


INTRODUCTION
Although a global effort has been made to manage H5N1 highly pathogenic avian influenza (HPAI), the virus has not yet been controlled. This failure is of international concern because of the continued impact of outbreaks in animal populations [Reference Hamilton and Bruckner1, 2], and the risk of a major human influenza pandemic that might result from mutations and re-assortment of H5N1 with human influenza viruses [Reference Rajagopal and Treanor3]. No human pandemic has occurred to date, but almost a decade after the re-emergence of the Asian lineage of H5N1, outbreaks in poultry and human deaths are still occurring [Reference Fournié, Glanville, Pfeiffer, Zilberman, Otte, Roland-Holst and Pfeiffer4]. Outbreaks of clinical disease and deaths in poultry have occurred in various countries [Reference Fournié, Glanville, Pfeiffer, Zilberman, Otte, Roland-Holst and Pfeiffer4–Reference Penny7] and HPAI H5N1 is currently endemic in several countries including Egypt, Vietnam and Indonesia [Reference Farnsworth8].
Why are we not able to control HPAI H5N1 despite substantial funding for control and research studies? Several studies have been conducted to identify risk factors associated with occurrence of HPAI outbreaks; putative factors assessed have included chicken and domestic waterfowl population density, specific climate/vegetation factors and human population density [Reference Martin9–Reference Henning, Pfeiffer and Vu12], but longitudinal studies identifying risk factors for incident infections are rare. Data from outbreaks reported to veterinary authorities and potential risk-factor data (e.g. poultry and human density, land use, elevation data) obtained from national statistics or land use databases are easier to obtain than prospective sampling and testing of poultry populations over long periods combined with regular duck farmer interviews. Thus studies focusing on risk factors for outbreaks can be more practical and less expensive. Such studies may identify mechanisms that lead to occurrence of outbreaks. However, HPAI H5N1 infection of ducks does not always result in outbreaks, as ducks can harbour the HPAI H5N1 virus asymptomatically [Reference Swayne13]. Identification of causal risk factors for infection would allow active intervention in the management of ducks, to prevent introduction of H5N1 virus into duck flocks and possible HPAI virus spread to chickens.
Two duck management systems are common in Indonesia: the home-based system and the fully mobile herding system [Reference Ketaren14]. In the home-based system ducks are allowed to leave the farm and scavenge during the day, but are confined at home overnight – hence these duck flocks remain ‘stationary’ [Reference Henning15]. We focused our investigations on the ‘stationary’ system and conducted a prospective longitudinal study (1) to describe the incidence of H5 infection in stationary duck flocks in central Java, Indonesia, and (2) to identify risk factors associated with H5 infection of these duck flocks.
MATERIALS AND METHODS
Study design and data collection
A prospective longitudinal study of duck farms was conducted from March 2007 until March 2008 in four districts of central Java, Indonesia. A total of 96 smallholder duck farms (farms with 10–700 ducks) were selected; sample size calculations and the multi-stage sampling strategy have been described previously [Reference Henning16]. Farms were visited 2-monthly by field veterinarians. For the purposes of this study, ducks were considered to have been managed as a single flock. At the first visit, the flock owner confined all ducks and selected the first 10 that could be caught; these ducks were marked with wing tags or leg bands and blood collected from the wing vein [Reference Henning16]. At each subsequent 2-monthly visit, blood samples were collected from the same ducks [Reference Henning16]. Further, at each 2-monthly visit, an interview was conducted with the flock owner to monitor how flocks were managed between visits. A questionnaire containing 36 questions was used to record information on the structure of the duck flock, trade of poultry and poultry products, hatching of birds, production performance of layer birds, health status of birds including mortalities, duck scavenging practices, contact between ducks and chickens, housing and feeding of birds, any duck farm visits by various groups of people and any possible contacts of duck farmers with animals or animal products on other farms or markets, as well as information on avian influenza vaccinations. Each interview took about 20 min. The questionnaire was developed by one of the authors (J.H.) in English, then translated into Bahasa Indonesia and administered by a field veterinarian fluent in that language. The questionnaire was pilot-tested in six farms that were not part of the 96 farms selected, resulting in minor modifications of some questions.
