



Introduction
The green mirid bug Apolygus lucorum is a highly destructive omnivorous pest, and in China it inflicts a broad spectrum of damage across an array of economically significant crops, including cotton, vegetables, grains and fruit orchards (Lu et al., Reference Lu, Wu, Jiang, Xia, Li, Feng, Wyckhuys and Guo2010; Tan et al., Reference Tan, Biondi, Desneux and Gao2012). Lu et al. (Reference Lu, Wyckhuys and Wu2024) summarised the integrated mirid management strategies in China's cotton regions and pointed out farmers getting accustomed to using chemical insecticides for mirid control. The frequent usage of insecticides has unavoidably caused resistance in some A. lucorum field populations for recent years (Zhen and Gao, Reference Zhen and Gao2016; Zhen et al., Reference Zhen, Miao, Liang and Gao2016; Zuo et al., Reference Zuo, Yang, Wu and Wu2019; Wang et al., Reference Wang, Lin, Wang, Duan, Li, Zhang, Cao, Yuchi and Wu2020). More studies suggested that cytochrome P450 (CYPs) were involved in multiple insecticide resistance of A. lucorum (Zhen et al., Reference Zhen, Tan, Miao, Wu and Gao2018; Wu et al., Reference Wu, Deng, Jiang, Zhang, Guo, Duan and Wang2019) and other Hemipteran pest (Ding et al., Reference Ding, Wen, Yang, Zhang, Liu, Liu and Han2013; Elzaki et al., Reference Elzaki, Miah, Wu, Zhang, Jiang and Han2017; Wu et al., Reference Wu, Xu, Pan, Gao, Xi, Zhang and Shang2018). For instance, Zhen et al. (Reference Zhen, Tan, Miao, Wu and Gao2018) reported that various cytochrome P450 genes such as six CYP6 genes (CYP6HM1, CYP6HM2, CYP6JB1, CYP6JB2, CYP6JC1, CYP6X2 and CYP395H1) were upregulated in lambda-cyhalothrin-resistant strain of A. lucorum. Wu et al. (Reference Wu, Deng, Jiang, Zhang, Guo, Duan and Wang2019) pointed out that the CYP395G1 overproduction appeared to be related to dinotefuran resistance in A. lucorum. Ding et al. (Reference Ding, Wen, Yang, Zhang, Liu, Liu and Han2013) proved that the induced CYP6AY1 was probably associated with the imidacloprid resistance in Nilaparvata lugensto. Elzaki et al. (Reference Elzaki, Miah, Wu, Zhang, Jiang and Han2017) demonstrated that CYP353D1v2 could metabolise imidacloprid in Laodelphax striatellus, and Wu et al. (Reference Wu, Xu, Pan, Gao, Xi, Zhang and Shang2018) documented that silencing CYP6CY14 significantly enhanced the susceptibility to thiamethoxam in resistant aphids of Aphis gossypii.
As an important enzyme, NADPH-cytochrome P450 reductase (CPR) (one representative of EC 1.6.2.4) existed in all organisms and participated in a series of biochemical reactions catalysed via cytochrome P450 (CYPs), which served as the electron transfer partner from nicotinamide adenine dinucleotide phosphate (NADPH) to the central haeme group in the CYPs (Feyereisen, Reference Feyereisen1999; Paine et al., Reference Paine, Scrutton, Munro, Gutierrez, Roberts, Wolf and Ortiz de Montellano2005). Up to now, the identification of insect CPRs has been confirmed in over 30 species. Among these characterised CPRs, some had been verified to be involved in P450-mediated metabolism resistance in medical or agricultural pests, such as two medical pests the mosquito Anopheles gambiae (Lycett et al., Reference Lycett, McLaughlin, Ranson, Hemingway, Kafatos, Loukeris and Paine2006) and the bed bug C. lectularius (Zhu et al., Reference Zhu, Sams, Moural, Haynes, Potter and Palli2012), and 12 agricultural pests including one Orthoptera pest Locusta migratoria known as the oriental migratory locust (Zhang et al., Reference Zhang, Wang, Liu, Li, Liu, Wu, Ma and Zhang2017), one pest mite Tetranychus cinnabarinus with common name the carmine spider mite (Shi et al., Reference Shi, Zhang, Shen, Xu, Wei, Zhang, Xu and He2015), six Hemiptera pests the rice brown planthopper N. lugens (Liu et al., Reference Liu, Liang, Zhou, Jiang, Zhu, Yu, Zhang, Gurra and Zhu2015a), the small brown planthopper L. striatellus (Zhang et al., Reference Zhang, Wang, Wang, Yao, Guo and Fang2016), the brown citrus aphid Aphis (Toxoptera) citricidus (Kirkaldy) (Jing et al., Reference Jing, Tan, Ding, Dou, Wei and Wang2018), the cotton aphid Ap. gossypii (Tang et al., Reference Tang, Li, He and Ma2023), the pea aphid Acyrthosiphon pisum (Qiao et al., Reference Qiao, Fan, Wu, Bai, Wang, Zhang, Wang and Liu2022), the tobacco whitefly Bemisia tabaci Q (He et al., Reference He, Liang, Liu, Zeng, Wang, Wu, Xie and Zhang2020), one Diptera pest, the oriental fruit fly Bactrocera dorsalis (Huang et al., Reference Huang, Lu, Wang, Wei, Feng, Zhang, Xiao, Dou and Wang2015), and three Lepidoptera pests the cotton bollworm Helicoverpa armigera (Tang et al., Reference Tang, Zhao, Feng, Liu and Qiu2012; Zhao et al., Reference Zhao, Tang, Feng and Qiu2014), the beet armyworm Spodoptera exigua (Zhao et al., Reference Zhao, Feng, Tang and Qiu2015) and the tobacco cutworm Spodoptera litura (Ji et al., Reference Ji, Staehelin, Jiang, Liu, Ma, Su, Zhang and Wang2019; Shi et al., Reference Shi, Li, Dong, Shi, Zhou and Liao2021). However, the characterisation and role of CPR in the resistance development in A. lucorum was scarce.
