



Introduction
Regulations regarding aeronautical operations in natural and protected areas are still poorly defined, although they do include some restrictions relating to migration routes of large birds and prohibitions/controls in certain locations (e.g., over wetlands or national parks; DeVault et al. Reference DeVault, Blackwell and Belant2013, Moreno-Opo & Margalida Reference Moreno-Opo and Margalida2017, Tobajas et al. in press). There are huge differences in the regulation of protected areas between different habitat types, countries and even regions, which make it extremely difficult to develop and apply effective global measures. The use of remotely piloted aircraft (e.g., drones) is growing not only due to their numerous applications in civil fields (Crutsinger et al. Reference Crutsinger, Short and Sollenberger2016), but also due to their popularity for recreational purposes (Rebolo-Ifrán et al. Reference Rebolo-Ifrán, Grilli and Lambertucci2019). The low-flying characteristics of drones and their ease of use in remote areas require assessments of the risks and possible effects that they may have on wildlife (Mo & Bonatakis Reference Mo and Bonatakis2021). However, although studies on the effects of drones on wildlife have increased significantly in recent times (Mulero-Pázmány et al. Reference Mulero-Pázmány, Jenni-Eiermann, Strebel, Sattler, Negro and Tablado2017, Bennitt et al. Reference Bennitt, Bartlam-Brooks, Hubel and Wilson2019, Weston et al. Reference Weston, O’Brien, Kostoglou and Symonds2020), there have been very few studies on the effects on wildlife of the use of free-flight aircraft or gliders (i.e., ultralights, paragliders, hang-gliders and their powered derivatives; Larson et al. Reference Larson, Reed, Merenlender and Crooks2016, Marion et al. Reference Marion, Davies, Demšar, Irvine, Stephens and Long2020). As has been shown in recent studies on drones (McEvoy et al. Reference McEvoy, Hall and McDonald2016, Brisson-Curadeau et al. Reference Brisson-Curadeau, Bird, Burke, Fifield, Pace, Sherley and Elliott2017, Weimerskirch et al. Reference Weimerskirch, Prudor and Schull2018) and previously regarding aeroplanes (e.g., Harms et al. Reference Harms, Fleming and Stoskopf1997, Hughes et al. Reference Hughes, Waluda, Stone, Ridout and Shears2008, Linley et al. Reference Linley, Kostoglou, Jit and Weston2018), the flights of aircraft can affect different species in different ways, and these effects can vary according to: the flight frequency in an area (animals may habituate to them); the type or shape of the aircraft; whether the animals impacted live in groups or are solitary; the time of year or day; or the reproductive status of the animals concerned (Zeitler & Georgii Reference Zeitler and Georgii1994, Verhulst et al. Reference Verhulst, Oosterbeek and Ens2001, Boldt & Ingold Reference Boldt and Ingold2005, Mulero-Pázmány et al. Reference Mulero-Pázmány, Jenni-Eiermann, Strebel, Sattler, Negro and Tablado2017, Barnas et al. Reference Barnas, Newman, Felege, Corcoran, Hervey and Stechmann2018). As a result, it is fundamentally important to evaluate the specific potentially negative effects of particular types of aircraft on particular species or groups of species in particular locations (Marion et al. Reference Marion, Davies, Demšar, Irvine, Stephens and Long2020).
Free-flights and their derivatives mainly comprise paragliding and hang-gliding, in which the aircraft gains altitude using thermal air currents; however, these activities also include powered versions of these same craft whereby auxiliary engines are used to take off and gain altitude (i.e., powered hang-gliders and paramotors). Also included in this group are small aircraft and sailplane gliders, which, as occurs with small aeroplanes, operate from aerodromes. A common characteristic of this group is that these are usually towed to flying height by a motorized aircraft or, in the case of motorsailers, by an auxiliary engine that is stowed in the fuselage after ascent.
Most of the regulations regarding free-flight and its derivatives in natural areas relate to sensitive times of the year and the areas around raptor nests (Tobajas et al. in press). According to the species concerned, in some countries buffer areas of between 500 and 1000 m have been established in protected or otherwise vulnerable areas for wildlife, overflights are usually prohibited (either throughout the year or only during times of greatest wildlife sensitivity) or minimum flight height levels are prescribed for the entire year or at sensitive times. However, the lack of scientific studies and guidance on the effects of flights on wildlife makes designing regulations difficult. In order to effectively manage recreational free-flight activities and make them compatible with the conservation of biodiversity in places where sensitive species occur, it is necessary to carry out focused studies to identify the potential gaps in our knowledge (Marion et al. Reference Marion, Davies, Demšar, Irvine, Stephens and Long2020). With such data, the risks and effects of free-flight activities on wildlife in different natural areas can be evaluated, guidelines can be formulated to regulate free-flight and scientific research gaps and evaluation requirements can be identified.
This study set out to review the existing scientific work evaluating the effects of free-flight on wildlife: what information exists, what are the gaps in knowledge and what is needed to help achieve the coexistence of recreational free-flight and wildlife. We present the review in three main parts. To begin with, the review presents an overview of the effect of aircraft on wildlife, which demonstrates that aviation activities have effects on individual animals and also at the population level, being a potential threat to biodiversity. This is followed by a review of the existing data of the effects of free-flight at the taxonomic-group level, paying attention not only to the observed effects, but also to the existing knowledge gaps, as well as the precautionary principles that should be taken into account when carrying out this activity in the presence of these groups of species. Finally, we evaluate the data available in order to suggest measures and guidelines to minimize recreational free-flight impacts on wildlife that could improve their coexistence.