Diagnostics
Serum was tested for H5 antibodies using the haemagglutination inhibition (HI) test following the World Organisation for Animal Health guidelines, using 4 haemagglutinating units per well [17]. The antigen and control antiserum used for the HI test were provided by Pusat Vetenerinaria Farma (Indonesia) [Reference Henning16]. Samples with a titre of ⩾1/16 (24) were classified as positive [17]; other samples were classified as negative.
Data analyses
Data were entered into a Microsoft Access 2003 database (Microsoft Corporation, USA) for data storage and data manipulation. Data analyses were conducted in Stata v. 11.0 (Stata Corporation, USA). Each farm visit with sampling of ducks was designated a flock sampling. Each 2-monthly interval from one flock sampling to the next was defined as a flock period. Each bird contributed 1 bird-day for each day between successive samplings of that bird, and each flock contributed 1 flock-day for each day in a flock period.
Incidence rate for H5 antibodies
Incidence rates were calculated at bird- and flock-level separately for groups of flock periods ending in May 2007, July 2007, etc. to March 2008. Flock-level incidence rate was also calculated for all eligible flock periods over the entire study period.
Only birds that tested seronegative initially and were also retested at the end of the flock period were used to calculate the bird- and flock-level incidence rates for those flock periods. Replacement birds first tested at the end of the flock period did not contribute to the definition of outcome statuses on bird and flock levels.
Bird-level incidence rate for a group of flock periods=
 $$\openup-2{\eqalign{ \tab \lpar {\rm number\ of\ tested\ birds\comma \ initially\ seronegative\comma \ that\ were } \ \cr \tab {\rm H5\ antibody\ seropositive\ at\ the\ next\ flock\ sampling\rpar } \cr} \over \eqalign{ \tab \lpar {\rm sum\ of\ bird {\hbox{-}} days\ between\ flock\ samplings\ for\ birds\ that\ } \cr \tab {\rm were\ seronegative\ at\ the\ next\ flock\ sampling \plus }{\raise0.5ex\hbox{$\scriptstyle 1$} \kern-0.1em/\kern-0.15em \lower0.25ex\hbox{$\scriptstyle 2$}}\lowast {\rm \ sum} \cr \tab {\rm of\ bird {\hbox{-}} days\ between\ flock\ samplings\ for\ birds\ that\ were\ } \cr \tab {\rm seropositive\ at\ the\ next\ flock\ sampling\rpar }{\rm .} \cr} }$$
$$\openup-2{\eqalign{ \tab \lpar {\rm number\ of\ tested\ birds\comma \ initially\ seronegative\comma \ that\ were } \ \cr \tab {\rm H5\ antibody\ seropositive\ at\ the\ next\ flock\ sampling\rpar } \cr} \over \eqalign{ \tab \lpar {\rm sum\ of\ bird {\hbox{-}} days\ between\ flock\ samplings\ for\ birds\ that\ } \cr \tab {\rm were\ seronegative\ at\ the\ next\ flock\ sampling \plus }{\raise0.5ex\hbox{$\scriptstyle 1$} \kern-0.1em/\kern-0.15em \lower0.25ex\hbox{$\scriptstyle 2$}}\lowast {\rm \ sum} \cr \tab {\rm of\ bird {\hbox{-}} days\ between\ flock\ samplings\ for\ birds\ that\ were\ } \cr \tab {\rm seropositive\ at\ the\ next\ flock\ sampling\rpar }{\rm .} \cr} }$$For flock-level incidence rate calculations, we only used flock periods where all birds tested at the start of the flock period were seronegative (i.e. flock periods that had initially seropositive birds were excluded as well as flock periods that had birds that tested initially seropositive, but were not retested).
