


Consumption of diets high in simple sugar and saturated fat (i.e. the Western diet) is linked to the development of obesity, insulin resistance (IR), and CVD( Reference Cordain, Eaton and Sebastian 1 ). Increased intake of highly energy-dense diets causes lipid accumulation in non-adipose tissue such as the liver, heart and skeletal muscle that leads to metabolic disturbances( Reference Samuel and Shulman 2 – Reference Rector, Thyfault and Uptergrove 5 ). Fat accumulation in the heart results in lipid-induced IR in cardiac myocytes and cardiovascular dysfunction( Reference Dirkx, van Eys and Schwenk 6 , Reference Kok and Brindley 7 ), and fat in the liver can lead to peripheral IR( Reference Hwang, Stein and Barzilai 8 , Reference An, Muoio and Shiota 9 ). With the mitochondria being a major cellular site involved in lipid metabolism and the main source of reactive O2 species (ROS), mitochondrial dysfunction plays a major role in fat deposition and increased oxidative stress which may result to impaired hepatic and cardiac function in high-fat–high sucrose (HFS) diet-induced obesity and IR( Reference Heilbronn, Smith and Ravussin 10 ). Therefore, approaches that reduce detrimental effects on mitochondrial function and ROS generation in these metabolically active tissues may help delay the development of IR.
Although medical advancements in recent years have increased the opportunities for the pharmacological management of IR and type 2 diabetes (T2D), these drugs may have potential side effects. Several drugs, including diabetic treatment and anti-inflammatory drugs, may undermine mitochondrial function and induce mitochondrial toxicity( Reference Chan, Truong and Shangari 11 , Reference Dykens and Will 12 ). Therefore, alternative approaches such as identification of dietary components that prevent or delay IR and hyperglycaemia, may consequently modulate mitochondrial function and associated metabolic stress.
Wheat germ (WG) is an understudied portion of the wheat grain that contains several bioactive compounds, including large amounts of tocopherols, carotenoids, flavonoids and phytosterols, each of which possesses antioxidant properties( Reference Brandolini and Hidalgo 13 ). Traditionally, WG has been mostly ignored as a byproduct of wheat milling. When utilised, it has been most frequently used to produce WG oil or to add bulk to livestock feed( Reference Brandolini and Hidalgo 13 ). However, extracts from fermented WG has been studied for their anti-tumour properties( Reference Boros, Nichelatti and Shoenfeld 14 ). In addition, WG oil showed a dose-dependent inhibitory potential towards acute and chronic inflammation in rats( Reference Mohamed, Ismael and Ibrahim 15 ). In their report, Alessandri et al.( Reference Alessandri, Pignatelli and Loffredo 16 ) concluded that WG oil supplementation, due to its n-3 fatty acid content, exerted anti-atherosclerotic effects by reducing oxidative stress and platelet CD40 ligand expression in hypercholesterolaemic patients. Furthermore, 6-month supplementation with high dose WG (30 g, twice daily), reduced fasting blood glucose and glycosylated Hb in T2D subjects( Reference Haripriya and Premakumari 17 ). Despite the reported health potential of WG and its bioactive components, its impact on mitochondrial function and antioxidant potential in metabolically active tissues such as the liver and heart is unknown. Therefore, using a model of HFS diet-induced obesity, this study evaluated the effects of WG supplementation on markers of IR, mitochondrial oxidative capacity, and innate antioxidant markers in the liver and heart of C57BL/6 mice fed a HFS diet for 12 weeks. We hypothesised that WG supplementation, due to its many bioactive compounds, will delay the development of IR, preserve mitochondrial oxidative capacity and maintain the antioxidant ability of the liver and heart, in mice fed a HFS diet.
Methods
Study design
In all, forty-eight, 6-week-old male C57BL/6 (Charles River Laboratory) mice were housed at the Oklahoma State University (OSU, Stillwater, OK, USA) Laboratory Animal Research facility in a temperature and humidity-controlled environment, maintained on a 12 h light–12 h dark cycle. Mice were acclimated to standard rodent diet (AIN-93M, Harlan Teklad Diets; Envigo) for 1 week. Following acclimation, mice were randomly assigned to four dietary treatment groups (n 12 mice/group): control (C, 10 % fat; AIN-93M), C+10 % wheat germ, HFS (60 % fat), and HFS+10 % WG (HFS+WG) for 12 weeks. The level of fat in the diet was based on our previous study( Reference Lucas, Li and Peterson 18 ), whereas the 10 % WG dose was based on similar studies that investigated other components of the wheat grain in animals( Reference Neyrinck, Possemiers and Druart 19 ) and WG supplementation in humans( Reference Alessandri, Pignatelli and Loffredo 16 , Reference Haripriya and Premakumari 17 ). The 10 % WG dose used in this study is equivalent to approximately 50 g WG when calculated on a dry human diet (w/w) basis, which is within the range of what has been used in other human studies( Reference Alessandri, Pignatelli and Loffredo 16 , Reference Haripriya and Premakumari 17 ).