Methodology
The review involved two stages: first, we collated published studies on the effects of free-flight activities on wildlife through a systematic search using Web of Science (n = 5 publications); and second, we supplemented this search with literature retrieved ad hoc through Google Scholar, reference lists in other papers and expert suggestions (n = 12 publications). The systematic search was done from the Web of Science Core Collection using specified English-language search strings (Supplementary Table S1, available online). Based on the search and the review, we retrieved a total of 16 unique publications (9 journal articles, 2 conference papers, 1 MSc thesis, 1 book chapter and 3 ‘other’ publications such as reports; Table 1). One study that includes data related to the effects of free-flight activities on vultures was removed because the article was retracted by the journal. We included studies conducted in the field and one study conducted in captivity because both approaches can provide valuable insights into the effects of free-flight activities on wildlife. The period of publication of the reviewed studies ranged from 1988 to 2013, with greater numbers of publications in the 1990s (n = 6) and 2000s (n = 5) (Table 1).
Table 1. Summary of studies that evaluate the effects of free-flight on wildlife.
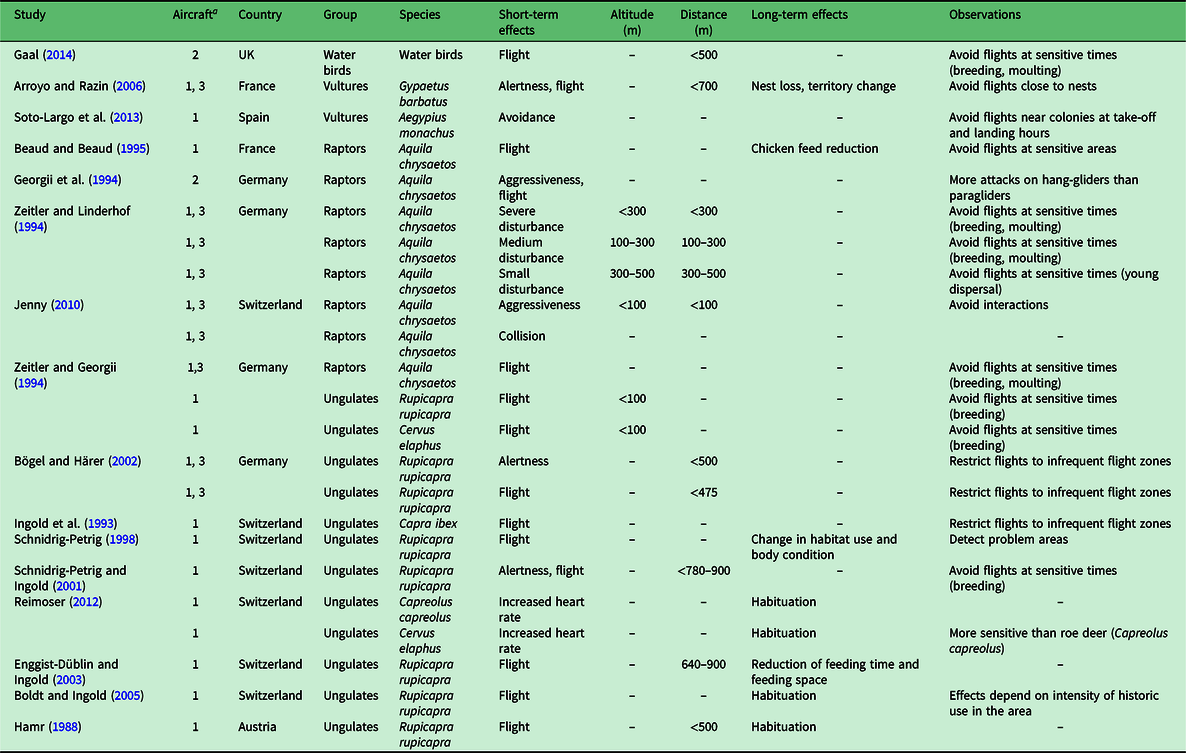
a 1 = paragliders; 2 = paramotors; 3 = hang-gliders.
Effects of aerial activities on biodiversity
In most animal species, the presence of humans generates a reaction that is similar to that of the presence of predators, triggering alert or flight responses after the corresponding risk has been assessed (Frid & Dill Reference Frid and Dill2002). The impacts of these reactions on wildlife can operate at the individual or local population level. At the individual level, modifications in behaviour can result in: increased energy expenditure as animals move to safe areas; reductions in time available for feeding; and loss of body condition (Schnidrig-Petrig Reference Schnidrig-Petrig1998, Enggist-Düblin & Ingold Reference Enggist-Düblin and Ingold2003). They can also cause: increased exposure to predation during movement; harm due to accidents during flight; abandonment of feeding areas; interruption of incubation; abandonment of nests; or reduction of attention given to young (Margalida et al. Reference Margalida, Garcia, Bertran and Heredia2003, Reference Margalida, Benitez, Sánchez-Zapata, Avila, Arenas and Donazar2012, González et al. Reference González, Arroyo, Margalida, Sánchez and Oria2006, Gill Reference Gill2007, Glądalski et al. Reference Glądalski, Bańbura, Kaliński, Markowski, Skwarska and Wawrzyniak2016). Less obvious effects may occur at the physiological level, such as the activation of stress responses and prolonged activation of the hypothalamic–pituitary–adrenal axis. This response to stress is associated with acute and chronic diseases, which produce high levels of glucocorticoids, leading to the depletion of energy reserves and a loss of body condition, which can affect reproductive and population parameters (Thiel Reference Thiel2007, Price Reference Price, Lunney, Munn and Meikle2008, Glądalski et al. Reference Glądalski, Bańbura, Kaliński, Markowski, Skwarska and Wawrzyniak2016).