Flock-level incidence rate for a group of flock periods=
 $$\openup-2{\hskip-8\eqalign{ \tab \lpar {\rm number\ of\ flock\ periods\ where\ all\ sampled\ birds\ were} \cr \tab {\rm initially\ seronegative\comma \ but\ where\ }\ges {\rm 1\ of\ these\ birds\ were} \cr \tab {\rm H5\ antibody\ seropositive\ at\ the\ next\ flock\ sampling\rpar } \cr} \over \eqalign{ \tab \lpar {\rm sum\ of\ flock {\hbox{-}} days\ between\ flock\ samplings\ for\ flocks} \cr \tab {\rm where\ all\ retested\ birds\ were\ seronegative\ at\ the\ next\ } \cr \tab {\rm flock\ samplings \plus }{\raise0.5ex\hbox{$\scriptstyle 1$}\kern-0.1em/\kern-0.15em \lower0.25ex\hbox{$\scriptstyle 2$}}\lowast {\rm \ sum\ of\ flock {\hbox{-}} days\ between\ flock} \cr \tab {\rm samplings\ for\ flocks\ where\ all\ sampled\ birds\ were\ initially} \cr \tab {\rm seronegative\comma \ but\ where\ }\ges {\rm 1\ of\ these\ birds\ were\ H5} \cr \tab {\rm antibody\ seropositive\ at\ the\ next\ flock\ sampling\rpar } \cr} }$$
$$\openup-2{\hskip-8\eqalign{ \tab \lpar {\rm number\ of\ flock\ periods\ where\ all\ sampled\ birds\ were} \cr \tab {\rm initially\ seronegative\comma \ but\ where\ }\ges {\rm 1\ of\ these\ birds\ were} \cr \tab {\rm H5\ antibody\ seropositive\ at\ the\ next\ flock\ sampling\rpar } \cr} \over \eqalign{ \tab \lpar {\rm sum\ of\ flock {\hbox{-}} days\ between\ flock\ samplings\ for\ flocks} \cr \tab {\rm where\ all\ retested\ birds\ were\ seronegative\ at\ the\ next\ } \cr \tab {\rm flock\ samplings \plus }{\raise0.5ex\hbox{$\scriptstyle 1$}\kern-0.1em/\kern-0.15em \lower0.25ex\hbox{$\scriptstyle 2$}}\lowast {\rm \ sum\ of\ flock {\hbox{-}} days\ between\ flock} \cr \tab {\rm samplings\ for\ flocks\ where\ all\ sampled\ birds\ were\ initially} \cr \tab {\rm seronegative\comma \ but\ where\ }\ges {\rm 1\ of\ these\ birds\ were\ H5} \cr \tab {\rm antibody\ seropositive\ at\ the\ next\ flock\ sampling\rpar } \cr} }$$Flock-level incidence rate was calculated for the entire study period as a pooled incidence rate.
Flock-level incidence rate over the entire study period=
 $$\openup-2pt{\hskip-2pt\eqalign{ \tab \lsqb {\rm number\ of\ flocks\ where\ all\ sampled\ birds\ were\ initially} \ \cr \tab {\rm seronegative\ but\ where\ }\ges {\rm 1\ of\ these\ birds\ were\ H5} \ \cr \tab {\rm antibody\ seropositive\ across\ all\ flock\ periods} \ \cr \tab {\rm \lpar i}{\rm .e}{\rm .\ the\ sum\ of\ the\ numerators\ used\ for\ the\ flock {\hbox{-}} level} \cr \tab {\rm incidence\ rate\ calculations\rpar \rsqb } \cr} \over \hskip-16pt\eqalign{ \tab \lsqb {\rm sum\ of\ all\ flock {\hbox{-}} days\ at\ risk\ across\ all\ flocks\ periods}\hfill \cr \tab {\rm \lpar i}{\rm .e}{\rm .\ the\ sum\ of\ the\ denominators\ used\ for\ the}\hfill \cr \tab {\rm flock {\hbox{-}} level\ incidence\ rate\ calculations\rpar \rsqb }\hfill \cr} }$$
$$\openup-2pt{\hskip-2pt\eqalign{ \tab \lsqb {\rm number\ of\ flocks\ where\ all\ sampled\ birds\ were\ initially} \ \cr \tab {\rm seronegative\ but\ where\ }\ges {\rm 1\ of\ these\ birds\ were\ H5} \ \cr \tab {\rm antibody\ seropositive\ across\ all\ flock\ periods} \ \cr \tab {\rm \lpar i}{\rm .e}{\rm .\ the\ sum\ of\ the\ numerators\ used\ for\ the\ flock {\hbox{-}} level} \cr \tab {\rm incidence\ rate\ calculations\rpar \rsqb } \cr} \over \hskip-16pt\eqalign{ \tab \lsqb {\rm sum\ of\ all\ flock {\hbox{-}} days\ at\ risk\ across\ all\ flocks\ periods}\hfill \cr \tab {\rm \lpar i}{\rm .e}{\rm .\ the\ sum\ of\ the\ denominators\ used\ for\ the}\hfill \cr \tab {\rm flock {\hbox{-}} level\ incidence\ rate\ calculations\rpar \rsqb }\hfill \cr} }$$Bird and flock-level incidence rates were multiplied by 1000 to express rates per 1000 bird-days (or 1000 flock-days) at risk and further multiplied with 365·25 to express rates per 1000 bird-years (or 1000 flock-years) at risk.
