3k_4}}{2 k_3}$
. Both of these steady-state values are positive precisely when the discriminant
 $k_2^2 - 4k_3k_4$
is positive (this is a straightforward computation; alternatively, see [Reference Dennis and Shiu9, Proposition 2.3]). Now we use the steady-state equation for
$k_2^2 - 4k_3k_4$
is positive (this is a straightforward computation; alternatively, see [Reference Dennis and Shiu9, Proposition 2.3]). Now we use the steady-state equation for
 $X_j$
, where
$X_j$
, where
 $j\geq 3$
, to compute the two positive steady states that exist whenever
$j\geq 3$
, to compute the two positive steady states that exist whenever
 $(\kappa ^*_2)^2 \gt 4 \kappa ^*_3 \kappa ^*_4$
:
$(\kappa ^*_2)^2 \gt 4 \kappa ^*_3 \kappa ^*_4$
:
 \begin{align*} & \left ( x_1^*,\, x_2^{+},\, \frac{\kappa _1}{\kappa _3} x_1^*x_2^{+},\, \dots, \, \frac{\kappa _1}{\kappa _n} x_1^*x_2^{+} \right ) \quad \textrm{and } \quad \left ( x_1^*,\, x_2^{-},\, \frac{\kappa _1}{\kappa _3} x_1^*x_2^{-},\, \dots, \, \frac{\kappa _1}{\kappa _n} x_1^*x_2^{-} \right ), \end{align*}
\begin{align*} & \left ( x_1^*,\, x_2^{+},\, \frac{\kappa _1}{\kappa _3} x_1^*x_2^{+},\, \dots, \, \frac{\kappa _1}{\kappa _n} x_1^*x_2^{+} \right ) \quad \textrm{and } \quad \left ( x_1^*,\, x_2^{-},\, \frac{\kappa _1}{\kappa _3} x_1^*x_2^{-},\, \dots, \, \frac{\kappa _1}{\kappa _n} x_1^*x_2^{-} \right ), \end{align*}
where
 $x^*_1\,:\!=\,\kappa _2/\kappa _1$
. Finally, nondegeneracy can be checked by computing the Jacobian matrix.
$x^*_1\,:\!=\,\kappa _2/\kappa _1$
. Finally, nondegeneracy can be checked by computing the Jacobian matrix.
Our next result concerns full-dimensional networks with one more reactant than the number of species, as follows.
Theorem 5.4 (Networks with
 $n+1$
reactants). Let
$n+1$
reactants). Let
 $G$
be a full-dimensional network, with
$G$
be a full-dimensional network, with
 $n$
species and exactly
$n$
species and exactly
 $n+1$
reactant complexes. If
$n+1$
reactant complexes. If
 $G$
is multistationary, then there exists a vector of positive rate constants
$G$
is multistationary, then there exists a vector of positive rate constants
 $\kappa$
such that
$\kappa$
such that
 $(G,\kappa )$
has no positive steady states, and hence
$(G,\kappa )$
has no positive steady states, and hence
 $G$
does not have unconditional ACR.
$G$
does not have unconditional ACR.
Proof. Assume that
 $G$
is full-dimensional, has exactly
$G$
is full-dimensional, has exactly
 $n+1$
reactant complexes (where
$n+1$
reactant complexes (where
 $n$
is the number of species), and is multistationarity. By definition, there exists
$n$
is the number of species), and is multistationarity. By definition, there exists
 $\kappa ^* \in \mathbb{R}^r_{\gt 0}$
, where
$\kappa ^* \in \mathbb{R}^r_{\gt 0}$
, where
 $r$
is the number of reactions, such that
$r$
is the number of reactions, such that
 $(G,\kappa ^*)$
is multistationary. Let
$(G,\kappa ^*)$
is multistationary. Let
 $N$
be the
$N$
be the
 $n \times (n+1)$
matrix arising from
$n \times (n+1)$
matrix arising from
 $(G,\kappa ^*)$
, as in (7); and let
$(G,\kappa ^*)$
, as in (7); and let
 $A$
be the
$A$
be the
 $n \times n$
matrix defined by
$n \times n$
matrix defined by
 $G$
, as in Proposition 3.7.
$G$
, as in Proposition 3.7.
We claim that
 $\operatorname{rank}(N) \leq n-1$
or
$\operatorname{rank}(N) \leq n-1$
or
 $\operatorname{rank}(A) \leq n-1$
. Indeed, if
$\operatorname{rank}(A) \leq n-1$
. Indeed, if
 $\operatorname{rank}(N) = n$
and
$\operatorname{rank}(N) = n$
and
 $\operatorname{rank}(A) = n$
, then the proof of Proposition 3.7 shows that
$\operatorname{rank}(A) = n$
, then the proof of Proposition 3.7 shows that
 $(G,\kappa ^*)$
is not multistationary, which is a contradiction.