In our study, the basic characterisation of CPR from A. lucorum (AlCPR) was determined. The quantitative PCR (qPCR) method was utilised to determine the gene expression patterns across various developmental times and body parts. The significantly increased sensitivity against lambda-cyhalothrin was only observed in the lambda-cyhalothrin-resistant A. lucorum strain (LcR) when knockdowning AlCPR. Our study would provide solid foundations for further elucidation of the CPR function in the CYP-mediated lambda-cyhalothrin resistance in A. lucorum.
Materials and methods
Insect cultures
The insect A. lucorum was nurtured on a diet of Phaseolus vulgaris within ventilated plastic containers measuring 20 cm by 15 cm by 10 cm. They were maintained under the conditions as follows: temperature 26 ± 1°C, relative humidity 60 ± 5% and photoperiod 16L: 8D. Additionally, adults were added 10% honey. Two strains, SS (susceptible to lambda-cyhalothrin) and LcR (resistant to lambda-cyhalothrin, with resistance ratio >7.8-fold), were developed in our laboratory, and the same strains with the previous study (Zhen et al., Reference Zhen, Tan, Miao, Wu and Gao2018). The susceptible (SS) strain was sourced from the transgenic Bt cotton fields at the Langfang Experimental Station, located in Hebei Province, China, with geographical coordinates of 39.517966 N latitude and 116.666811 E longitude in year 2008 and then raised in our laboratory without insecticide exposure. The LcR strain was obtained by the same population with lambda-cyhalothrin selection for 11 generations.
RNA isolation and cDNA preparation
Adult specimens of A. lucorum were prepared for total RNA isolation with TRIzol reagent supplied by Invitrogen, Carlsbad, CA as described by the manufacturer's instruction. A NAS-99 spectrophotometer from ACTGene was employed for assessing the RNA integrity and concentration. The synthesis of first-strand complementary DNA (cDNA) for qPCR was facilitated by employing the PrimeScript® RT Reagent Kit with the gDNA Eraser from Takara Bio Inc. (Dalian, China) with 1 μg of the total RNA as template. The first-strand synthesis of cDNA for amplifying cDNA 3′ and 5′ ends was conducted with a SMARTerTM RACE cDNA Amplification Kit (Clontech, Palo Alto, CA, USA).
Full-length AlCPR cDNA amplification
Amplification of the conserved CPR segment was achieved by employing degenerate primers NADPHSP/NADPHASP (table 1). Furthermore, rapid amplification of cDNA end (RACE) was accomplished through a nested PCR approach using two combinations of gene-specific and generic primers (UPM and NUP applied by RACE cDNA Amplification Kit). The cloned AlCPR cDNA fragment was sequenced by Invitrogen (Shanghai, China). Subsequently, the complete sequence was validated through PCR utilizing targeted primers as listed in table 1.
Table 1. Primers designed for cloning, RACE, qRT-PCR, RNAi and housekeeping gene.

Sequence and phylogenetic analysis
ProtParam tool (http://web.expasy.org/protparam/) was used to predict the AlCPR basic physicochemical parameters, such as molecular weight (MW) and isoelectric point (pI). Sequence identification and searching for orthologues were executed through the BLASTp tool (http://www.ncbi.nlm.nih.gov). The prediction of the catalytic residues and binding domains was conducted through the Conserved Domain Search service (http://www.ncbi.nlm.nih.gov/cdd/). ClustalW2 alignment tool available at the EBI website (http://www.ebi.ac.uk/Tools/msa/clustalw2/) was used for homology analysis of AlCPR and multiple other insect CPR homologues. The evolutionary relationships were delineated through a phylogenetic tree with MEGA 6.0 (Tamura et al., Reference Tamura, Stecher, Peterson, Filipski and Kumar2013) using the neighbour-joining approach under bootstrap 1000 replicates.