The standard errors for incidence rates were calculated as [Reference Dohoo, Martin and Stryhn18]:
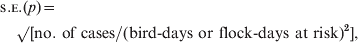 $$\eqalign{\tab {\scaps{\rm s}}.{\sc {\rm E}}. \lpar p\rpar\equals \cr\tab\quad {\rm \surd \lsqb {\rm no}{\rm .\ of\ cases\sol \lpar bird{\hbox{-}} days\ or\ flock{\hbox{-}}days\ at\ risk\rpar }^{\setnum{2}} \rsqb \comma \cr} $$
$$\eqalign{\tab {\scaps{\rm s}}.{\sc {\rm E}}. \lpar p\rpar\equals \cr\tab\quad {\rm \surd \lsqb {\rm no}{\rm .\ of\ cases\sol \lpar bird{\hbox{-}} days\ or\ flock{\hbox{-}}days\ at\ risk\rpar }^{\setnum{2}} \rsqb \comma \cr} $$where cases were number of tested birds, initially seronegative, that had seroconverted by the next flock sampling or number of tested flocks where all sampled birds were initially seronegative, but where ⩾1 of these birds had seroconverted by the next flock sampling. The 95% confidence intervals were calculated as incidence rate ±1·96∗s.e.(p).
Risk factor analyses
For risk factor analyses, we used logistic General Estimation Equation (GEE) models with Stata's xtgee command, with exchangeable correlation structures and with flock as the grouping (i.e. panel) variable. The flock period was used as the unit of analysis and the dependent (i.e. outcome) variable was the flock seroconversion status during the flock period. GEE logistic models (which produce population-averaged estimates) were chosen over random-effects logistic regression (which produce subject-specific estimates) as we were interested in estimating effects of risk factors across flocks rather than within any particular flock. Each flock period was classified as seroconverting (coded as 1) where the flock developed H5 antibodies (i.e. where all sampled birds in the flock were initially seronegative but where ⩾1 of these birds were H5 antibody seropositive at the next sampling) or non-seroconverting (coded as 0) where the flock did not developed H5 antibodies (i.e. where all tested birds were seronegative at both flock samplings). Only flock periods where all birds tested at the start of the flock period were seronegative were used in these analyses; these included flock periods with birds initially tested but not retested. However, only birds that tested seronegative initially that were retested at the end of the flock period were used to determine the seroconversion status of the flock period. A total of 129 dichotomous, nominal and ordinal potential risk factors were derived from the questionnaire data. We fitted study month number in which the flock period ended as a categorical variable in all (bivariable and multivariable) models to remove any confounding due to additional factors that varied over time. Initially bivariable analyses were conducted to identify risk factors to be included in a multivariable modelling process; those with bivariable P values ⩽0·2 were selected. The multivariable model was built with a backward elimination procedure, hence the maximum model was fit and then risk factor variables were sequentially removed, with the variable with the highest P value at each step removed, until all variables remaining in the model had P values <0·05. Once removed, a variable was not eligible for re-entry into the model during the model building with the exception that some initially removed variables that were identified as potentially important risk factors based on a priori considerations were forced into the final model and retained if their P value in that model was <0·05. Huber–White sandwich (‘robust’) estimators of variance were used for all models. Joint Wald tests performed with the testparm command were used to test the overall significance of risk factor variables with more than two levels. In contrast to General Linear Models (GLMs) which are based on maximum-likelihood estimations, the GEE method is based on the quasi-likelihood theory [Reference Wedderburn19]. Therefore, Akaike's Information Criterion (AIC), a widely used method for model selection in GLM, is not directly applicable to GEE [Reference Cui20]. Accordingly, the model selection was confirmed based on the quasi-likelihood under the independence model criterion (QIC), which is an extension of the AIC criterion [Reference Cui20]. The QIC was calculated in Stata using the qic command. Under the exchangeable correlation structure, the subset of covariates with the smallest QIC was the preferred model.
