


INTRODUCTION
Canine leishmaniasis (CanL) is a zoonotic parasitical infection of dogs caused by Leishmania ssp. and transmitted by infected phlebotomine sandflies [Reference Solano-Gallego1]. In dogs, CanL causes chronic infection that may progress to a clinical stage and can be fatal if untreated [Reference Solano-Gallego1]. The infection can be transmitted to humans causing visceral or cutaneous leishmaniasis, and it is the second most important protozoan infection after malaria [Reference Rodríguez-Cortés2].
In Europe, CanL is endemic in the Mediterranean basin but has been reported more frequently in northern latitudes of Europe where sandflies were previously thought to be absent or present only in very low densities [Reference Ready3]. In CanL-free areas of Europe, seropositive dogs have been associated with imported dogs or dogs returning from endemic areas [Reference Ready3–Reference Mettler5].
Clinical trials have been used to evaluate the individual effect of mitigation measures (MMs) to control CanL infection [Reference Killick-Kendrick6–Reference Coleman8]. In a simulation model, Dye [Reference Dye9] also evaluated the relative reduction in CanL incidence by varying the (unknown) efficacy of different MMs.
Compartmental deterministic models have helped in understanding CanL transmission in dogs [Reference Dye9–Reference Palatnik-De-Sousa11] and evaluating MMs in endemic areas [Reference Solano-Gallego1, Reference Dye9]. Nonetheless, stochastic models can more realistically model disease transmission and persistence in small populations such as contact networks of dogs, where the effect of randomness largely influences the variability of the results [Reference Westgarth12]. Stochastic models also facilitate the modelling of uncertainty in key parameters.
The objectives of this study were: (1) to estimate the probability of CanL persistence in a previously CanL-free area with competent vectors following the introduction of infected dogs and (2) to evaluate the effectiveness of repellent use, vaccination, prophylactic medication, insecticide use, or test and exclusion of infected animals in reducing the probability of introduction and persistence of CanL in a free area.
METHODS
The overall probability of CanL persistence in a previously CanL-free area was estimated using a model with three main simulation steps. In the first step, two disease introduction pathways were considered: imports of infected dogs from endemic areas (P infCA), and infection of dogs during travels to endemic areas with their owners (P inf). In the second step, CanL transmission in a previously free area in the presence of sandflies was simulated to estimate the probability of persistence (P per) in a hypothetical independent contact network of dogs (Fig. 1). Finally, the joint uncertainty distributions for P per, P inf and P infCA, were used in a third step to simulate the probability of persistence in at least one independent contact network in a previously CanL-free region with sandflies (P per region).

Fig. 1. Schematic representation of the model of canine leishmaniasis transmission and movement of dogs between endemic and CanL-free areas of Europe. Solid black boxes represent infection states of the dog population. Solid arrows indicate transitions between infection stages. Dashed black boxes represent the sandfly population. Dashed black arrows represent mitigation measures. Grey dashed arrow represents movement of dogs between endemic and non-endemic areas of Europe.
P inf and P per were both modelled using an individual-based stochastic model of CanL transmission in a contact network of dogs. A contact network of dogs represented a group of dogs that interact and may be able to transmit CanL to each other through sandflies (e.g. dogs in a neighbourhood, or a dog park [Reference Westgarth12]). The infection state of dogs in the contact network was followed-up individually for the duration of the simulation. Dogs could be:
-
Susceptible (S): uninfected with Leishmania but susceptible to infection;
-
Latent (L): exposed but not yet infectious (unable to transmit the infection to other S dogs);
-
Infectious Sub-Clinical (I): able to transmit Leishmania to S dogs but not showing clinical signs of CanL;
-
Infectious Clinical (C): able to transmit Leishmania and with clinical signs of CanL;
-
Resistant (R): exposed to Leishmania and having developed an immune response (via effective vaccination or curative treatment) preventing them from becoming infectious.