$(G,\kappa ^*)$
is not multistationary, which is a contradiction.
If
 $\operatorname{rank}(N) \leq n-1$
, then Proposition 3.6(2) implies that there exists
$\operatorname{rank}(N) \leq n-1$
, then Proposition 3.6(2) implies that there exists
 $\kappa ^{**} \in \mathbb{R}^r_{\gt 0}$
such that
$\kappa ^{**} \in \mathbb{R}^r_{\gt 0}$
such that
 $(G,\kappa ^{**})$
has no positive steady states. Similarly, in the remaining case, when
$(G,\kappa ^{**})$
has no positive steady states. Similarly, in the remaining case, when
 $\operatorname{rank}(N) =n$
and
$\operatorname{rank}(N) =n$
and
 $\operatorname{rank}(A) \leq n-1$
, the desired result follows directly from Proposition 3.7(2).
$\operatorname{rank}(A) \leq n-1$
, the desired result follows directly from Proposition 3.7(2).
5.2. Non-full-dimensional networks
In an earlier section, we saw a family of networks with
 $n$
species,
$n$
species,
 $n$
reactant complexes, and exactly one conservation law, for which ACR and nondegenerate multistationarity coexist (Proposition 4.11). Our next result shows that this
$n$
reactant complexes, and exactly one conservation law, for which ACR and nondegenerate multistationarity coexist (Proposition 4.11). Our next result shows that this
 $n$
is the minimum number of reactant complexes (when there is one conservation law), and, furthermore, as the number of conservation laws increases, the minimum number of reactant complexes required decreases.
$n$
is the minimum number of reactant complexes (when there is one conservation law), and, furthermore, as the number of conservation laws increases, the minimum number of reactant complexes required decreases.
Theorem 5.5 (Minimum number of reactants). Let
 $G$
be a reaction network with
$G$
be a reaction network with
 $n \geq 3$
species and
$n \geq 3$
species and
 $k \geq 1$
conservation laws (more precisely,
$k \geq 1$
conservation laws (more precisely,
 $G$
has dimension
$G$
has dimension
 $n-k$
). If there exists a vector of positive rate constants
$n-k$
). If there exists a vector of positive rate constants
 $\kappa ^*$
such that the system
$\kappa ^*$
such that the system
 $(G,\kappa ^*)$
is nondegenerately multistationary and has ACR in some species, then
$(G,\kappa ^*)$
is nondegenerately multistationary and has ACR in some species, then
 $G$
has at least
$G$
has at least
 $n-k+1$
reactant complexes.
$n-k+1$
reactant complexes.
Proof. If
 $G$
has
$G$
has
 $k \geq 1$
conservation laws and at most
$k \geq 1$
conservation laws and at most
 $n-k$
reactant complexes, then Propositions 3.5(1) and 3.9 together imply that
$n-k$
reactant complexes, then Propositions 3.5(1) and 3.9 together imply that
 $G$
is not nondegenerate multistationarity.
$G$
is not nondegenerate multistationarity.
As noted earlier, the bound in Theorem 5.5 is tight for
 $k=1$
, due to Proposition 4.11. We also know that, for
$k=1$
, due to Proposition 4.11. We also know that, for
 $k=n-1$
, the bound holds vacuously (Proposition 3.10). Our next result shows that the bound is also tight for all remaining values of
$k=n-1$
, the bound holds vacuously (Proposition 3.10). Our next result shows that the bound is also tight for all remaining values of
 $k$
(namely,
$k$
(namely,
 $ 2 \leq k \leq n-2$
).
$ 2 \leq k \leq n-2$
).
Proposition 5.6.