Cramer's coefficient V was calculated to assess the correlation between dichotomous variables. Multicollinearity of exposure variables was assessed using the variance inflation factor (VIF), which was estimated using the collin command in Stata (http://www.nd.edu/∼rwilliam/stats2/l11.pdf). The mean VIF was calculated to express the overall collinearity of the exposure variables remaining in the final multivariable model. The correlation matrix used in the final model was obtained using the wcorrelation command.
Goodness-of-fit and discriminatory ability of the final model were assessed as for ordinary logistic regression models [Reference Hosmer, Lemeshow, Hosmer and Lemeshow21], using linear predicted probabilities for each of the flock periods included in the final model. Nine groups were used for the Hosmer–Lemeshow goodness-of-fit table and statistic as deciles could not be calculated due to the limited number of covariate patterns. Area under the receiver-operating characteristic (ROC) curve was assessed using a binormal model fitted with Stata's rocfit command.
RESULTS
Incidence rates of ducks and duck flocks developing H5 antibodies
Bird-level incidence rates are shown in Figure 1. For the flock periods ending in May, July, September, and November 2007 and in January and March 2008, a total of 48 024, 52 464, 45 850, 48 347, 48 144 and 41 584 bird-days at risk were available, respectively. In these flock periods, a total of 872, 837, 780, 783, 777 and 758 birds were monitored over the respective 2-month intervals. Bird-level incidence remained stable during the first three samplings, i.e. reflecting the period from March to September 2007, and then decreased to a low for flock periods ending in January 2008, followed by a rise again in January to March 2008.

Fig. 1. Bird-level incidence rates (number of birds developing H5 antibodies per 1000 bird-years at risk) for flock periods ending in May 2007, July 2007, etc., to March 2008 in smallholder stationary duck flocks in Java, Indonesia. Error bars indicate point-wise 95% confidence intervals.
Flock-level incidence rates are shown in Figure 2. For the flock periods ending in May, July, September, and November 2007 and in January and March 2008, a total of 2976, 3128, 2379, 3086, 2949 and 2729 flock-days at risk were available, respectively. In these flock periods, a total of 61, 59, 43, 53, 51 and 57 flocks were monitored over the respective 2-month intervals.

Fig. 2. Flock-level incidence rate (number of flocks developing H5 antibodies per 1000 flock-years at risk) for flock periods ending in May 2007, July 2007, etc., to March 2008 in smallholder stationary duck flocks in Java, Indonesia. Error bars indicate point-wise 95% confidence intervals.
Flock-level incidence rates were high at the first samplings (i.e. reflecting the period from March to May 2007), peaked for flock periods ending in July 2007 and then decreased and remained low until January 2008, before rising again in January to March 2008.
The pooled flock-level incidence rate for the entire study period was 1673·0 seroconverting flocks per 1000 flock-years at risk (95% CI 1304·1–2041·9).
Risk factors for flocks developing H5 antibodies
A total of 310 flock periods from 88 flocks were used to assess potential risk factors; 75 (24·2%) of these flock periods were seroconverters. Eight flocks from the 96 enrolled flocks contributed no flock periods, because they had no flock periods where all tested birds were initially seronegative. No flock periods where all tested birds were initially seronegative were excluded. Results of the bivariable analyses are shown in Table 1. Of 98 potential risk factors assessed, 23 were significant at P < 0·2 in the bivariable analysis and were included in the multivariable modelling process. The 75 non-significant risk factors at P ⩽ 0·2 are listed in the Table 2 and further details on these non-significant risk factors are provided in Supplementary Table S1 (available online).