It was assumed that there are no naturally resistant dogs. C dogs were more likely to be diagnosed and treated against CanL than I dogs, and were thus modelled separately. Dogs becoming R remained in that state for the duration of the simulation [Reference Solano-Gallego1, Reference Oliva13]. A dog's probability of death was age-dependent. A proportion of dogs were replaced with S dogs after a period of time, creating new S introductions into the contact network (see detailed description of model in the Supplemental material).
Transmission from infectious to susceptible dogs occurred via sandfly bites, and was modelled using vectorial capacity (VC), representing the number of secondary cases resulting from an infectious case in time t in a fully susceptible population [Reference Dye9]:
 $${\rm VC} = m{\alpha}^2{{\rm e}^{-\mu l}}/\mu ,$$
$${\rm VC} = m{\alpha}^2{{\rm e}^{-\mu l}}/\mu ,$$
where m is the number of female sandflies per dog, α is the number of female sandfly bites per day, μ is the daily mortality rate of female sandflies, and l is the latency period of Leishmania in sandflies (l = 1/τ, the transition rate from latent to infectious, assuming an exponentially distributed latent period in the sandfly). VC was assumed to be independent of the prevalence in the sandfly population [Reference Massad and Coutinho14]. Sandflies were assumed to be present in CanL-free areas for 90 days per year, during which time transmission could thus occur.
A two-dimensional modelling approach [Reference Cohen, Lampson and Bowers15] was used to report the uncertainty in P inf and P per in the form of 95% predictive intervals (95% PI). The model was implemented in MS Excel 2010, using the Monte Carlo simulation @RISK 6·0 add-in (Palisade Corporation, USA).
Probability of introducing an infected dog into a non-endemic area (P infCA and P inf)
Commercial dog movement records were unavailable, so it was assumed that all dogs from endemic areas were equally likely to be imported. Thus, P infCA corresponds to the true prevalence of CanL infection in endemic areas, which was estimated using a Bayesian latent class model [Reference Branscum, Gardner and Johnson16] based on the IFAT seroprevalence estimates obtained from three cross-sectional studies [Reference Leontides17–Reference Baldelli19].
P inf was estimated using the individual-based stochastic model previously described (see Supplementary material) with short simulation periods representing households travels to endemic areas. Up to five dogs travelling per household were simulated. As the number of travelling dogs did not affect the results and conclusions, only results for one dog per household are reported here. Given CanL's long incubation period, it was assumed that the number of infected dogs in the endemic area remained constant for the duration of the household trip, so CanL infection and progression was only simulated for the travelling dog. The dog's infection state was evaluated daily for the duration of the trip, and recorded at the end of the simulation period. A transmission season of 150 consecutive days per year represented the period when sandflies were present in endemic areas and transmission occurred [Reference Dye, Vidor and Dereure20, Reference Oliva21]. P inf was calculated as the proportion of iterations where the travelling dog became infected.
Probability of persistence in a previously CanL-free area with competent vector (P per)
The individual-based stochastic model was also used to estimate P per. In this case, the spread of CanL was modelled within an independent contact network of dogs with seasonal presence of sandflies for 90 consecutive days. Simulations started with the introduction of an infected dog in the contact network, and CanL transmission was simulated for 3 years. Given the slow progression of the disease, the model was simulated in weekly steps and the daily parameters for transmission and VC (Table 1) were converted to weekly parameters. CanL infection was considered persistent in the network if ⩾1 L, I or C dogs (other than the initially introduced dog) remained at the end of the simulation period. P per was calculated as the proportion of iterations that led to persistence of CanL within the contact network.
Table 1. List of parameters used to model the transmission of canine leishmaniasis in a non-endemic area following the introduction of an infected dog with competent vector and the transmission of canine leishmaniasis to susceptible dogs traveling to endemic areas

n.a., Non-available.