Real-time quantitative PCR
The primers used for qPCR were listed in table 1. qPCR was proceeded with the ABI 7500 real-time PCR apparatus from Applied Biosystems, and the reaction mixture for each sample comprised 1 μl cDNA, 0.5 μl of each primer, 12.5 μl SYBR Green qPCR SuperMix-UDG and 10.5 μl of H2O, adhering to the protocol outlined in the Invitrogen Platinum SYBR Green qPCR SuperMix-UDG kit. Reaction thermal and time conditions were as follows: UDG incubation at 50℃ for 2 min, initial denaturation at 94°C for 2 min, succeeded by 40 repeated cycles of 94°C for 15 s and 60°C for 30 s. After the above procedure, the dissociation curve from 60 to 95°C was produced for testing the primer specificity. Three independent biological experiments were repeated. The 2−ΔΔCt method as originally described by Livak and Schmittgen (Reference Livak and Schmittgen2001) was applied for the calculation of gene relative expression levels. Reference gene RPL27, based on Luo et al. (Reference Luo, Wang, Cheng, Rong, Guo, Peng and Xu2020) was used for normalisation in our study.
RNAi of AlCPR
The primers carrying T7 RNA polymerase promoter were shown in table 1 to amplify GFP and AlCPR fragments. Purified PCR amplicons served as the template of dsRNA synthesis with the MEGscript RNAi Kit (Ambion, USA). The quality and the concentration of the synthesised dsRNAs were confirmed through agarose gel electrophoresis and NAS-99 spectrophotometer (ACTGene, USA), respectively. The dsRNAs were dissolved in RNase-free water. The A. lucorum adults with 3-day-old were anaesthetised by exposure to carbon dioxide, and immediately put on ice for dsRNA injection. Approximately 1 μg dsAlCPR was administered via injection into the delicate area of the conjunctiva, specifically at the juncture of the prothorax and mesothorax in each A. lucorum adult with a Nanoliter 2000 Injector (WPI, USA). Negative and positive controls were treated with an equivalent RNase-free water and dsGFPs, respectively. Each treatment included three bio-repeats, with 15 adults per repeat. The AlCPR transcription levels were tested for determining RNAi efficiency after 24 h dsRNA injection.
Bioassay after RNAi
The pesticide technical lambda-cyhalothrin with 98% purity from Jiangsu Yangnong Chemical Co., Ltd (Yangzhou, Jiangsu, China) was used in our study and diluted in acetone. After 24 h dsRNA injection, a measured volume of the lambda-cyhalothrin preparation (0.6 μl drop−1) microliters per drop was carefully applied to the thorax region of the mirid bugs employing a PB-600 semi-automatic dropper (PAT, USA). The doses of lambda-cyhalothrin applied on each adult were 9 and 30 ng adult−1 (the dose value adopted LD50 of AL-S and LD20 of AL-R according to Zhen et al., Reference Zhen, Tan, Miao, Wu and Gao2018), respectively. After 24 h lambda-cyhalothrin exposure, the mortality was tested. Three replications were conducted, and in each replication at least 20 adults were dropped by lambda-cyhalothrin solution.
Data analyses
Data statistical evaluation was performed with the GraphPad Prism 5 software. A two-tailed paired t-test, a type of Student's t-test, was implemented for determining the statistical significance of the difference between two groups. To assess the variability across multiple groups, a one-way ANOVA complemented by Tukey's post hoc test was conducted. Statistical significance was determined at the threshold of P ⩽ 0.05.
Results
AlCPR characterisation
The complete cDNA sequence of AlCPR with GenBank accession number: MG678043 possessed an ORF of 2052 bp nucleotides encoding 683 amino acid residues, while the lengths of 5′ UTR and 3′ UTR obtained were 272 and 234 bp nucleotides, respectively. In the 3′ UTR, a poly A was found without a typical polyadenylation signal site (AATAAA). The calculated pI and the estimated MW of AlCPR protein were identified as 5.19 and 76.82 kDa, respectively.
Signal peptide was absent within the deduced protein sequence of AlCPR. However, it was deduced that the AlCPR protein possesses an N-terminal membrane anchoring domain composed of 20 amino acid residues (20F–39L: FGPLDIVLLVALLGVALWWL) and was predicted at the AlCPR N-terminus, which was possibly involved in the orientation between AlCPR and the endoplasmic reticulum. All conserved function binding motifs of AlCPR, including the FMN-, FAD- and NADP-domains, were found in the primary structure of AlCPR (fig. 1). The detailed FAD combining sites in AlCPR was composed of a triplet of amino acids (Arg 460, Tyr 462 and Ser 463) (fig. 1A). Additionally, a pair of FMN connection sites were also discovered at the N-terminus of FMN domain. The catalytic site of AlCPR is composed of key residues including Ser 463, Cys 635, Asp 680 and Try 682 (fig. 1A). The amino acids forming the binding pockets of FMN, FAD and NADP in AlCPR were conserved with CPRs from other known insects by multiple alignments (fig. 1B). Alignment analysis showed that AlCPR shares 69–85% deduced amino acid identity with CPRs from other insects.