Let
 $n \geq 3$
, and let
$n \geq 3$
, and let
 $k \in \{2,3,\dots, n-2\}$
. If
$k \in \{2,3,\dots, n-2\}$
. If
 $k \neq n-2$
, consider the following network:
$k \neq n-2$
, consider the following network:
 \begin{align*} G_{n,k} = &\left \{X_1+X_2+\sum _{j=n+2-k}^{n}X_j \xrightarrow{\kappa _1} 2X_3+\sum _{j=4}^nX_j, \, X_3\xrightarrow{\kappa _2} X_1, \,2X_3\xrightarrow{\kappa _3} 2X_2\right \}\\[9pt] &\bigcup \left \{ X_{4}\overset{\kappa _{4}}\to 0,\, \dots,\, X_{n+1-k}\overset{\kappa _{n-k+1}}\to 0 \right \}. \end{align*}
\begin{align*} G_{n,k} = &\left \{X_1+X_2+\sum _{j=n+2-k}^{n}X_j \xrightarrow{\kappa _1} 2X_3+\sum _{j=4}^nX_j, \, X_3\xrightarrow{\kappa _2} X_1, \,2X_3\xrightarrow{\kappa _3} 2X_2\right \}\\[9pt] &\bigcup \left \{ X_{4}\overset{\kappa _{4}}\to 0,\, \dots,\, X_{n+1-k}\overset{\kappa _{n-k+1}}\to 0 \right \}. \end{align*}
On the other hand, if
 $k = n-2$
, consider the following network:
$k = n-2$
, consider the following network:
 \begin{align*} G_{n,k} = \left \{X_1+X_2+\sum _{j=4}^{n}X_j \xrightarrow{\kappa _1} 2X_3+\sum _{j=4}^nX_j, \, X_3\xrightarrow{\kappa _2} X_1, \,2X_3\xrightarrow{\kappa _3} 2X_2\right \} \end{align*}
\begin{align*} G_{n,k} = \left \{X_1+X_2+\sum _{j=4}^{n}X_j \xrightarrow{\kappa _1} 2X_3+\sum _{j=4}^nX_j, \, X_3\xrightarrow{\kappa _2} X_1, \,2X_3\xrightarrow{\kappa _3} 2X_2\right \} \end{align*}
Each such network
 $G_{n,k}$
satisfies the following:
$G_{n,k}$
satisfies the following:
-
1.
$G_{n,k}$
has
$n$
species,
$n-k+1$
reactants, and
$n-k+1$
reactions;
-
2.
$G_{n,k}$
has dimension
$n-k$
, and the following are
$k$
linearly independent conservation laws:
$x_1+x_2+x_3=T$
and
$x_j = T_j$
for
$j\in \{n-k+2,\dots,n\}$
. -
3. for every vector of positive rate constants
$\kappa$
, the system
$(G_n,\kappa )$
is nondegenerately multistationary and also has ACR in species
$X_3$
,
$X_4, \dots, X_{n-k+1}$
.














Proof. This result can be checked directly, in a manner similar to the proof of Proposition 4.11. Indeed, for every vector of positive rate constants, there is ACR in species
 $X_3,X_4, \ldots,X_{n-k+1}$
and exactly two nondegenerate positive steady states when
$X_3,X_4, \ldots,X_{n-k+1}$
and exactly two nondegenerate positive steady states when
 $T$
is large enough.
$T$
is large enough.
The reaction networks in Proposition 5.6 are not bimolecular, and they contain reactions with many catalyst-only species (namely
 $X_{n-k+2},\dots,X_n$
). We do not know whether there exist reaction networks that are bimolecular and do not contain reactions with catalyst-only species, and yet (like the networks in Proposition 5.6) show that the lower bound in Theorem 5.5 is tight.
$X_{n-k+2},\dots,X_n$
). We do not know whether there exist reaction networks that are bimolecular and do not contain reactions with catalyst-only species, and yet (like the networks in Proposition 5.6) show that the lower bound in Theorem 5.5 is tight.
6. Discussion
In this article, we proved lower bounds in terms of the dimension and the numbers of species, reactant complexes (and thus reactions), and all complexes (both reactant and product complexes) needed for the coexistence of ACR and nondegenerate multistationarity. Additionally, we showed that these bounds are tight, via the network
 $\{A+B \to 2C \to 2B,\, C\to A \}$
(Example 4.10).
$\{A+B \to 2C \to 2B,\, C\to A \}$
(Example 4.10).
Networks like the one in Example 4.10 contain special structures that may be biologically significant. Exploring such structures will aid in establishing design principles for creating networks with ACR and multistationarity. We plan to explore such networks and their architecture in the future.
In the present work, our interest in multistationarity comes from the fact that it is a necessary condition for multistability. Another interesting direction, therefore, is to investigate conditions for coexistence of ACR and multistability, rather than multistationarity. The “minimal” networks in the current work admit only two positive steady states and are not multistable. Hence, we conjecture that the lower bounds (on dimension and the numbers of species, reactant complexes, and all complexes) for the coexistence of ACR and multistability are strictly larger than the bounds proven here for ACR and multistationarity.
Finally, we are interested in the conditions for the coexistence of other combinations of biologically significant dynamical properties, such as ACR and oscillations. In addition to the minimum requirements for their coexistence, we also hope to discover new network architectures or motifs that can be used to design synthetic networks possessing these dynamical properties.
Acknowledgements
The authors thank Elisenda Feliu, Oskar Henriksson, Badal Joshi, and Beatriz Pascual-Escudero for many helpful discussions. The authors also acknowledge the referees for their helpful suggestions.
Funding statement
This project began at an AIM workshop on “Limits and control of stochastic reaction networks” held online in July 2021. AS was supported by the NSF (DMS-1752672). NK has been funded by the Independent Research Fund of Denmark.
Competing interests
None.



 Open access
Open access