WG from Oklahoma red winter wheat were obtained from Shawnee Milling Co., analysed for its nutrient composition by NP Analytical Laboratories, and added to the control or HFS diets (10 %, w/w). The WG diets were adjusted to have the same macronutrient composition, Ca and P to the control or HFS diets. Summary of diet compositions are presented in the online Supplementary Table S1. Individual body weights were measured weekly and food intake was recorded three times/week. Deionized water and food were provided ad libitum. The Institutional Animal Care and Use Committee at OSU approved all animal handling and procedures and were strictly followed.
Glucose tolerance test
After 11 weeks of dietary treatment, mice were fasted for 6 hours before undergoing an intraperitoneal glucose tolerance test (GTT). Blood glucose was measured from tail blood (baseline) followed by injection of a 20 % glucose solution (2 g /kg body weight). Blood glucose was measured again at 15, 30, 60 and 120 min post-glucose injection using an AlphaTrak glucometer (Abbott Laboratories).
Necropsy and tissue processing
At the end of the 12-week dietary treatment, mice were feed-deprived for 3 h and anaesthetised with a cocktail of ketamine/xylazine (80 and 8 mg/kg, respectively). Body composition was assessed using a whole-body densitometer (PixiMus; GE Lunar). Serum was processed from blood collected from the carotid artery and stored at −80°C for later analyses. Abdominal fat, heart and liver were collected, weighed, snap-frozen in liquid N2, and stored at −80°C for further analyses. A portion of fresh liver and heart was isolated for mitochondrial function analyses (as detailed below).
Serum measurements
Serum lipids, inflammatory markers, and diabetes markers were determined as described previously( Reference Ojo, El-Rassi and Payton 20 ). In brief, serum concentrations of TAG, NEFA, total cholesterol (TC) and HDL-cholesterol were measured using a Biolis 24i automated analyzer (Carolina Chemistry). Diabetic markers were assessed with the Bio-Plex mouse diabetic markers kit (product no. 171F7001M; gastric inhibitory peptide (GIP), glucagon-like peptide 1 (GLP-1), insulin, leptin, plasminogen activator inhibitor-1 and resistin) using the Bio-Plex MAGPIX Multiplex reader (Bio-Rad Laboratories Inc.) according to the manufacturer’s instructions. IR was estimated utilising the homoeostatic model assessment of IR (HOMA-IR) as described previously( Reference Berglund, Li and Poffenberger 21 ): HOMA-IR=(fasting insulin (µU/ml)×fasting glucose (mg/dl)/405.
Mitochondrial respirometry
Liver and heart samples (60 mg) were homogenised on ice at low speed (Qiagen homogenizer) in 0·5 ml of mitochondrial isolation buffer (210 mm mannitol, 70 mm sucrose, 5 mm HEPES, 1 mm ethylene glycol-bis(β-aminoethyl ether)-N,N,N’,N’-tetraacetic acid (EGTA) and 0·5 % (w/v) fatty acid-free bovine serum albumin, pH 7·2), followed by a series of centrifugation steps as described earlier( Reference Bharadwaj, Tyrrell and Lyles 22 ). Isolated mitochondrial pellets were suspended in mitochondrial assay solution (1X MAS, 70 mm sucrose, 220 mm mannitol, 5 mm MgCl2, 2 mm HEPES, 10 mm KH2PO4, 1 mm EGTA, 0·2 % (w/v) fatty acid-free bovine serum albumin) and protein concentration was determined using Pierce BCA protein assay kit (Thermo Scientific). Mitochondrial pellet was re-suspended in 200 µg/ml mitochondrial assay solution for further analyses. Mitochondrial respiration was measured as VO2 rates (OCR) using the Seahorse XFe 96 flux analyzer (Seahorse Bioscience) following the manufacturer’s instructions. In brief, isolated mitochondria from the liver and heart samples were seeded in triplicates into a ninety-six-well microplate at 4 and 1 μg/well, respectively. For the coupling assay, final concentrations of compounds after port injections were 4 mm ADP (port A), 1·5 µm oligomycin (port B), 4 µm carbonilcyanide p-triflouromethoxyphenylhydrazone (FCCP) (port C) and 4 µm antimycin A (port D). Final concentrations of compounds for the electron flow assay were 2 µm rotenone (port A), 10 mm succinate (port B), 4 µm antimycin A (port C) and 10 mm/100 µm ascorbate/5,10,15,20-tetrakis(2,4,6-trimethylphenyl)porphyrin (TMP) (port D). From these reactions, basal respiration, proton leak, maximal respiratory capacity, complex 1, 2, 4 and 5 OCR were calculated using the adjusted method from previous study( Reference Lay, Sanislav and Annesley 23 ) and as illustrated in the online Supplementary Figure S1. Results are reported as pmol O2/min per µg protein.