Probability of persistence within a region (P per region)
P per region was estimated using equation (2):
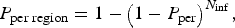 $${P_{{\rm per}\thinspace {\rm region}}} = 1 - {\left( {1 - {P_{{\rm per}}}} \right)}^{N_{{\rm inf}}},$$
$${P_{{\rm per}\thinspace {\rm region}}} = 1 - {\left( {1 - {P_{{\rm per}}}} \right)}^{N_{{\rm inf}}},$$
where N inf is the number of infected dogs introduced into the CanL-free areas given by P inf or P infCA (depending on whether the dogs were travelling to endemic areas or imported from endemic areas, respectively).
It was assumed that a proportion of the total number of dogs introduced into CanL-free areas (following commercial imports, adoptions, individual purchases from endemic areas, or travels to endemic areas) were infected with CanL, and that a proportion of those infected may transmit the infection to other dogs in their contact network and generate persistence in the previously CanL-free area.
Mitigation measures
Vaccination, prophylactic medication, repellent use, insecticide use, and diagnostic test and exclusion were modelled alone or in combination. Test and exclusion was only considered for P per region, whereas the others affected P inf and P per. The effect of MMs was modelled using the product of their level of use and their efficacy. The level of use represented the proportion of dogs treated with MMs and the efficacy represented the proportion of dogs on which it had its intended effect – with the exception of insecticide use, as discussed later. Efficacies were estimated from published data (Table 1), while different levels of use were tested via scenario analysis.
Vaccination and prophylactic medication prevented infection and therefore transitioned dogs from the S to the R state (Fig. 1). When used on S dog(s), repellents reduced the transmission of CanL by decreasing the sandfly biting rate α [equation (1)] proportionally to their efficacy, thus reducing VC. When used on I or C dog(s), repellents reduced the number of I and C dogs in the network proportionally to its efficacy (Fig. 1).
Unlike other MMs, insecticides were applied to the environment and therefore affected the entire contact network rather than individual dogs. Thus, the level of insecticide use corresponded to the proportion of contact networks on which insecticides were used, and insecticide efficacy corresponded to the proportional reduction of the sandfly density within the contact network where applied.
Dogs imported from endemic areas were tested for infection and positive dogs were denied entry (excluded). Test and exclusion use was the proportion of imported dogs that were tested, whereas efficacy was the sensitivity (Se) of the diagnostic test (i.e. probability that an infected animal is positive by the test).
It was assumed that when used, MMs were applied regularly following instructions/recommendations and thus remained efficacious during the simulation period. The effectiveness of MMs was measured in terms of the proportional reduction in the mean P inf and P per.
Scenario analysis
For P inf and P per, the effectiveness of MMs was evaluated in 20% increments of levels of use (from 0% to 100%). The combined effect of pairs of MMs was implemented for high (80%) and medium to low (40%) levels of use. Levels of use and efficacies were assumed to be independent between MMs. The following combinations were assessed: vaccination and repellent, vaccination and insecticide, repellent and prophylactic medication, repellent and insecticide, and insecticide and prophylactic medication. Vaccination and prophylactic medication were not combined as they are both aimed at inducing resistance: they were modelled using the same mechanism and the difference in their effectiveness depends on efficacy only, and it was assumed they are unlikely to be used in combination on a dog.
For P per region, test and exclusion was individually evaluated when used in 0%, 50% and 100% of the dogs moved to CanL-free areas, and also combined with repellent and vaccination (at 0% and 80% use in dogs).
P per region was calculated for 10, 100 and 5000 dogs travelling to or imported from endemic areas. The number of dogs needed to reach a P per region of 100% was also calculated.
Sensitivity analysis
A sensitivity analysis using the conditional effect of the 2·5th and 97·5th percentiles of input parameters on the mean outputs (P inf and P per) was reported.
Validation
Two cohort studies were used to validate P inf estimation. In these studies, a cohort of CanL-free dogs was introduced into an endemic area in southern Italy [Reference Baldelli19] and southern France [Reference Keck and Dereuer18], and dogs were physically examined and sampled every 1–3 months to evaluate the clinical signs of CanL and positivity to tests (PCR and IFAT). The incidence rate ratio (IRR) was used to compare P inf to the results of these field studies. As the information available on CanL outbreaks or the presence of the vector in non-endemic areas is scarce, it was not possible to validate P per estimates.