Figure 1. Structure of AlCPR. (A) Outline of AlCPR including membrane anchoring motif (red rectangle), conserved binding regions (FMN, blue rectangle; FAD, purple rectangle; NADP, pink rectangle), and catalytic residues. The segment allocated for dsRNA production is indicated. (B) Conserved domain alignment encompassing FMN, FAD and NADP. Binding site residues are shaded. The amino acid sequences of CPR were retrieved from NCBI.
Phylogenetic relationship with CPRs from other insects
Phylogenetic analysis indicated that insect CPRs, when grouped in one branch together, originated from identical insect taxonomic orders, whereas Phthiraptera was observed to group within the Hemiptera branch. AlCPRs fell within the Hemiptera clade and most closed to C. lectularius CPR (ClCPR) (fig. 2), which was in accordance with the sequence alignment result. AlCPR and ClCPR from C. lectularius shared the highest identity with sequence comparison (table S1). Hence, AlCPR and ClCPR were firstly branched into the small group, and then clustered with other Hemiptera CPRs into Hemiptera branch.
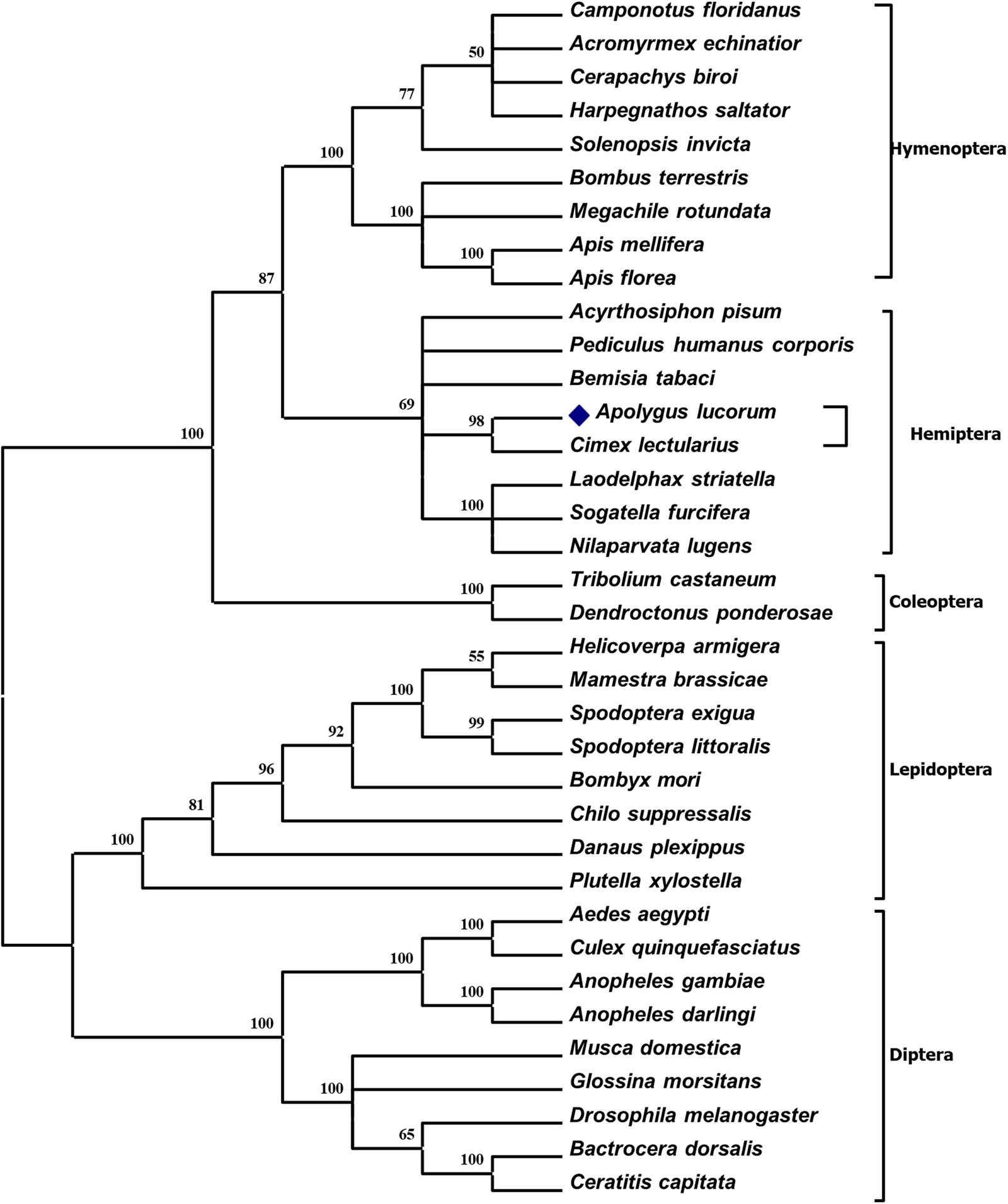
Figure 2. Phylogenetic relationship of AlCPR alongside other arthropod CPRs. The A. lucorum CPR is indicated by a dark blue rhombus. The GenBank numbers utilised to assemble the phylogenetic tree are detailed in table S1.