Gene expression analyses
Total RNA was isolated from frozen liver and heart samples using Trizol reagent (Thermo Fisher Scientific) following the manufacturer’s protocol. RNA concentration and quality were assessed using a Nanodrop spectrophotometer (Thermo Fisher Scientific) and agarose gel electrophoresis(
Reference Lucas, Li and Peterson
18
). Real time PCR was carried out as previously described(
Reference Ojo, El-Rassi and Payton
20
). In the liver and heart samples, relative abundance of genes involved in mitochondrial biogenesis (PPARγ coactivator (Pgc) 1-α, and Pgc-1β) and innate antioxidant defense (superoxide dismutases (Sod1, Sod2, Sod3)) were assessed using SYBR green chemistry on an ABI 7900HT system (Applied Biosystems). Relative quantitation for each gene was determined using the
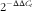 $$2^{{{\minus}\Delta \Delta C_{t} }} $$
method(
Reference Schmittgen and Livak
24
) with cyclophilin B as the invariant control. Oligonucleotide primer sequences used in these experiments are listed in the online Supplemental Table S2. The results are presented relative to the control group.
$$2^{{{\minus}\Delta \Delta C_{t} }} $$
method(
Reference Schmittgen and Livak
24
) with cyclophilin B as the invariant control. Oligonucleotide primer sequences used in these experiments are listed in the online Supplemental Table S2. The results are presented relative to the control group.
Statistical analyses
To calculate power, or the probability of detecting a specified difference, a two-tailed significance level of 0·05 was used. All data are reported as means with their standard errors. Differences between treatments groups were determined using two-way ANOVA with factors of HFS and WG, followed by Tukey’s adjustment. For the weekly body weights and GTT data, repeated-measures ANOVA were also conducted. All statistical analyses were carried out using SAS 9.4 software (SAS Institute). P<0·05 was considered statistically significant.
Results
Body weights, food intake and body composition
Mice had similar body weights at the start of dietary treatment through the 3rd week of feeding (Fig. 1(A)). Starting at the 4th week of dietary treatment, significant differences in weekly body weights were observed until the end of the 12-week study (P<0·0001, Fig. 1(A)). A statistically significant main effect of HFS (P<0·0001) and WG (P=0·008) on weekly body weight was observed. HFS diets increased body weight compared with those receiving the control diets starting at week 5 of treatment (P<0·0001, Fig. 1(A)). At the end of the 12-week treatment, mice fed the control diet had the lowest body weight and the rest of the groups were statistically similar.

Fig. 1 (A) Weekly body weights, (B) food intake and (C) body composition and liver weight of C57BL/6 mice fed either a control (C) or high-fat–high-sucrose (HFS) diet supplemented with 10 % wheat germ (WG) for 12 weeks. Values are means (n 12/group), with standard errors represented by vertical bars. Dietary treatments: C, C+10 % WG, HFS and HFS+10 % WG. P values for significant main effects are shown in each panel. (A): ![]() , C;
, C; ![]() , C+WG;
, C+WG; ![]() , HFS;
, HFS; ![]() , HFS+WG (significant differences between groups: P<0·05, two-way repeated-measures ANOVA). * C is different from HFS and HFS+WG; † C+WG is different from HFS and HFS+WG; ‡ C is different from HFS+WG. (C):
, HFS+WG (significant differences between groups: P<0·05, two-way repeated-measures ANOVA). * C is different from HFS and HFS+WG; † C+WG is different from HFS and HFS+WG; ‡ C is different from HFS+WG. (C): ![]() , C;
, C; ![]() , C+WG;
, C+WG; ![]() , HFS;
, HFS; ![]() , HFS+WG. a,b,c Differences between groups as determined by the post hoc testing, when the interaction HFS×WG was significant (P<0·05, two-way ANOVA).