RESULTS
In the absence of MMs, the mean probability that a dog travelling to endemic areas became infected, P inf, was 7·8% (95% PI 2·6–16·4), and the mean probability of importing an infected dog from endemic areas, P infCA, was 10·7% [95% credibility interval (CrI) 1·9–22·4]. Vaccinating 100% of travelling dogs decreased P inf by 88·5%. By contrast, low levels of vaccine use (i.e. 20%) only reduced P inf by 25·6% (Fig. 2a ). Similarly, repellents used in all travelling dogs reduced P inf by 99·6% while P inf decreased by only 28·2% when used in 20% of travelling dogs (Fig. 2b ). When prophylactic medication was used in 100%, 80%, and 20% of dogs, P inf decreased by 75·6%, 61·5% and 19·2%, respectively (Fig. 2c ). P inf decreased by 55·1% when insecticide was used in all endemic areas (Fig. 2d ). All combinations of MMs with high level of use (80%) reduced P inf to ⩽2·5%. The most effective was the combination of vaccination and repellent, which reduced P inf by 94·9%, compared to a reduction of 78·2% and 80·7%, respectively, when used alone. At 40% use, the combinations of vaccination and repellent, and repellent and prophylactic medication were the most effective, reducing P inf by 45·6% and 47·7% compared to individual use of 40% for vaccine and prophylactic medication, respectively.

Fig. 2. Probability of infection [mean (solid line) and 95% predictive interval (dotted line)] of a dog after a trip to a CanL endemic area (P inf), by proportions of use of mitigation measures (black lines) compared to no use of mitigation measures (grey lines). (a) Vaccine use, (b) repellent use, (c) prophylactic medication and (d) insecticide use.
The mean number of secondary cases a week after the index case became infectious was 0·32 (range 0–3) with seasonality (Fig. 3a ) and 1·32 (range 0–5) without seasonality (i.e. uninterrupted transmission, Fig. 3b ). When no MMs were used, the mean P per following the introduction of an infectious dog was 72·0% (95% PI 67·8–76·0). When vaccination, repellent, and prophylactic medication were used on all dogs, and insecticide in the environment of all contact networks, P per decreased by 58·3%, 99·6%, 21·7% and 1·8%, respectively (Fig. 4). When repellent was used in combination with vaccination, prophylactic medication or insecticide at 80% usage each, P per decreased by 76·9%, 67·1% and 36·1%, respectively, compared to no MMs. P per also decreased when use of insecticide was combined with vaccination (22·2%) or prophylactic medication (12·5%). Com-bining MMs at 40% usage further reduced P per by only 2–7% compared to using each MM separately.

Fig. 3. Distribution of the number of secondary of canine leishmaniasis cases at the end of the first week after the introduction of an infectious dog in a non-endemic area with competent vectors when (a) seasonality was implemented, and (b) no seasonality was included.

Fig. 4. Probability of persistency (P per) [mean (solid line), 95% PI (dotted line)] following the introduction of a CanL-infected dog into a non-endemic area with competent vector, by proportions of use of mitigation measures (black lines) compared to no use of mitigation measures (grey lines). (a) Vaccine use, (b) repellent use, (c) Prophylactic medication and (d) insecticide use.
In the absence of control measures, high P inf, P infCA and P per resulted in a high P per region. A P per region of 100% was reached for 170, 240 and 350 dogs returning from travel to an endemic area when 0%, 50% and 100% of the dogs were tested and excluded if positive, respectively. The effectiveness of test and exclusion in reducing P per region was high when a very small numbers of dogs (e.g. 10) were returning from endemic areas, but quickly decreased and was almost zero for large numbers of travelling dogs (e.g. 5000, Table 2). The use of both repellent and vaccination in 80% of travelling dogs, in addition to test and exclusion of infected dogs on 100% of dogs returning to CanL-free areas decreased P per region by 98·0%, 94·9% and 32·0% for 10, 100 and 5000 travelling dogs, respectively, compared to test and exclusion only (Table 2).