At the local population level, prolonged disturbance can produce negative effects that compromise the conservation status of the affected populations and affect the presence or absence of a species at the local level (Gill et al. Reference Gill, Sutherland and Watkinson1996, Gill & Sutherland Reference Gill, Sutherland, Gosling and Sutherland1999). The magnitude of such effects is determined by: the character and tolerance of each species to each type of disturbance; the number of individuals in the population that are affected; the frequency and intensity of the disturbance; the activity that the animals are engaged in at the time of the disturbance; the amount of alternative safe habitat available; or the distance to a safe refuge (Batten Reference Batten1977, Gill et al. Reference Gill, Sutherland and Watkinson1996, Enggist-Düblin & Ingold Reference Enggist-Düblin and Ingold2003). High levels of increasing disturbance can drive the simplification of ecological communities towards subsets dominated by the most generalist species and those most tolerant to disturbance, as well as the displacement of the most sensitive species towards less disturbed areas (Fernández-Juridic Reference Fernández-Juridic2002, Bautista et al. Reference Bautista, García, Calmaestra, Palacín, Martín and Morales2003).
However, wildlife can become habituated to some extent to repetitive human activities, especially activities that do not produce real danger. Animals learn that there is nothing to fear and apparently ignore the stimulus, saving the costs of unnecessary flight (Hamr Reference Hamr1988, Price Reference Price, Lunney, Munn and Meikle2008). But there are often species that do not habituate to disturbance, which can suffer continuous and chronic health effects in areas with recurrent disturbance (Fernández-Juridic Reference Fernández-Juridic2002, Thiel Reference Thiel2007, Glądalski et al. Reference Glądalski, Bańbura, Kaliński, Markowski, Skwarska and Wawrzyniak2016). There are also species and specific situations in which partial habituation may occur, in which case the negative physiological effects of human disturbance can become chronic.
Free-flying and derivative activities
The main difference in the possible impact on wildlife between powered and unpowered paragliding and hang-gliding is the noise produced by the engine. The few studies that have evaluated the effects of these two aircraft types have not shown significant differences in their effects on wildlife (Georgii et al. Reference Georgii, Zeitler and Hofer1994, Gaal Reference Gaal2014). However, there are very few studies on the effects of free-flight aircraft on wildlife, particularly of paramotors and powered hang-gliders, and they have been focused on ungulate and raptor species (Table 1).
The effects of free-flight craft on different ecosystems are mainly due to direct disturbance of wildlife behaviour, and also to direct impacts on the ground and vegetation in the jump/take-off and landing areas (Roubault et al. Reference Roubault, Delanoë and Montmollin2003). Studies on wildlife behaviour have mainly focused on the assessment of the direct effects of the presence of paragliders and hang-gliders on behavioural changes in wildlife, the possible effects that these behavioural changes may have on physiology (Schnidrig-Petrig Reference Schnidrig-Petrig1998, Reimoser Reference Reimoser2012) and how the costs of these changes can affect species populations (Enggist-Düblin & Ingold Reference Enggist-Düblin and Ingold2003).
Effects of free-flight at the species-group level
The effects of free-flight and its derivatives on wildlife have been poorly studied and, therefore, the effects of these activities on most species and ecosystems are almost completely unknown. As discussed below, most of these studies have been carried out in the European Alps (Switzerland (n = 7), Germany (n = 4), France (n = 3), Austria (n = 1); Table 1), where the animal communities differ from those found in areas outside the boreal and Alpine ecosystems.
Bird species
Although there is a large body of research on human disturbance on birds (e.g., Verhulst et al. Reference Verhulst, Oosterbeek and Ens2001, Quan et al. Reference Quan, Wen and Yang2002, Blackmer et al. 2004), very few publications consider the disturbance to bird species caused by free-flight activities (raptors (n = 5), vultures (n = 2), water birds (n = 1); Table 1). The data available on the effects of free-flying on waterfowl in wetlands are almost non-existent (Gaal Reference Gaal2014). Similarly, data on the effects on raptors are limited to the bearded vulture (Gypaetus barbatus) on its breeding territories (Arroyo & Razin Reference Arroyo and Razin2006) and the golden eagle (Aquila chrysaetos), mainly on its reaction to the presence of free-flight craft in areas where they share airspace (Georgii et al. Reference Georgii, Zeitler and Hofer1994, Beaud & Beaud Reference Beaud and Beaud1995, Jenny Reference Jenny2010). Data on the effects of free-flight on other species such as passerines or medium- to small-sized birds are limited to a single study (Table 1).