Table 1. Number of flock periods and results of bivariable analyses of potential risk factors for duck flocks in central Java developing H5 antibodies during flock periods between March 2007 and March 2008

a Within a flock, the ducks could scavenge on any or all of these locations during the same flock period.
b Deaths usually associated with human involvement (i.e. birds killed by cars, motorbikes, buses, etc.).
c Deaths occur suddenly without prolonged clinical signs detected.
d Birds could be disposed by either of these means during a flock period.
e Birds can be confined by either of these means during a flock period.
f Adjusted for study month number in which flock period ended.
g Birds refers to both ducks and chickens.
h Results of joint Wald tests.
Table 2. Non-significant risk factors (P ⩽ 0·20) in the bivariable analysis for duck flocks in central Java developing H5 antibodies during flock periods between March 2007 and March 2008a

a Information on the number and percent of flock periods in which H5 antibodies were developed, including the odds ratio (with 95% confidence interval) and the P values are provided in Supplementary Table S1.
Four variables were retained in the final multivariable model (‘Dead birds consumed by the family’, ‘Duck scavenging around neighbouring houses’, ‘Duck confinement overnight on the farm’, and ‘Sudden deaths of birds’; Table 3). The variables ‘Duck scavenging on own rice paddies’ and ‘Duck confinement overnight in the rice paddies’ were then each separately forced into the final model, but neither was significantly associated with development of H5 antibodies when adjusted for the variables in the final multivariable model (OR 0·51, 95% CI 0·25–1·03, and OR 1·6, 95% CI 0·50–5·36, respectively).
Table 3. Final multivariable model of risk factors associated with duck flocks in central Java developing H5 antibodies during flock periods between March 2007 and March 2008

OR, Odds ratio; CI, confidence interval.
a Adjusted for study month number in which flock period ended and the other three explanatory variables listed.
Duck scavenging around neighbouring houses within the village was associated with increased risk of the flock developing H5 antibodies. Consumption of carcases of dead birds by the family was also associated with increased risk but this was based on only six flock periods that were exposed over the study period. Duck flocks confined overnight in enclosures on the farm were at reduced risk of developing H5 antibodies, and surprisingly, development of H5 antibodies was less likely in flock periods in which birds on the farm died suddenly.
The mean variance inflation factor of the four variables in the final multivariable model was 1·01, indicating a low degree of multicollinearity. Under the exchangeable correlation structure, we fitted different models with different subsets of covariates and the final model in Table 3 had the smallest QIC, thus it was the best fitting model to the data. The fitted correlation between residuals for repeated flock periods within the same flock in the final model was 0·08.
The final model fitted the data well. Observed numbers of flock periods in each of the nine groups based on predicted probabilities were close to expected numbers and the P value for the Hosmer–Lemeshow goodness-of-fit statistic was 0·90, providing no evidence of poor fit. However the discriminatory ability of the final model was limited. Area under the ROC curve was 0·72 (95% CI 0·65–0·78), indicating just acceptable discrimination [Reference Wedderburn19]. The sum of sensitivity and specificity of the final model was maximized at a probability threshold of 0·25. At this threshold, sensitivity and specificity of the final model were 0·72 and 0·70, respectively. This limited discriminatory ability is likely to be because the study flock periods were exposed to additional unmeasured risk factors that determine odds and probability of seroconversion.
DISCUSSION
These are the first published longitudinal results describing the incidence of development of H5 avian influenza antibodies in stationary duck populations and factors associated with risk of H5 antibody development. The bird-level sensitivity and specificity of the HI for detecting previous exposure to avian influenza virus have been estimated as 99% and 90%, respectively [Reference Yamamoto22]. However, test sensitivity and specificity are highly dependent on choice of antigen and the antigen used in that research differed from that used in our study. We are not aware of any previous studies describing HPAI incidence in ducks in Indonesia prior to the current study. However, HPAI infection was known to be spreading in poultry in Indonesia at the time of the current study [23] so the prior probability of infection was not negligible in study flocks. Accordingly, even with imperfect bird-level specificity, a substantial proportion of birds that developed H5 antibodies would have been exposed to H5 field virus. With up to 10 birds tested at sequential flock visits, and flocks classified as seroconverting if ⩾1 bird became seropositive, specificity for seroconversion at the flock level would have been lower. However two of the four risk factors identified for flocks seroconverting were highly biologically plausible based on prior knowledge, suggesting that the flock-level specificity for seroconversion was at least modest.