Table 2. Effect of different levels of testing and exclusion and mitigation measures on the probability of CanL persistence in at least one network of a non-endemic region (Pper pegion), for different numbers of dogs travelling to endemic areas

Twenty, 30 and 80 dogs imported (commercial imports, adoptions or individual purchases) from endemic areas resulted in P per region of 100% when 0%, 50% and 100% of the animals were tested and excluded if positive, respectively (Table 3). The reduction of P per region from test and exclusion quickly diminished as N increased, and was almost zero for large numbers of dogs (e.g. 5000) moved into CanL-free areas (Table 3).
Table 3. Effect of different levels of testing and exclusion and mitigation measures on the probability of CanL persistence in at least one network of a non-endemic region (Pper region), for different numbers of dogs imported from endemic areas (commercial imports, adoptions, individual purchases)

When testing and exclusion of positives dogs was used on 0%, 50% and 100% of imported dogs in combination with repellent use in 80% of dogs in contact networks in non-endemic areas, P per region reached 100% when 270, 350 and 600 dogs were imported from endemic areas, respectively. Similarly, when testing and exclusion of positive dogs was used on 0%, 50% and 100% of imported dogs in combination with vaccination of 80% of dogs in contact networks in non-endemic areas, P per region reached 100% for 320, 450 and 710 imported dogs from endemic areas, respectively (Table 3). When testing and exclusion of positive dogs was used on 0%, 50% and 100% of imported dogs in addition to the combined use of vaccination and repellent at 80% each, a P per region of 100% was reached for 770, 1000 and 1700 imported dogs from endemic areas, respectively (Table 3).
P inf was most influenced by the day of the year travelling (transmission season), followed by the number of travelling days and CanL prevalence in the endemic area (Fig. 5a ). The most influential parameter for P per was the day of the year when the index case dog became infectious (transmission season), followed by the efficacy of MMs, VC and infection transition parameters (Fig. 5b ).

Fig. 5. Sensitivity analysis using the conditional effect of the 2·5th and 97·5th percentiles of input parameters on the (a) mean probability of infection of a dog after a trip to a CanL-endemic area (P inf) and (b) mean probability of persistency following the introduction of a CanL-infected dog into a non-endemic area with competent vector (P per).
After adjusting for the time of exposure and transmission period, the IRRs predicted by the P inf module were comparable to those from the cohort studies used for validation [Reference Dye, Vidor and Dereure20, Reference Oliva21]. Oliva et al. [Reference Oliva21] reported an incidence rate of 0·036 cases/dog per week, which is not significantly different to the 0·051 cases/dog per week incidence estimated by the model (IRR 0·71, 95% CrI 0·38–1·29). Simolarly, Dye et al. [Reference Dye, Vidor and Dereure20] reported an incidence of 0·028 cases/dog per week, which was not significantly different from the 0·025 cases/dog per week predicted by the model (IRR 1·1, 95% CrI 0·68–1·79).
DISCUSSION
The scenarios evaluated suggest that the probability of introducing CanL-infected dogs to CanL-free regions can be high and, furthermore, the probability that these dogs will transmit the infection within their contact networks can also be high. These probabilities may seem high at first inspection but are plausible as dogs exhibit a prolonged infectious period which is often sub-clinical, allowing them to spread the disease for a very long time period [Reference Solano-Gallego1, Reference Dye9] before they are identified as infected. Nonetheless, several MMs can reduce these probabilities.
Several key assumptions were made for this modelling work, one of the most important being that dogs become resistant after effective treatment. Although it is possible that some clinical dogs return to the clinical stage 6–24 months after treatment [Reference Solano-Gallego1, Reference Oliva13], it was assumed that dogs with a history of clinical infection would be monitored on a regular basis for seroconversion and clinical signs, and therefore treated as needed within a short period [Reference Solano-Gallego1]. P per may have been underestimated if R dogs can transition back to I after treatment or after losing vaccine immunity.