, HFS+WG. a,b,c Differences between groups as determined by the post hoc testing, when the interaction HFS×WG was significant (P<0·05, two-way ANOVA).
There was a significant main effect of HFS (P=0·003) on food intake (energy content consumed/d) but not WG (Fig. 1(B)). As expected, HFS diet decreased relative amount of lean mass and increased relative amount of fat mass in comparison to the control diets (P<0·0001, Fig 1(C)). The addition of WG also tended to decrease relative lean mass (P=0·056) and increase relative fat mass (P=0·057). Similarly, HFS decreased relative liver weight and increased relative visceral fat mass (P<0·0001) and the addition of WG has no effect on liver weight (Fig. 1(C)). Compared with control, the HFS diet significantly increased relative visceral fat mass by 32 % (P<0·0001). However, WG supplementation in the HFS diet resulted in a 16 % reduction (P=0·006) in relative visceral fat mass compared with HFS (Fig. 1(C)).
Glucose and insulin metabolism
Data from the GTT showed that after 11 weeks of dietary treatment, mice that received the HFS diet had approximately 23 % higher blood glucose (P<0·0001, Fig. 2(A)) compared with control before glucose injection (baseline, 0 min). Mice that received WG in combination with the control or HFS diet had similar baseline glucose to both the control and HFS diets. Following glucose injection, there is a significant interaction effect of HFS and WG (P=0·037) as well as main effect of HFS (P<0·00010) but not WG (P=0·340, Fig. 2(A)). Both HFS and HFS+WG group had significantly higher (P<0·05) blood glucose after 15, 30, 60 and 120 min of glucose injection, compared with both control diets. (Fig. 2(A)). The 19 % decrease in blood glucose at the 120-min time point in HFS+WG compared with HFS was not statistically significant (P=0·256; Fig. 2(A)). In addition, HFS feeding significantly elevated (P<0·0010) glucose total AUC (tAUC) by 48 % compared with control (Fig. 2(B)). The addition of WG to both the control and HFS diets had no effect on tAUC. Despite an 8 % decrease in tAUC in the HFS+WG group compared with HFS, this did not reach significant levels (P=0·242).

Fig. 2 (A) Glucose tolerance test (GTT), (B) GTT total AUC (tAUC), (C) serum insulin concentration, (D) serum gastric inhibitory peptide concentration, (E) serum glucagon-like peptide 1 concentration and (F) homoeostatic model assessment of insulin resistance (HOMA-IR) in C57BL/6 mice fed either a control (C) or high-fat–high-sucrose (HFS) diet supplemented with 10 % wheat germ (WG) for 12 weeks. Values are means with standard errors represented by vertical bars; (A)–(B): n 10–12/group, (C)–(F): n 9/group. P values for significant main effects are shown in each panel. Dietary treatments: C, C+10 % WG, HFS and HFS+10 % WG. (A): ![]() , C;
, C; ![]() , C+WG;
, C+WG; ![]() , HFS;
, HFS; ![]() , HFS+WG (differences between groups at a given time point as determined by the post hoc testing, when the interaction HFS×WG was significant: P<0·05, two-way repeated-measures ANOVA). Δ C is different from HFS, * C is different from HFS and HFS+WG; † C+WG is different from HFS and HFS+WG; ‡ C+WG is different from HFS.
, HFS+WG (differences between groups at a given time point as determined by the post hoc testing, when the interaction HFS×WG was significant: P<0·05, two-way repeated-measures ANOVA). Δ C is different from HFS, * C is different from HFS and HFS+WG; † C+WG is different from HFS and HFS+WG; ‡ C+WG is different from HFS.
Addition of WG to both the control and HFS diets resulted in approximately 50 % reduction in fasting insulin (P=0·0009; Fig. 2(C)). Furthermore, WG also decreased serum concentration of the insulinotropic hormone, GIP (P=0·0003, Fig. 2(D)), and a trend to decrease the other insulinotropic hormone, GLP-1 (−27 %, P=0·088, Fig. 2(E)). Finally, WG but not the HFS diet, significantly reduce (−46 %, P=0·006, Fig. 2(F)) the surrogate measure of in IR, HOMA-IR.