An apparent seasonality of infection incidence was observed. Flock-level incidence rate peaked in May–July 2007. As times from infection to development of H5 antibodies in ducks are relatively short [Reference Middleton24], this probably reflects flocks becoming infected during this period. This peak corresponds with seasonal patterns in HPAI clinical outbreak peaks described previously [Reference Henning16]. From July 2007 to January 2008, flock-level incidence was low before increasing between January and March. Despite the serological patterns observed in our study flocks, few ducks in the study died from HPAI infection and the majority remained healthy during the study period [Reference Henning16]. Our study was conducted with ducks that were farm-based, although allowed to scavenge during the day; therefore they are described as ‘stationary’ ducks and our findings must be viewed in this context. Under the other duck management system practised in South East Asia, in which ducks are moved throughout the country (often described as ‘moving’, ‘mobile-herding’ or ‘nomadic’ ducks), risk of H5 infection might be related to other practices specific to that management system.
Seasonality of infection has been variously attributed to scavenging in post-harvest rice harvest rice paddies, climatic conditions [Reference Gilbert, Slingenbergh and Xiao25], frequency of poultry trading [Reference Sanchez-Vizcaino26] and temporary wild-bird abundance [Reference Prosser27]. Rainfall data for the years 2007–2009 in the study district of Bantul in Indonesia (http://hukum.bantulkab.go.id/unduh/peraturan-bupati/2011/28) are shown in Supplementary Figure S1 (available online). In the current study, flock-level incidence of infection was highest during the dry season, when rainfall was low and at times when most of the rice harvest would have been completed [28] and ducks allowed to scavenge on the spilled rice in the paddies. However, the association between rice-cropping, duck density and HPAI outbreaks in Indonesia has been described as being not as strong as in countries in the Mekong Delta [Reference Gilbert29]. Nevertheless, it seems likely that both climatic conditions in the dry season (possibly promoting virus survival in the environment) and the rice-farming pattern that influences the duck management during this period, increase the rate of transmission of HPAI virus.
In contrast to the flock-level incidence rate, bird-level incidence rate remained high after July 2007, peaking in the period from July to September 2007 and then sharply declining until January. The difference in timing of peaks between flock-level and bird-level incidence is likely to be the result of methodologies used to calculate the incidence rates. For the flock-level incidence calculations, a flock with seropositive birds could not contribute to the subsequent flock period (because flocks that had initially seropositive birds were excluded, while for bird-level analyses, all the seronegative birds that continued to be monitored contributed, including those in seropositive flocks. Thus, birds that seroconverted in flocks that had been removed from flock-level analyses would still be included in bird-level analyses even though their flock was not included in flock-level analyses. Such birds probably contributed to the later peak in bird-level incidence compared to the flock-level peak. This could occur if infection continued to be transmitted through the flock in the flock period subsequent to the period when infection entered the flock.
Scavenging of ducks around neighbouring houses of the village increased the risk of flocks developing H5 antibodies. This activity may have increased risk of contact with other birds, people and other possible sources of infection. The number of infections occurring is probably influenced by the contact rate, the survival of the virus in the environment, the amount of virus shed by infected birds and the ability of the virus to establish infection in susceptible birds. If both environmental conditions for virus survival and the infectiousness of the virus in ducks are limited, a high contact rate will be important in permitting new infections. This highlights the importance of the preventive measure of separating stationary duck flocks from other flocks, not only in their scavenging areas, but also in their village environment. In contrast, there was no evidence from bivariable results that scavenging of ducks on the farmer's own rice paddies (where usually no other duck flocks than the farmers' own flock were allowed) was associated with increased risk of infection, this variable was not selected in the multivariable selection process, and was not significantly associated with development of H5 antibodies when refitted with the variables in the final multivariable model. Scavenging on the farmer's own rice paddies was in fact negatively correlated with scavenging of the ducks around the neighbouring houses of the village (Cramer's coefficient V = − 0·10).