Studies show that the negative effects of free-flying arise from its high noise levels (in the case of powered craft) and the visual similarity of such aircraft to large raptors, which elicits escape or aggression behaviours (Frid & Dill Reference Frid and Dill2002). In a study on the effect of paramotors on waterfowl, Gaal (Reference Gaal2014) suggested that the recurrent presence of paramotors during potentially sensitive periods had a negative effect on waterfowl over the long term. The most sensitive time for possible disturbance of bird species by these aircraft was during the breeding season and moulting period (Verhulst et al. Reference Verhulst, Oosterbeek and Ens2001, Gaal Reference Gaal2014). Birds become flightless during moult and have a lower tolerance to disturbance depending on the species. Sensitivity to human disturbance can result in significant increases in heart rate and stress and increased levels of hormonal stress (Ellenberg et al. Reference Ellenberg, Mattern, Seddon and Jorquera2006). Similarly, during the breeding season, disturbances can reduce the care given to eggs and chicks, so reducing breeding success, and could also increase predation risk (Verhulst et al. Reference Verhulst, Oosterbeek and Ens2001, Bolduc & Guillemette Reference Bolduc and Guillemette2003). Although the use of paramotors and gliders over wetlands is not very common, when they are present and not regulated by temporal or spatial zoning they could be considered highly disruptive (Gaal Reference Gaal2014). Paramotor flights have been observed to disrupt chick-rearing behaviour and to cause avoidance and escape behaviours in raptors (Beaud & Beaud Reference Beaud and Beaud1995). Experimental paragliding flights in areas with no previous activity caused an increase in the heart rate of incubating birds, followed by flight from the area, producing a reaction similar to that caused by the presence of predators (Ingold et al. Reference Ingold, Huber, Neuhaus, Mainini, Marbacher, Schnidrig-Petrig and Zeller1993).
Three main effects of free-flights on large vultures and other raptor species that usually inhabit the cliff areas where these activities mainly take place have been observed (Table 1): collisions with aircraft (Georgii et al. Reference Georgii, Zeitler and Hofer1994, Jenny Reference Jenny2010); behavioural modifications that provoke an aggressive attack reaction (Jenny Reference Jenny2010); and flight from the areas including nest abandonment (Beaud & Beaud Reference Beaud and Beaud1995, Arroyo & Razin Reference Arroyo and Razin2006). As mentioned above, detailed information is lacking on the effects of these activities on birds, including precise information on collisions between birds and these aircraft (Table 1). However, Jenny (Reference Jenny2010) documented 20 cases of collisions between golden eagles and free-flight craft in the European Alps (50% of the collisions had lethal consequences for the eagles), at least two-thirds of which were due to eagle attacks related to territorial defence, the rest being fortuitous or of unknown cause. Most accidents involved sailplanes; only one collision was with a paraglider, another with a helicopter and another with a small aeroplane. However, Georgii et al. (Reference Georgii, Zeitler and Hofer1994) observed that hang-gliders were eight times more likely to be attacked by raptors than paragliders. It seems that the mode of flight and the silhouette of sailplanes and hang-gliders were more likely to trigger the innate aggressive response of eagles to a territorial intrusion. The collisions occurred at altitudes between 1500 and 3200 m, which corresponds to the altitudinal zones occupied by foraging golden eagles in the Alps. The critical approach distance triggering an eagle attack is 100 m for territorial adults, and no relationship was found between the breeding season and attacks (Georgii et al. Reference Georgii, Zeitler and Hofer1994); it appears that the sensitivity of this species, which defends its territory year-round, is not restricted to the breeding season (Jenny Reference Jenny2010). Jenny (Reference Jenny2010) suggested that there may be especially aggressive individuals and that intraspecific variability in aggressive behaviour must be taken into account.
Arroyo and Razin (Reference Arroyo and Razin2006) directly quantified the effects of observable disturbances (alert and escape reactions) on the breeding parameters of bearded vultures in the Pyrenees; disturbances caused by the passage of small aeroplanes and other low-noise craft (including free-flight craft) were tolerated by the birds with no apparent reaction, unless they were closer than 500–700 m to the nest. However, high frequencies of passes by very noisy aircraft (such as helicopters) within a 2km radius of the nest resulted in a high probability of reproductive failure. Territories not subjected to human activities of any kind were on average five times more successful than those where disturbances were frequent. In particular, they found a significant effect of very noisy activities during the breeding season. In raptor species with long reproductive cycles, where laying often occurs in cold seasons, neglect of the nest caused by disturbance may have a greater effect (Arroyo & Razin Reference Arroyo and Razin2006, Margalida et al. Reference Margalida, Benitez, Sánchez-Zapata, Avila, Arenas and Donazar2012). It is also possible that frequently disturbed areas may be more likely to be abandoned in subsequent breeding attempts and that disturbances could therefore have long-term effects on breeding output (Arroyo et al. Reference Arroyo, Lafitte, Sourp, Rousseau, Albert and Heuacker2021). The continued absence of parents from the breeding area, caused by repeated disturbances during the period prior to laying, could also cause instability in pairs. For this reason, Arroyo and Razin (Reference Arroyo and Razin2006) recommended maintaining quiet areas around known nests throughout the year and not only during the reproductive period. Cinereous vultures (Aegypius monachus) have been observed to modify their behaviour in the presence of paragliders, flying over different areas and at different altitudes at times of day when paragliders were active (Soto-Largo et al. Reference Soto-Largo, Prada, Arroyo, García, Blanco, Durán and García2013). In addition, more vultures were observed in the study area on days without paragliding (Soto-Largo et al. Reference Soto-Largo, Prada, Arroyo, García, Blanco, Durán and García2013). It has been suggested that species such as the bearded vulture could habituate to human activity in areas where negative effects on their reproduction had previously been observed. However, the lack of solid evidence and studies on other raptor species means that habituation in raptors cannot be assumed (Duriez et al. Reference Duriez, Margalida, Albert, Arroyo, Couanon and Loustau2020, Arroyo et al. Reference Arroyo, Lafitte, Sourp, Rousseau, Albert and Heuacker2021) and that the possible effects of free-flying on the populations of these species cannot be ignored (Table 1).