Information on the number of networks, size of networks, and movement of dogs between endemic and non-endemic areas within Europe was unavailable. Therefore, the estimation of the probability of persistence in a region (P per region) relied on simplified assumptions such as independence between networks, which may not reflect reality. It was also assumed that dogs moved between networks independently (i.e. any contact network has the same chance of ‘receiving’ one or more infected imported dog). P per region may have been overestimated if this assumption is unwarranted. Nonetheless, these assumptions can be easily relaxed to estimate P per region as more detailed information becomes available.
VC represents the potential of sandflies to transmit leishmaniasis between dogs and it is independent of the prevalence of L. infantum infection in sandflies [Reference Massad and Coutinho14]. VC was a parsimonious choice used to avoid modelling the transmission of L. infantum between sandflies, which occurs on a faster time-scale than in dogs, and because few sandflies live long enough to acquire infection [Reference Dye9]. The density of vectors depends on environmental factors such as temperature and humidity which are highly variable throughout the year. Given the lack of information on such heterogeneous parameters, the density of sandflies within the contact network was assumed to be constant throughout the breeding season; however, the length of the breeding season (transmission period) was assumed to be 150 days in endemic areas and 90 days in non-endemic areas, to reflect shorter summers of northern non-endemic areas of Europe. This assumption may have overestimated the initial size of the outbreak because VC may take a few weeks to reach stability at the start of the short breeding season in non-endemic areas. By contrast, by assuming a single introduction of an infected dog in a network instead of a continuous inflow of dogs, P per may have been underestimated. As P per is measured as the probability of at least one new infected dog at the end of the simulated 3-year period, it is unlikely that the above assumption affected this parameter, nor the relative effectiveness of the different MMs.
In the present study, dogs travelling to endemic areas were exposed to sandflies only for relatively short travel periods [22]. However, after adjusting for the exposure period, the model-predicted incidence agreed with that from field studies where cohorts of CanL-free dogs were introduced into endemic areas in southern Italy [Reference Oliva21] and southern France [Reference Dye, Vidor and Dereure20], and their infection status measured at the end of the sandfly breeding season, suggesting that the model predictions were valid against two independent datasets not used to estimate model parameters.
When no MMs were implemented, both P inf and P per were high (means of 7·8% and 72·0%, respectively) resulting in a P per region of 100% even for small numbers (n = 20) of dogs moved to a CanL-free region with competent vectors. CanL-infected dogs exhibit a prolonged infectious period (median ~200 days, [Reference Dye9]) which is often sub-clinical, allowing them to silently spread the disease for extended time periods, especially since no culling of infectious dogs was considered in the model. Moreover, P per region reports the probability of persistence of CanL in at least one dog in at least one contact network in a region, therefore this parameter does not readily relate to the actual disease prevalence in the region when CanL is introduced, and this prevalence may range from extremely low to extremely high.
The most effective MM in reducing P inf and P per was repellent either used individually or in combination with other MMs, followed in decreasing order of effectiveness by vaccine, prophylactic medication and insecticide. Although the ranking of MMs was similar for P inf and P per, the differences between MMs were more important for P per than P inf. P inf was modelled to estimate the probability that one susceptible dog, protected with a certain MM and travelling to an endemic area, will become infected. By contrast, P per, was modelled to estimate the probability that one infected dog introduced in a network of susceptible dogs, all protected with a certain MM, will lead to a persistent infection of at least one of these dogs in the network. The larger efficacies of the MMs observed when P per was modelled may be explained by the longer modelling time for P per than P inf. For each time step, there was a probability that a susceptible dog becomes infected, which was dependent on the effectiveness of the MMs applied to the susceptible dogs, thus the longer the modelling period, the larger the differences in the overall probability that the travelling dog becomes infected. Furthermore, for P inf, the MMs were applied to only one dog, with the exception of insecticide use, while for P per the MMs were applied to all dogs in the network, thus the efficacy of the MMs was amplified for P per.