Expression patterns of AlCPR across the various development phases and tissue
According to the quantitative reverse transcription PCR (qRT-PCR) results, AlCPR transcripts were found to be most abundant in the abdomen, whereas the lowest expression was detected in the head (fig. 3A). The transcription level of AlCPR was expressed during the whole development stages (fig. 3B). Furthermore, AlCPR expression remained relatively stable with no significant fluctuations during the 3rd–5th nymph stage (fig. 3B). The AlCPR mRNA expression levels in nymph stages 1st, 2nd, 3rd, 4th, 5th and adults were 3.35-, 4.23-, 5.87-, 6.13-, 5.48- and 13.29-fold of the transcript level in eggs (fig. 3B).
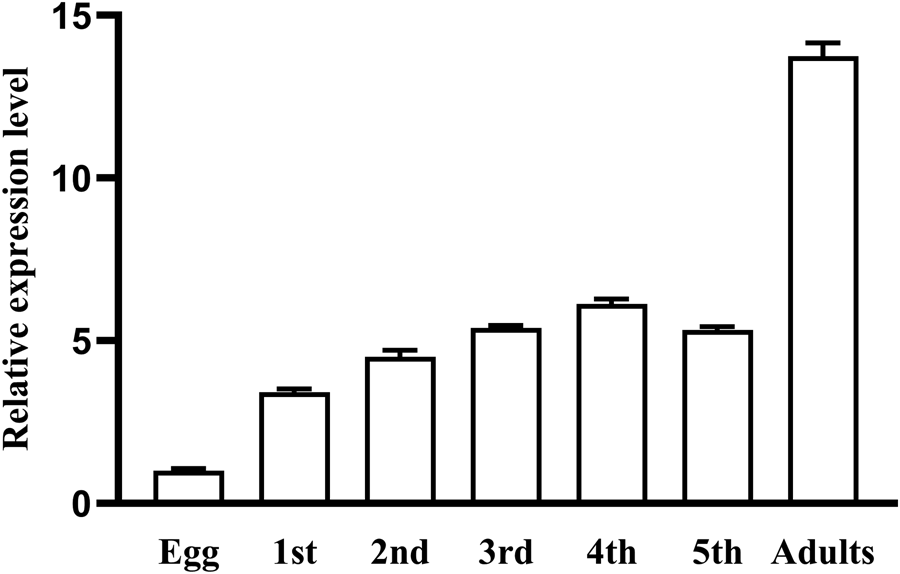
Figure 3. Expression profiles of AlCPR across various tissues in adults (A) and throughout the whole life stages (B).
Expression profiles in LcR and SS strains
Compared to the SS strain, the LcR strain exhibited a pronounced increase (4.99-fold) in AlCPR transcript levels (fig. 4), suggesting AlCPR was most possibly involved in lambda-cyhalothrin resistance in A. lucorum.
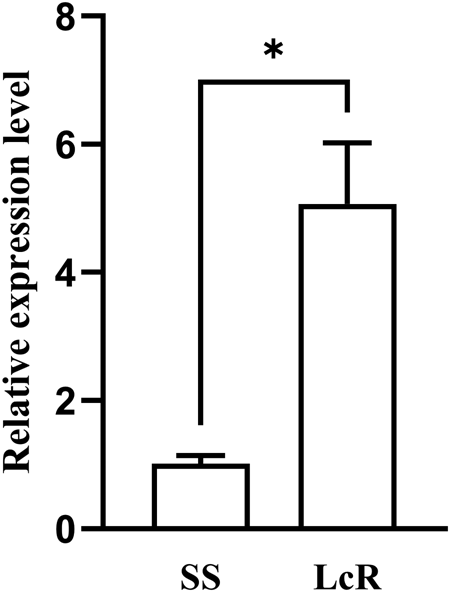
Figure 4. Expression level of AlCPR in SS and LcR strains.
Silencing AlCPR by RNAi
Following a 24 h exposure to dsGFP, the expression levels of AlCPR transcripts remained largely unchanged compared with DEPC-H2O treated group (fig. 5). DsAlCPR injection caused the AlCPR transcripts significantly decrease by 47.5 and 59.0% in the SS strain and the LcR strain, respectively (fig. 5).