The reaction distances of raptors to disturbance caused by free-flight could vary between 300 and 1000 m (Table 1). Due to the lack of local data on the effects of free-flights on raptor species and considering the possibility that gliding practitioners may pass very close to nests on rocky cliffs, distances greater than 1000 m from these areas should be maintained during flights. Therefore, flights within 1000 m of the nests of golden eagles and other cliff-nesting species should be avoided (for both occupied and known historical breeding areas). Furthermore, efforts should be made to minimize possible nuisance in breeding areas. The period of greatest sensitivity around large raptor species’ nests is highly variable and will depend on the phenology of each species, but in general it appears to be highest during breeding – between January and June in the Northern Hemisphere (Margalida et al. Reference Margalida, Garcia, Bertran and Heredia2003, Reference Margalida, González, Sanchez, Oria, Prada and Caldera2007). Depending on the specific characteristics of each area and the species concerned, this date window could be shorter or longer. In any case, all users of powered and unpowered paragliders and hang-gliders, as well as sailplanes, should avoid approaching flying raptors as much as possible, avoid approaching closer than 1000 m and carry out immediate avoidance manoeuvres as soon as an individual, nest or colony of these birds is detected (Zeitler & Linderhof Reference Zeitler and Linderhof1994, Jenny Reference Jenny2010, Gaal Reference Gaal2014). In situations where a fortuitous encounter with these species cannot be avoided due to unpredictable circumstances or ignorance of the location of a sensitive area, avoidance manoeuvres must be carried out and the area left as quickly as possible without approaching within 100 m of the birds, which is the range within which some raptors show aggressive behaviour (Jenny Reference Jenny2010). In the same way, in order to minimize possible encounters with vultures – species that are most commonly encountered during free-flight because they use the same thermal currents at the same time (Soto-Largo et al. Reference Soto-Largo, Prada, Arroyo, García, Blanco, Durán and García2013) – in areas with a known and constant presence of vultures every effort should be made to keep flights above 1200 m in altitude (see Moreno-Opo & Margalida Reference Moreno-Opo and Margalida2017, Arrondo et al. Reference Arrondo, García-Alfonso, Blas, Cortes-Avizanda, De la Riva and Devault2021; Table 2). In this way, the impacts of free-flying on those species known to be sensitive to disturbances caused by free-flight activities can be minimized (Arroyo & Razin Reference Arroyo and Razin2006, Soto-Largo et al. Reference Soto-Largo, Prada, Arroyo, García, Blanco, Durán and García2013; Table 1).
Table 2. Distribution of flight heights (%), in metres above the ground, of the species of nesting vultures in the Iberian Peninsula.
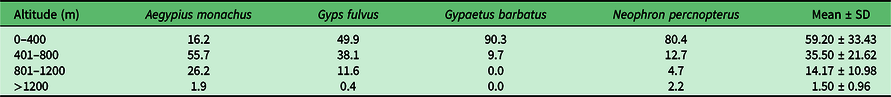
Modified from Moreno–Opo & Margalida (Reference Moreno-Opo and Margalida2017), and original data for Egyptian vultures (Neophron percnopterus) are from Carbonell (Reference Carbonell2020).
The Mediterranean region has a high abundance and diversity of raptor species (del Hoyo Reference del Hoyo2020), and free-flight activities can have a significant impacts on them. The reaction distance to disturbance varies according to species (Ruddock & Whitfield Reference Ruddock and Whitfield2007), so it should be possible either to create specific regulations for each area depending on the species present or to create a common regulation that minimizes the possible effects on all birds that may be present in the areas to be regulated. However, there is a lack of data on the specific reaction distances to paragliders and hang-gliders (powered and unpowered) for most species, so recommended minimum approach distances must be set on the basis of the maximum values observed for other aircraft types or human disturbance for all species (see Ruddock & Whitfield Reference Ruddock and Whitfield2007). Most of the species that may be affected by free-flight activities, such as the cinereous, griffon (Gyps fulvus) and bearded vultures, Spanish imperial eagle (Aquila adalberti), Egyptian vulture (Neophron percnopterus), golden eagle and peregrine falcon (Falco peregrinus), react to human disturbance at distances between 500 and 1000 m (Boeker & Ray Reference Boeker and Ray1971, Arroyo & Razin Reference Arroyo and Razin2006, González et al. Reference González, Arroyo, Margalida, Sánchez and Oria2006, Moreno-Opo & Guil Reference Moreno-Opo and Guil2007, Ruddock & Whitfield Reference Ruddock and Whitfield2007, Zuberogoitia et al. Reference Zuberogoitia, Zabala, Martínez, Martínez and Azkona2008, Margalida et al. Reference Margalida, Benitez, Sánchez-Zapata, Avila, Arenas and Donazar2012). These distances are not specific for these free-flight craft but simply for the presence of human beings (generally on foot), and it is possible that the effects of paragliders and hang-gliders are greater due to their larger size, so in order to minimize their effects based on the precautionary principle, the maximum observed impact distances should be respected. Therefore, the minimum fly-by distances of individuals and in the breeding areas of these species should be greater than 1000 m.