Serum lipids
Analysis of serum TAG showed a significant main effect of HFS (P=0·002) and a trend in the HFS and WG interaction (P=0·063, Fig. 3(A)). WG significantly increased (P=0·010) serum NEFA in C diet group, but not in the groups consuming the HFS diet (Fig. 3(B)). Both the HFS (P<0·0001) and WG (P<0·005) treatments increased serum cholesterol (Fig. 3(C)). Finally, HDL was elevated in the HFS group compared with control (P<0·0001), whereas the addition of WG had no effect (Fig. 3(D)).

Fig. 3 Serum concentrations of (A) TAG, (B) NEFA, (C) total cholesterol and (D) HDL-cholesterol of C57BL/6 mice fed either a control (C) or high-fat–high-sucrose (HFS) diet supplemented with 10 % wheat germ (WG) for 12 weeks. Values are means (n 12/group), with standard errors represented by vertical bars. Dietary treatments: C, C+10 % WG, HFS and HFS+10 % WG. P values for significant main effects are shown in each panel. a,b Differences between groups as determined by the post hoc testing, when the interaction HFS×WG was significant (P<0·05, two-way ANOVA).
Hepatic and cardiac mitochondrial respirometry
Mitochondrial function may be assessed by measuring its OCR to various challenges( Reference Montgomery and Turner 25 ). Mitochondrial respirometry data showed that neither HFS nor WG had an effect on hepatic mitochondrial OCR during basal respiration, maximum respiration, at each complexes of the electron transport chain, and proton leak (Fig. 4(A)). However, in the mitochondria from the myocardium, complex 1 OCR was significantly reduced by both HFS (P=0·011) and WG (P=0·021; Fig. 4(B)). In complex 2, the HFS diet significantly elevated OCR by 45 % (P=0·024) compared with control, but OCR was normalised to the level of control in the HFS+WG group (Fig. 4(B)). Despite the increase in complex 2 OCR by HFS diet, complex 5 OCR was significantly reduced by the HFS diet compared with control (23 % decrease; P=0·002, Fig. 4(B)). The addition of WG to the HFS diet had no statistically significant effect on complex 5 OCR (Fig. 4(B)). Complex 4 OCR tended (P=0·060) to be decreased by the addition of WG.

Fig. 4 Mitochondrial VO2 rates (OCR) in the (A) liver and (B) heart of C57BL/6 mice fed either a control (C) or high-fat–high-sucrose (HFS) diet supplemented with 10 % wheat germ (WG). Mitochondrial OCR was measured in both liver and heart samples using the Seahorse XFe 96 extracellular flux analyzer (see the ‘Methods’ section). Values are means (n 5/group), with standard errors represented by vertical bars. Dietary treatments: C, C+10 % WG, HFS and HFS+10 % WG. P values for significant main effects are shown in each panel. ![]() , C;
, C; ![]() , C+WG;
, C+WG; ![]() , HFS;
, HFS; ![]() , HFS+WG. a,b Differences between groups as determined by the post hoc testing, when the interaction HFS×WG was significant (P<0·05, two-way ANOVA).
, HFS+WG. a,b Differences between groups as determined by the post hoc testing, when the interaction HFS×WG was significant (P<0·05, two-way ANOVA).
Mitochondrial biogenesis and gene expression of innate antioxidants
Similar to the results of hepatic mitochondrial OCR, hepatic gene expression of mitochondrial biogenesis and antioxidants were not significantly affected by dietary treatments, with the exception of WG effects on hepatic Sod2 (Fig. 5(A)). A 38 % increase (P=0·048) in hepatic gene expression of Sod2 was observed with the addition of WG to both the C and HFS diets. In the heart, the HFS diet significantly suppressed the expression of Pgc1α by 49 % (P=0·0034) compared with control. WG decreased Pgc1α when added to the C diet, but increased in the HFS+WG group to the level of both the C and HFS groups (Fig. 5(B)). Pgc1β was not significantly modulated by the dietary treatments (Fig. 5(B)). Furthermore, expression of the mitochondrial isoform of the innate antioxidant gene, Sod2, was significantly lowered (−45 %; P=0·02) by HFS feeding compared with control (Fig. 5(B)). In contrast to the HFS group, HFS+WG diet showed a trend (P=0·11) to counteract the HFS-induced decrease in Sod2 expression (Fig. 5(B)). Finally, neither the cytoplasmic Sod1 nor the extracellular Sod3 was impacted by dietary treatments (Fig. 5(B)).