The main mechanism of action of repellent use is to avoid sandfly bites, which reduces the VC by several-fold. Vaccination and prophylactic medication both reduce CanL transmission by inducing resistance and the slight difference in their effectiveness is due to their different efficacies. Insecticide use was shown to be the least effective MM in reducing P inf and P per. By contrast, in an earlier CanL modelling article, Dye [Reference Dye9] concluded that insecticide use was more effective than vaccination. However, Dye [Reference Dye9] modelled the disease in an endemic area, and the effect of the insecticide as a percentage of change in the mortality rate of sandflies, whereas in the present study P per was estimated in a previously CanL-free area and insecticide use was modelled as decreasing the vector density proportionally to its efficacy [Reference Coleman8].
Test and exclusion of positive dogs moving into non-endemic areas was effective for low numbers of dogs, but its benefits diminished as the number of dogs moved to CanL-free areas increased. The main drivers of these results were the high P per, the relatively high prevalence of CanL in endemic areas (P infCA range 2·0–22·6%), and the low sensitivity (52·6%, 95% CrI 30·8–74·0) of the diagnostic test used to model the test and exclusion policy. Furthermore, it was assumed that the dogs imported were a random sample from the dog population in the endemic area. However, CanL prevalence in commercial dogs (P infCA) may be lower than that of the general population since they may be younger animals and commercial breeding facilities may regularly use vaccination or repellent, or may also house dogs indoors, further reducing the exposure to sandflies, in which case the results of the import pathway may be overestimated and more comparable to those of the travelling pathway with MMs implemented. Tests with higher sensitivity would be more effective at reducing P per region, although as N increases some infected animals will inevitably go undetected and enter the non-endemic region.
The day of year when the trip to an endemic area was made, followed by the travelling length were the greatest drivers of P inf. The first parameter was assumed to randomly occur at any time of the year, although travelling to endemic areas may be concentrated during certain periods of the year. The transmission of CanL depends on the presence of the vector and is thus seasonal [Reference Oliva21]. In addition, longer trips increase the time of exposure to infected vectors.
The most influential parameter for P per was the time of the year when the index case dog became infectious. Naturally, if no vectors were present at that time, transmission was delayed until the next vector breeding season started, and could be avoided altogether if the index case dog was removed or died.
In conclusion, the model results suggest that the introduction of infected dogs in previously CanL-free regions with competent sandflies can result in a high probability of CanL persistence in the absence of MMs. The best mitigation options evaluated were: prevention of infection during travel (i.e. using repellent, vaccination, or prophylactic medication), and test and exclusion of infected dogs coming from endemic areas. The testing and exclusion policy had a high impact only for small numbers of dogs moved between endemic and free areas. The most effective measures to reduce the probability of CanL infection during travel to endemic areas and CanL persistence in a network of dogs was the use of repellent, followed closely by vaccination and prophylactic medication.
SUPPLEMENTARY MATERIAL
For supplementary material accompanying this paper visit http://dx.doi.org/10.1017/S0950268814002726.
ACKNOWLEDGEMENTS
The authors are grateful to the entomologists that provided expert opinion to parameterize the vectorial capacity. The authors also thank Laia Solano-Gallego, Claire Wylie, David Brodbelt, Madeleine Mattin and Huybert Groenendaal for their valuable inputs to the modelling work. Finally, the authors thank the EFSA scientific staff who provided input and guidance during this project, including Sofie Dhollander, José Cortinas Abrahantes, and EFSA AHAW panel members Aline de Koeijer and Edith Authié. This study was financed by EFSA (CFT/EFSA/AHAW/2012/02). The present article is published under the sole responsibility of the authors and may not be considered as an EFSA scientific output. For the views or scientific outputs of EFSA, please consult its website under http://www.efsa.europa.eu.
DECLARATION OF INTEREST
None.