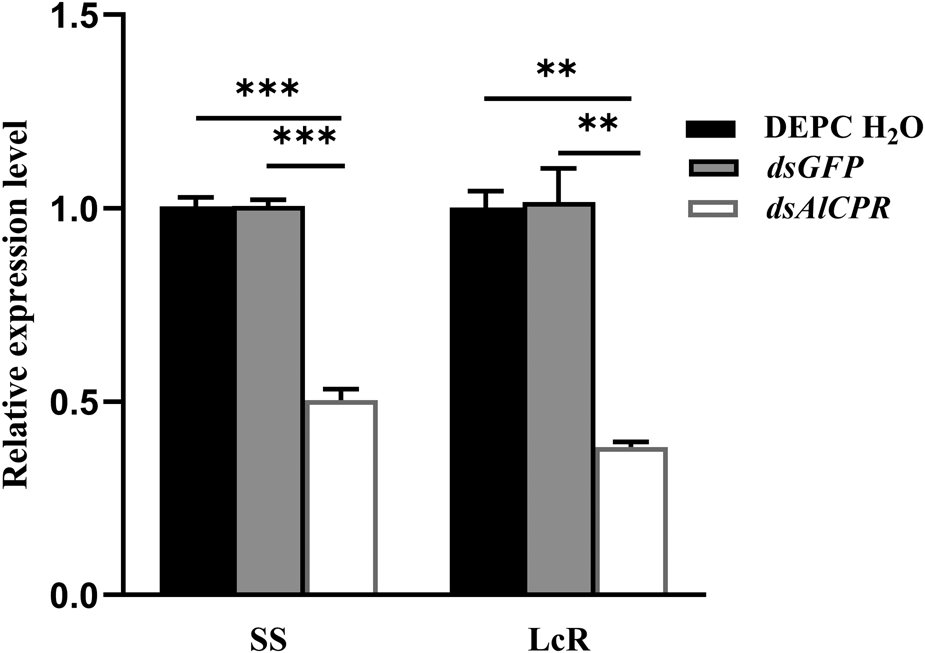
Figure 5. The AlCPR mRNA levels under AlCPR knockdown. SS, lambda-cyhalothrin-susceptible strain; LcR, lambda-cyhalothrin-resistant strain.
AlCPR knockdown increases LcR strain susceptibility to lambda-cyhalothrin
After 24 h dsAlCPR treatment, the susceptibility to lambda-cyhalothrin was significantly elevated in the lambda-cyhalothrin-resistant LcR strain, with the mortality rate enhanced by 23.2% (fig. 6). Conversely, there was no significant increase in the sensitivity of the susceptible SS strain to lambda-cyhalothrin between dsAlCPR treatment and dsGFP control (fig. 6). These results revealed that silencing AlCPR only had a significant impact on the toxicity of lambda-cyhalothrin on resistant A. lucorum.
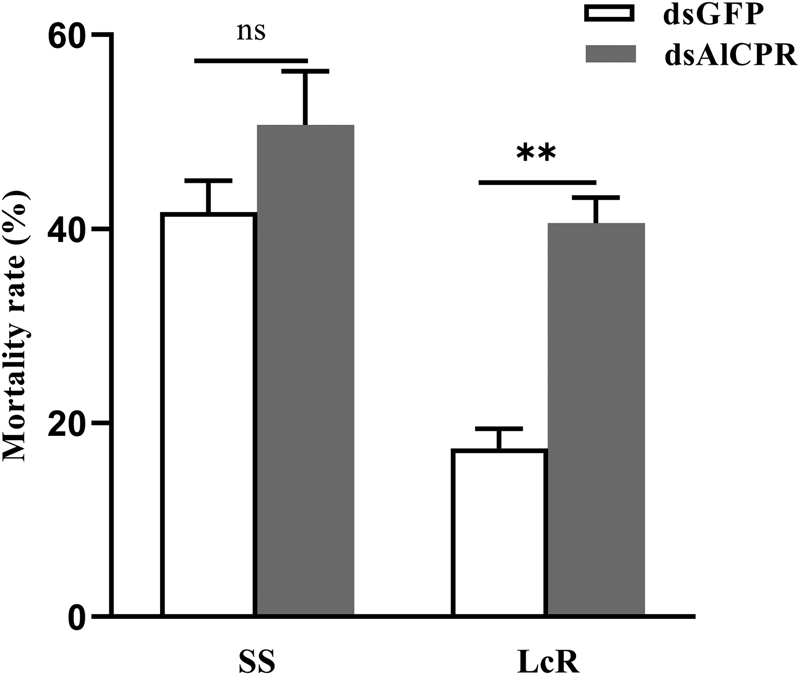
Figure 6. Mortality of A. lucorum adults to lambda-cyhalothrin in the SS strain (A) and LcR strain (B) after silencing AlCPR.