Fig. 5 Relative gene expression of mitochondrial biogenesis (PPAR-γ coactivators 1-α, 1-β (Pgc1α, Pgc1β)) and antioxidant markers (superoxide dismutases (Sod1, Sod2, Sod3)) in the (A) liver and (B) heart of C57BL/6 mice fed either a control (C) or high-fat–high-sucrose (HFS) diet supplemented with 10 % wheat germ (WG) for 12 weeks. Values are means (n 6/group), with standard errors represented by vertical bars. Dietary treatments: C, C+10 % WG, HFS and HFS+10 % WG. P values for significant main effects are shown in each panel. ![]() , C;
, C; ![]() , C+WG;
, C+WG; ![]() , HFS;
, HFS; ![]() , HFS+WG. a,b Differences between groups as determined by the post hoc testing, when the interaction HFS×WG was significant (P<0·05, two-way ANOVA).
, HFS+WG. a,b Differences between groups as determined by the post hoc testing, when the interaction HFS×WG was significant (P<0·05, two-way ANOVA).
Discussion
We evaluated the effects of WG supplementation on markers of IR, mitochondrial oxidative capacity, and antioxidant markers in the liver and heart of mice fed a HFS diet. The liver and the heart are metabolically active tissues and increased intake of highly energy-dense diets causes lipid accumulation and oxidative stress in these tissues that leads to metabolic disturbances. In this study, WG supplementation reduced IR and stabilized complex 2 mitochondrial metabolism in the heart but not the liver. This study also demonstrates the potential of WG in modulating mitochondrial biogenesis and antioxidant markers.
Few studies have investigated the impact of WG in reducing components of T2D, which is characterized by IR and prolonged hyperglycaemia( Reference Haripriya and Premakumari 17 , Reference Liu, Deseo and Morris 26 ). In diabetic patients, daily intake of WG (60 g) reduced fasting blood glucose and glycosylated Hb levels( Reference Haripriya and Premakumari 17 ). Furthermore, WG possess α-glucosidase inhibitory properties that may be vital in preventing post-prandial hyperglycaemia, a key factor in managing T2D( Reference Liu, Deseo and Morris 26 ). The present study shows from a preventive standpoint that WG supplementation may afford protection against the hyperinsulinaemic response and IR associated with high fat feeding. This effect may be aided by the reduction in visceral fat seen in this study, as accumulation of such fat may predict the development of IR( Reference Miyazaki, Glass and Triplitt 27 ). It should also be noted that WG supplementation lowered serum GIP in both control and HFS mice, accompanied by a reduction in serum insulin and the IR index, HOMA-IR. Interestingly, reports have shown that ablation of the GIP signalling pathway with a GIP antagonist reverses IR in mice( Reference McClean, Irwin and Cassidy 28 , Reference Gault, Irwin and Green 29 ). This suggests that WG supplementation in this study may have modulated insulin metabolism via reduction in the production or signalling of GIP, although this effect needs to be further explored.
HFS diet-induced damage to organs such as the liver and the heart can be due to ROS overproduction by the mitochondrial respiratory chain( Reference Rolo and Palmeira 30 ). ROS overproduction results from the hyperpolarization of the mitochondrial membrane potential, leading to accumulation of electrons which consequently drives the reduction of O2 to the superoxide radical( Reference Rolo and Palmeira 30 ). High fat feeding in this study significantly elevated complex 2 OCR in the cardiac mitochondria compared with control, without increasing complex 5 OCR. WG supplementation normalised the complex 2 OCR to the level of the control. Turner et al. ( Reference Turner, Bruce and Beale 31 ) previously reported similar findings in the skeletal muscle of HFS-fed mice, where excess lipid availability increased the protein expression of mitochondrial complexes, including complex 2. It is noteworthy that HFS feeding in this study significantly elevated serum TAG compared with control. Therefore, our findings suggest that electrons generated in the cardiac mitochondria of the HFS group were not efficiently passed down the subsequent mitochondrial complexes for ATP generation which may potentiate increased ROS production. Subsequently, WG supplementation in the HFS diet showed the ability to counteract the HFS-induced cardiac mitochondrial inefficiency and dysfunction.