Terrestrial mammals
Studies on the effects of powered and unpowered free-flights on mammal species (n = 9) have focused on mountain ungulate species living in Alpine areas where this activity is usually practiced (Switzerland (n = 6), Germany (n = 2), Austria (n = 1); Table 1). The Alpine ibex (Capra ibex), roe deer (Capreolus capreolus), red deer (Cervus elaphus) and particularly the chamois (Rupicapra rupicapra) have all been studied (Table 1). All of the studies show that paragliding and hang-gliding (unpowered and powered) have an effect on these wild ungulates, without observing any great differences among the species in their response to disturbance except that roe deer seem to be more sensitive than red deer (Reimoser Reference Reimoser2012). The main effects are to change their behaviour (Table 1), primarily increasing vigilance (Schnidrig-Petrig & Ingold Reference Schnidrig-Petrig and Ingold2001, Bögel & Härer Reference Bögel and Härer2002) and flight (Ingold et al. Reference Ingold, Huber, Neuhaus, Mainini, Marbacher, Schnidrig-Petrig and Zeller1993, Zeitler & Georgii Reference Zeitler and Georgii1994, Hamr Reference Hamr1988, Schnidrig-Petrig & Ingold Reference Schnidrig-Petrig and Ingold2001, Boldt & Ingold Reference Boldt and Ingold2005). In addition to direct behavioural changes, changes in habitat use (seeking refuge) and foraging areas have been observed (Schnidrig-Petrig Reference Schnidrig-Petrig1998, Enggist-Düblin & Ingold Reference Enggist-Düblin and Ingold2003), as well as reductions in their time spent feeding (Enggist-Düblin & Ingold Reference Enggist-Düblin and Ingold2003). Furthermore, physical and physiological effects such as increased heart rate and loss of body condition have also been recorded (Schnidrig-Petrig Reference Schnidrig-Petrig1998, Reimoser Reference Reimoser2012). Wild ungulates prone to the effects of free-flight activity suffer the effects immediately, but also over the long term when disturbance is continuous (Zeitler & Georgii Reference Zeitler and Georgii1994, Schnidrig-Petrig Reference Schnidrig-Petrig1998). While habituation by these species to disturbance has been observed in areas with high free-flight frequency (Zeitler & Georgii Reference Zeitler and Georgii1994), the possible effects of free-flight craft on wild ungulate species should be avoided in areas with both high and low frequencies of disturbance (Ingold et al. Reference Ingold, Huber, Neuhaus, Mainini, Marbacher, Schnidrig-Petrig and Zeller1993, Bögel & Härer Reference Bögel and Härer2002). The few studies published on the effects of these aircraft suggest that a minimum flight height of 300 m over areas where these species are present can avoid 75% of the expected disturbance (Bögel & Härer Reference Bögel and Härer2002); increasing the minimum flight height and approach distance to 500 m can remove most of the disturbance to these species (Hamr Reference Hamr1988, Schnidrig-Petrig & Ingold Reference Schnidrig-Petrig and Ingold2001, Bögel & Härer Reference Bögel and Härer2002). Therefore, the minimum recommended flight heights and approach distances should be set at 500 m to minimize the effects of such aircraft on these species. However, the lack of studies on the effects of free-flight on other species means that these values must be treated with caution since there may be negative effects on other mammal species that have not been considered.
Despite the obvious negative effects of free-flight activities on ungulate species in mountainous areas, most studies have observed different responses to the presence of paragliders and hang-gliders according to the frequency and overall number of flights in different areas (Zeitler & Georgii Reference Zeitler and Georgii1994, Schnidrig-Petrig & Ingold Reference Schnidrig-Petrig and Ingold2001). Specifically, studies show that the animals pay less attention to free-flights in those areas where the frequency and number of flights are higher (Boldt & Ingold Reference Boldt and Ingold2005), and they become habituated to them (Hamr Reference Hamr1988, Reimoser Reference Reimoser2012). This variation in effects between different areas can be very important. Zeitler and Georgii (Reference Zeitler and Georgii1994) observed a minimal effect (5% of individuals reacted) in areas with a high frequency of flights, a medium effect (50% of individuals reacted) in area with occasional flights and a strong effect (80% of individuals reacted) in areas in which flights were rare or uncommon. Because of this habituation process, most studies recommend restricting flights to common jumping areas and avoiding flights in new and little-used areas to reduce the effects on wildlife (Ingold et al. Reference Ingold, Huber, Neuhaus, Mainini, Marbacher, Schnidrig-Petrig and Zeller1993, Zeitler & Georgii Reference Zeitler and Georgii1994, Bögel & Härer Reference Bögel and Härer2002).
The observed effects of free-flight activities on terrestrial wildlife in the literature occur mainly in the take-off and landing areas, in cliff areas where cliff-nesting species breed and over high mountain meadows where ungulates feed. Therefore, minimum flight heights and approach distances should be adapted to each location based on the species presence and the specific characteristics of each location in order to minimize the effects on these species (Table 1). It is difficult to reduce or eliminate the effects of free-flight activities on the species of birds and mammals that live in or near take-off and landing areas beyond limiting the opening of new jump/take-off zones. In addition, because mountaintops are often areas of high plant endemicity (Pauli et al. Reference Pauli, Gottfried, Dirnböck, Dullinger, Grabherr, Grabherr, Korner, Thompson and Nagy2003), plants should also be considered.