Discussion
In insects, the researches on the pyrethroids resistance mechanisms mainly focused on target site mutations, enhanced metabolic detoxification and epidermal thickening, especially in malaria vectors (Zhu et al., Reference Zhu, Yang, Zhong, Lao, O'Neill, Hong, Zhang and Zhao2020). Among the three major mechanisms of pyrethroid resistance, target-site insensitivity (Soderlund and Knipple, Reference Soderlund and Knipple2003; Scott, Reference Scott2019) and enhanced metabolic detoxification (Panini et al., Reference Panini, Manicardi, Moores and Mazzoni2016) obtained more attention due to their importance (Liu, Reference Liu2015; Smith et al., Reference Smith, Kasai and Scott2016). Zhen and Gao (Reference Zhen and Gao2016) pointed out that the kdr mutation (L1015F) was conferred for the lambda-cyhalothrin resistance in one A. lucorum field population collected from Shandong, Binzhou transgenic Bt cotton. Meanwhile, the metabolic mechanism mediated by overexpressed CYPs was also identified in A. lucorum lambda-cyhalothrin-resistant strain (Zhen et al., Reference Zhen, Tan, Miao, Wu and Gao2018).
CYPs are verified to participate in the insecticide resistance development across numerous insect species (Liu et al., Reference Liu, Li, Gong, Liu and Li2015b; Li et al., Reference Li, Deng and Chen2021; Nauen et al., Reference Nauen, Bass, Feyereisen and Vontas2022). In the P450 system, CPR served as an indispensable electron donor necessary in transferring the electron(s) from NADPH to CYPs, then CYPs catalyse the metabolism of various endogenous substances and xenobiotic chemicals (Pandey et al., Reference Pandey, Kempna, Hofer, Mullis and Fluck2007). Further research on identification and characterisation of insect CPR would be helpful to reveal the detoxification of xenobiotics, especially insecticides mediated by cytochrome P450s via CYPs/CPR system.
In our research, AlCPR, a NADPH-CPR, was obtained from A. lucorum. The expression patterns and potential role in insecticide resistance were characterised. Bioinformatics analysis revealed that a membrane anchor at N-terminus, three types of the conserved binding domains, including FMN, FAD and NADP binding regions, among these motifs, the critical catalytic residues were overall confirmed (fig. 1). The membrane anchor at AlCPR N-terminus is hydrophobic, facilitating the protein molecule anchoring to the endoplasmic membrane in the P450 catalytic cycle and involving the electron transfer via a correct spatial interaction between the CPR protein and CYPs (Wang et al., Reference Wang, Roberts, Paschke, Shea, Masters and Kim1997; Laursen et al., Reference Laursen, Jensen and Møller2011). The FMN and FAD/NADPH binding domains were situated at the N-terminus and near the C-terminus, respectively. These two domains interconnected each other with random coils. Two FMN binding domains were identified in AlCPR including FMN1 and FMN2, suggesting their crucial function on the CYP interaction. Meanwhile, three highly conserved amino acids (R460, Y462 and S463) were found in the FAD binding region of AlCPR. Four conserved amino acid residues (S463, C635, D680 and W682) consisted of AlCPR catalytic centres, which was essential for the hydride transfer circulation. Therefore, the fundamental architecture of CPRs exhibited remarkable conservation across diverse biological groups, which demonstrated the vital role of this enzyme in evolutionary progression (Laursen et al., Reference Laursen, Jensen and Møller2011).
The AlCPR transcript expression pattern was determined throughout the whole life-cycle phases and across a range of tissues. For stage-expression profile, AlCPR transcripts were detected at every developmental stage, with the most abundant at adult stage (fig. 3B), suggesting that AlCPR might be important in the development of A. lucorum and have a crucial role in adults. This expression trend similarly was found in C. lectularius ClCPR (Zhu et al., Reference Zhu, Sams, Moural, Haynes, Potter and Palli2012), T. cinnabarinus (Shi et al., Reference Shi, Zhang, Shen, Xu, Wei, Zhang, Xu and He2015) and L. migratoria (Zhang et al., Reference Zhang, Wang, Liu, Li, Liu, Wu, Ma and Zhang2017). However, the CPR displayed variable expression profiles during insect development stages among species. Generally, protein tissue distribution was usually associated with its role in certain tissues. In insect abdomen part, there were many important tissues that existed in this section, such as metabolism detoxification organs, including midgut, fat body and Malpighian tubule. It is worth noting that the highest AlCPR mRNA level was found in abdomen. This tissue distribution profile was similar to C. lectularius CPR (Zhu et al., Reference Zhu, Sams, Moural, Haynes, Potter and Palli2012) and N. lugens CPR (Liu et al., Reference Liu, Liang, Zhou, Jiang, Zhu, Yu, Zhang, Gurra and Zhu2015a), all potentially indicating CPR may be involved in the xenobiotic metabolism by P450s.