Flocks in which the farmers reported sudden deaths of ducks in the previous 2 months were less likely to seroconvert over that time. This was counter-intuitive as the cause of the sudden deaths was not known for all cases, but based on the results from cases investigated by veterinary laboratories, most of these birds died from HPAI. In the 25 flock periods where the farmer reported sudden deaths, the carcases of the birds were always removed. In the majority of flock periods, duck owners buried the carcases (13/25), but carcasses were also collected by the veterinary authorities for further diagnosis (9/25), burned by the farmer (5/25) or thrown into a river (3/25). Some owners employed more than one disposal method. Farmers may have disposed of carcases as they attempted to prevent further spread of HPAI infection within the flock, and the negative association between reporting sudden deaths and flock seroconversion may be because the farmers reporting sudden deaths of ducks were more aware of biosecurity, including risks associated with these carcases, and were more likely to have been implementing other unmeasured biosecurity measures.
The consumption of carcases of dead birds by the family was strongly associated with H5 seroconversion of the flock. This association should be viewed with some caution as such consumption occurred during only six flock periods on a total of five farms (during one flock period on four farms and during two flock periods on one farm). This association could be explained if the birds consumed were infected with HPAI. Such birds may have been contagious before death as their carcases might contain a high concentration of virus [Reference Bingham30]. Thus, through the process of slaughtering and disposal of the remains, the virus might have been spread across the farm, resulting in transmission of infection to other ducks on the farm. The causes of death of birds that were consumed is unknown but it is possible that most were sick birds in the terminal stages of a disease and/or birds injured with a risk of dying were slaughtered and consumed. On farms in the six flock periods where carcases were consumed, ducks died during three flock periods from unspecified ‘accidents’ and during three flock periods from illnesses not further specified. No sudden deaths of birds were reported in these flock periods. Hence it seems that most consumed birds were slowly dying birds or terminally ill birds slaughtered before succumbing naturally and few were birds that died suddenly. This may be because these farmers were aware of the risks associated with eating suddenly succumbed birds in areas where H5 avian influenza is endemic, but when deaths occurred after slow-progressing diseases, these carcases were considered to be appropriate for eating. Variables describing consumption and sudden deaths of birds in the final multivariable model include consumption and sudden deaths of both ducks and chickens. However, mortality of chickens was more common over the study period than of ducks [Reference Henning16].
If birds were confined overnight on the farm, the risk of developing of H5 antibodies was reduced, most likely because of reduced risk of contact with potentially infected birds or other sources of infection. Contact with wild birds, potentially infected with HPAI H5N1, is probably more likely to happen when ducks are confined overnight in the rice paddies rather than on the farm. In fact, confinement of ducks overnight in the rice field was associated with increased risk of development of H5 antibodies in the bivariable analysis, although this variable was not selected in the multivariable selection process and was not significantly associated with development of H5 antibodies when refitted with the variables in the final multivariable model.
Our study differed from other HPAI risk factor studies, which have used administrative information or census data on duck densities, and numbers of ducks, duck farms and HPAI outbreaks at, village, subdistrict or district level, and analysed these data to identify associations between duck densities and HPAI outbreaks [Reference Gilbert31, Reference Gilbert32]. In contrast, we focused on associations between farm-management factors and H5 antibodies (rather than clinical disease) in stationary duck flocks. Our study is also the first to estimate incidence rates for the development of H5 antibodies in duck flocks, where the majority of ducks appeared to be clinically healthy. Scavenging around neighbouring houses and confinement overnight were independently associated with development of H5 antibodies and are likely to be causal factors for infection by H5 virus. Messages about these factors should be included in awareness and education programmes aimed at changing farmers' attitudes and management practices to reduce the risk of HPAI virus introduction into susceptible flocks. It is perhaps unlikely that in the near-future HPAI will be eradicated from most countries that are currently endemically infected, including Indonesia. However, based on our results, practical and simple interventions can reduce risks of HPAI infection of village poultry, and consequently, risks to duck owners and their families.
SUPPLEMENTARY MATERIAL
For supplementary material accompanying this paper visit http://dx.doi.org/10.1017/S0950268812001100.
ACKNOWLEDGEMENTS
This project was funded through a grant provided by the Australian Centre for International Agricultural Research (project no. AH/2004/040).
DECLARATION OF INTEREST
None.