Other species
Compared with other types of aircraft that have been used to study marine wildlife, free-flight craft have been little used in wildlife research at sea (Roos et al. Reference Roos, Pelletier, Ciccione, Taquet and Hughes2005, Olson Reference Olson2006). However, most of the studies that use remote-controlled helicopters and low-altitude drones to observe cetaceans and other aquatic mammal species found no significant observable effects on the behaviour of these species (Acevedo-Whitehouse et al. Reference Acevedo-Whitehouse, Rocha-Gosselin and Gendron2010, Arona et al. Reference Arona, Dale, Heaslip, Hammill and Johnston2018). However, paragliders and hang-gliders are not generally suited to use over the sea, and their flights are limited to the coastal zone (Roos et al. Reference Roos, Pelletier, Ciccione, Taquet and Hughes2005). In addition, such flights that are made are generally at a high enough altitude to avoid any effects on cetaceans and other marine animals (Roos et al. Reference Roos, Pelletier, Ciccione, Taquet and Hughes2005). However, the lack of studies to evaluate any such possible effects prevents us from judging whether or not overflights by powered and unpowered paragliders and hang-gliders have any effects on marine species, especially cryptic physiological and stress effects (Ditmer et al. Reference Ditmer, Vincent, Werden, Tanner, Laske and Iaizzo2015).
Coexistence between free-flight activity and wildlife
There are very few studies in countries and regions outside the European Alps where free-flight is commonly practiced (Table 1). In-depth studies on the different species groups that may be affected (Table 1), leading to effective regulations agreed between administrations and related stakeholders, are urgently required (Tobajas et al. in press).
Current data recommend the identification of problem areas and the formulation of guidelines that are acceptable to all of the different stakeholder groups, explaining the effects of free-flight activities on wildlife and the measures and restrictions required to mitigate them (Fig. 1 & Appendix S1). This information should then be provided to free-flight clubs in the immediate vicinity of jump/take-off areas and also be posted on the most commonly used specialist websites (Schnidrig-Petrig Reference Schnidrig-Petrig1998, Schnidrig-Petrig & Ingold Reference Schnidrig-Petrig and Ingold2001). The observed effects suggest that free-fliers should only traverse established routes, that the number of free-flight jump/take-off sites should be restricted and that a minimum flight height should be established (Bögel & Härer Reference Bögel and Härer2002). Effective zoning of permitted free-flight areas should be based on: the expected effects on wildlife; specific restrictions on flight height and permitted times of year; and bans or strict restrictions on free-flight in more sensitive areas where flights are currently infrequent (Ingold et al. Reference Ingold, Huber, Neuhaus, Mainini, Marbacher, Schnidrig-Petrig and Zeller1993).
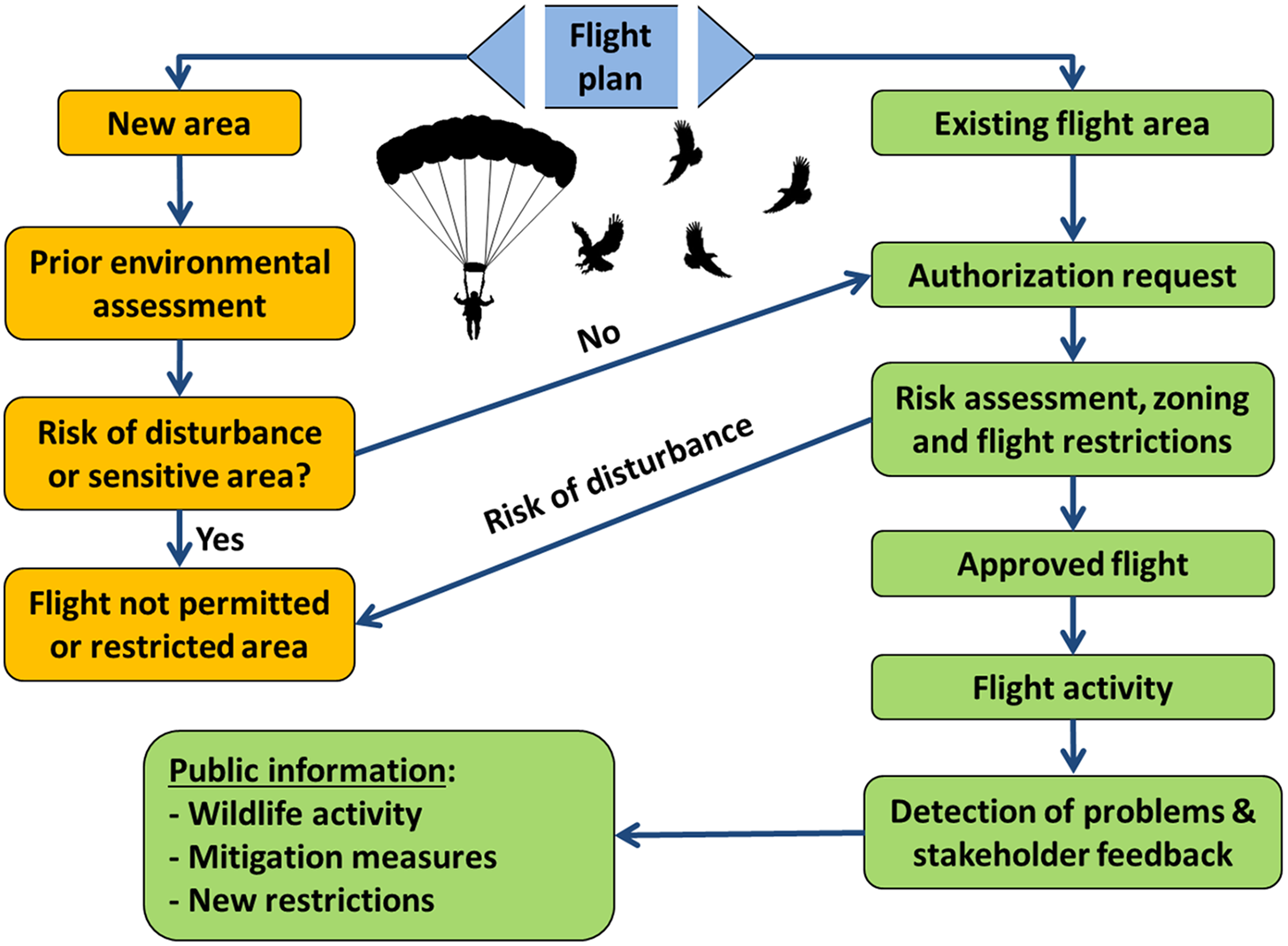
Fig. 1. Guidelines to address the impacts of free–flight activities on wildlife and to foster the coexistence of free–flight activities and wildlife conservation.