Multiple researches have verified that up-regulation of CPRs in insect is involved in insecticide resistance. The significant enhancement of CPR mRNA level (3.12-fold) was discovered in a fenpropathrin-resistant T. cinnabarinus strain in contrast to a susceptible strain (Shi et al., Reference Shi, Zhang, Shen, Xu, Wei, Zhang, Xu and He2015). The significantly increased mRNA expression level of HaCPR was found in H. armigera midgut from fenvalerate-resistant strain, contrasting with the susceptible strain (Tang et al., Reference Tang, Zhao, Feng, Liu and Qiu2012). In the rice leaffolder, Cnaphalocrocis medinalis, exposed to non-lethal concentrations of pesticides such as abamectin, chlorantraniliprole and chlorpyrifos, the expression of CmCPR was up-regulated (Zhang et al., Reference Zhang, Li, Rao and Liu2018). In the β-cypermethrin-resistant strain of Plutella xylostella, the CPR mRNA expression was significantly enhanced with 12-fold increase in contrast to the β-cypermethrin-susceptible strain (Chen and Zhang, Reference Chen and Zhang2015). Knockdown of CPR increased the susceptibility against permethrin in the mosquito An. gambiae (Lycett et al., Reference Lycett, McLaughlin, Ranson, Hemingway, Kafatos, Loukeris and Paine2006). ClCPR knockdown only strikingly increased the susceptibility of resistant bed bug population against deltamethrin, but not in the susceptible C. lectularius (Zhu et al., Reference Zhu, Sams, Moural, Haynes, Potter and Palli2012). Silencing the NlCPR gene markedly enhanced the susceptibility of N. lugens nymphs against imidacloprid and β-cypermethrin (Liu et al., Reference Liu, Liang, Zhou, Jiang, Zhu, Yu, Zhang, Gurra and Zhu2015a). The fenvalerate susceptibility of H. armigera was remarkably increased with dsCYP6B7 and dsCPR mixture treatment in the HDFR strain (He et al., Reference He, Liang, Liu, Zeng, Wang, Wu, Xie and Zhang2020). In S. litura, the suppression of SlCPR via RNAi led to enhanced phoxim susceptibility (Ji et al., Reference Ji, Staehelin, Jiang, Liu, Ma, Su, Zhang and Wang2019). Similarly, in the two-spotted spider mite, T. urticae, down-regulation of TuCPR reduced the resistance to three acaricides, fenpyroximate, bifenthrin and abamectin (Adesanya et al., Reference Adesanya, Cardenas, Lavine, Walsh, Lavine and Zhu2020). In Sitophilus zeamais, knockdowning SzCPR resulted in the increased sensitivity to terpinen-4-ol (Huang et al., Reference Huang, Liao, Yang, Shi, Xiao and Cao2020). The thiamethoxam and imidacloprid susceptibility was significantly increased when DcCPR silencing in Asian citrus psyllid Diaphorina citri (Yuan et al., Reference Yuan, Jing, Li, Liu, Liu, Liu, Chen, Jiang, Yuan, Dou and Wang2021). In S. litura, knocking down SlCPR significantly increased the susceptibility against indoxacarb in both SS and InRS strains (Shi et al., Reference Shi, Li, Dong, Shi, Zhou and Liao2021). In the pea aphid Ac. pisum, silencing ApCPR caused a significantly increased susceptibility to three contact insecticides, encompassing chlorpyrifos, imidacloprid and betacypermethrin (Qiao et al., Reference Qiao, Fan, Wu, Bai, Wang, Zhang, Wang and Liu2022). Tang et al. (Reference Tang, Li, He and Ma2023) found that the susceptibility of Ap. gossypii against sulfoxaflor increased during silencing of AgCPR. Our findings indicated that the AlCPR expression was notably elevated in the lambda-cyhalothrin-resistant LcR strain, contrasting with the levels observed in the SS strain (fig. 4), suggesting P450 system was possibly associated with the lambda-cyhalothrin resistance development, consistent with the previous conclusion that the overexpression of numerous cytochrome P450 genes was observed in the LcR strain of A. lucorum (Zhen et al., Reference Zhen, Tan, Miao, Wu and Gao2018). In LcR strain, no kdr mutation was found, implying the detoxification metabolism playing main role in the lambda-cyhalothrin-resistant development. Besides, suppression of AlCPR by dsCPR injection could significantly enhance the susceptibility to lambda-cyhalothrin in LcR strain (fig. 6). All these results implied a significant function for CPR in the CYPs-mediated metabolism resistance against lambda-cyhalothrin in LcR strain.
In conclusion, the transcript profile of AlCPR was characterised from A. lucorum. Furthermore, knockdowning AlCPR only enhanced the susceptibility of A. lucorum to lambda-cyhalothrin in the lambda-cyhalothrin-resistant strain (LcR). All these findings indicated that AlCPR was probably involved in the P450-mediated detoxification of lambda-cyhalothrin. In future, the CPR activity assay between SS and LcR strain should be conducted by assessing and comparing the activity of NADPH-dependent cytochrome C reductase. Besides, the heterologous over-expression of AlCPR in Sf9 cells and cytotoxicity assays with lambda-cyhalothrin at different concentrations were executed to uncover the real role of AlCPR in the resistance of A. lucorum to lambda-cyhalothrin.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S0007485324000488
Acknowledgements
The present study was funded by the National Key R&D Program of China (2022YFD1400300). We are also grateful to our supervisor professor Gao Xiwu, whose guidance has been invaluable throughout our research.
Competing interests
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.