Correspondingly, our results indicate that the ability of the heart muscle to cope with the potential ROS generation was impaired as the HFS-fed mice had a significant decrease in cardiac Sod2 gene compared with control. This is in agreement with the study of Sreekumar et al. ( Reference Sreekumar, Unnikrishnan and Fu 32 ) where HFS feeding reduced free radical scavenging enzymes, including Sod1 and Sod2, in rat skeletal muscle. Sod2 is particularly important in preventing mitochondrial oxidative injury as Sod2-deficient mice experienced perinatal mortality due to cardiomyopathy and fatty liver( Reference Li, Huang and Carlson 33 , Reference Lebovitz, Zhang and Vogel 34 ). WG supplementation in the HFS diet showed an antioxidant potential to up-regulate hepatic and cardiac Sod2 transcript. This effect may be due to the richness of WG in tocopherols and bioactive compounds such as carotenoids and flavonoids( Reference Brandolini and Hidalgo 13 ). Although cardiac Sod2 up-regulation by WG in the HFS diet is still statistically similar to HFS, it is probable that the 12-week supplementation in this study was insufficient, as a 36-week supplementation period with antioxidants in HFS diet was reported to up-regulate Sod1 and Sod2 transcript in rat skeletal muscle( Reference Sreekumar, Unnikrishnan and Fu 32 ). Furthermore, it is interesting to note that our data on Sod2 transcript follows a similar pattern to Pgc1α gene expression in the heart of mice fed a HFS diet. HFS feeding significantly reduced Pgc1α compared with control, whereas WG supplementation showed a potential for up-regulation compared with HFS diet. Apart from its mediation of mitochondrial biogenesis, Lu et al. ( Reference Lu, Xu and Hu 35 ) demonstrated that Pgc1α plays a vital role in regulating the expression of mitochondrial antioxidants including Sod2, in the hearts of mice. Therefore, our results suggests that HFS feeding reduces the ability of the heart to cope with ROS generation by suppressing Pgc1α and Sod2 gene expression, but WG supplementation showed the potential to counter this effect.
Furthermore, WG supplementation in HFS-fed mice increased serum total cholesterol. This finding is in contrast to a previous report in humans( Reference Cara, Armand and Borel 36 ). Apart from the physiological differences in mice v. humans, a factor in modulating cholesterol action of WG may be its phytosterol content. Phytosterol-rich WG significantly reduced plasma cholesterol compared with phytosterol-depleted WG in normolipidaemic volunteers( Reference Ostlund, Racette and Stenson 37 ). This effect of phytosterols on cholesterol metabolism was also found to be dose dependent( Reference Racette, Lin and Lefevre 38 ). As we did not assess the phytosterol content of the WG used in this study, it is possible that the phytosterol content of the WG was insufficient to modulate cholesterol metabolism. Limitations of our study include the use of single dose of WG and we did not assess the individual bioactive components present in the WG used in this study. Therefore, future studies should address these limitations as well as explore different sources of WG (i.e. from different wheat varieties), its dose-dependent effects on markers of IR and mitochondrial function, and including other mechanistic pathways it modulates. Furthermore, studies focused on the effects of WG on lipid metabolism should consider analysing the phytosterol content.
In conclusion, this study showed that WG supplementation in HFS-fed C57BL/6 mice reduced IR and improved cardiac, but not liver mitochondrial function. WG also showed a potential to improve the HFS diet-induced reduction in Sod2 and Pgc1a gene expression in the hearts of mice. The bioactive components in WG may be vital in mitigating some of the detrimental effects of HFS diet-induced obesity and IR.
Acknowledgements
The authors would like to acknowledge funding from Oklahoma Agriculture Experiment Station.
Authors contribution were as follows: E. A. L, B. J. S, D. L and B. O designed the research; B. O, A J. S., C. O., L. W., X. G. and S. K. P conducted the research; B. O., E. A. L, and B. J. S. Analysed the data; B. O., A. J. S., E. A. L and B. J. S and wrote the manuscript. E. A. L. had primary responsibility for final content. All authors read and approved the final manuscript.
The authors declare that there are no conflicts of interest.
Supplementary material
For supplementary material/s referred to in this article, please visit https://doi.org/10.1017/S0007114517002082