In the case of wild ungulates, Bögel and Härer (Reference Bögel and Härer2002) suggest 300 m as a practical minimum height guideline, although they point out that 75% of the total flight distance covered by flights already causing observable disturbance is at this height, so that the most sensitive species would continue to suffer disturbance. They also comment that powered flights should be restricted to rescue flights only and that these flights should be conducted at the highest altitude possible. Other studies have shown disturbance effects from heights less than 500 m, although the impact varies greatly depending on the degree of habituation of the ungulates in the area concerned (Hamr Reference Hamr1988, Schnidrig-Petrig & Ingold Reference Schnidrig-Petrig and Ingold2001). It is also recommended that flights should avoid areas where ungulates are at the end of pregnancy and during the breeding period (April–June) to reduce effects on reproduction (Schnidrig-Petrig & Ingold Reference Schnidrig-Petrig and Ingold2001). In order to minimize the effects on birds of prey, it is necessary to zone and restrict flights both in the breeding areas and in the general vicinity of species sensitive to these disturbances, as well as to manage and regulate the spatiotemporal aspects of flight routes, limiting them to no less than 1000 m from sensitive areas for these species (Arroyo & Razin Reference Arroyo and Razin2006). However, the presumed habituation phenomena observed in raptors and ungulates suggest that an exhaustive assessment of the situation regarding specific populations in frequently used jump/take-off areas is also required. In addition, it is necessary to study the effects of free-flight jump zones on the substrate and vegetation in the jump/take-off areas, because at present very little is known in this respect (Roubault et al. Reference Roubault, Delanoë and Montmollin2003). Most jump/take-off areas are situated on the higher parts of cliffs, which in many cases are home to endemic species or those with small populations. Jump/take-off areas can also be close to breeding areas and territories of large birds of prey that can be affected by the resulting traffic and large numbers of people. Jump/take off areas should therefore be continuously monitored to evaluate their possible impacts on the surrounding wildlife and vegetation.
Free-flight is a largely seasonal leisure pursuit (Davenport Reference Davenport2004). Seasonality of activities can make it easier to protect wintering waterfowl from disturbance and enable planning for the use of these aircraft in certain areas and at particular times of the year. Attention should be given to the sensitivity of the environment and users should be encouraged to care more about wildlife. Zoning and access point management can successfully exclude powered and non-powered paragliders and hang-gliders from sensitive areas. These aircraft can be launched virtually anywhere and the people using them cannot be identified, making them particularly difficult to control. Registration and the display of unique registration numbers would make it possible to identify the people responsible for each flight, which would allow for penalties that would, in turn, reduce the number of incidents and prevent future recurrences. Information brochures explaining the legislative situation and the wildlife conservation issues should be included in free-flight school learning packages. They could contain local and national data and could be downloaded from websites, or paper copies could be given out at induction courses, making users aware of the sensitivity of nearby areas.
Impact studies on wildlife should be mandatory in newly proposed jump/take-off areas (Fig. 1 & Appendix S1). These impact reports should be presented by the club(s) interested in managing these new flight zones and then approved by a competent authority before any such new developments can proceed. It is also recommended that all jumps be recorded to enable the monitoring of disturbances caused by free-flight activities and by those who do not respect the recommendations/regulations in the flight zones concerned and, in turn, to persuade the users of these aircraft not to fly in unauthorized zones and at times where unnecessary disturbance to wildlife would arise.
Conclusion
The data available on the effects of free-flight activities on wildlife are scarce, and this sport activity is potentially affecting numerous species, some of which are endangered (Arroyo & Razin Reference Arroyo and Razin2006, Soto-Largo et al. Reference Soto-Largo, Prada, Arroyo, García, Blanco, Durán and García2013). In addition, the data available are focused on a small group of species from the European Alps; thus, the main part of the potentially affected species around the world could be under considerable negative effects, potentially affecting their conservation. Currently, due to the paucity of data available, the effects of free-flights on wildlife should be considered a gap in biodiversity conservation knowledge. Although articles on this subject are scarce, the problem exists (as evidenced in newspaper reports and media videos) and deserves attention. We urge researchers, institutions and managers to develop studies evaluating the conservation status and the potential effects of these activities on the species beyond the mountain ungulates and raptors in the free-flight areas. A summary of recommendations to improve the coexistence between free-flight and wildlife can be found in Appendix S1.
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.1017/S0376892921000412.
Acknowledgements
The comments of three anonymous reviewers improved a previous version of this article. Saloro SL kindly granted us access to Egyptian vulture GPS data.
Financial support
This work was supported by the Spanish Ministry for Ecological Transition and the Demographic Challenge.
Competing interests
The authors declare none.
Ethical standards
None.






 Open access
Open access