


Vitamin A is an essential nutrient for the normal growth of fish( Reference Lall and Lewis-McCrea 1 ). Previous studies from our laboratory have indicated that vitamin A deficiency could decrease weight gain (WG), specific growth rate (SGR) and feed utilisation of Jian carp (Cyprinus carpio var. Jian)( Reference Yang, Zhou and Jiang 2 ). To our knowledge, fish growth is closely related to intestinal health, which is strongly associated with intestinal immune function( Reference Cain and Swan 3 ). A previous study has shown that damages to the intestinal immune system could lead to bacterial translocation and enteritis and even result in high mortality in fish( Reference Urán, Schrama and Rombout 4 ). However, no reports have addressed the effects of vitamin A on immune function in the intestine of fish so far.
In fish, the intestinal immune function is strongly linked to a variety of immune compounds such as lysozyme (LA), acid phosphatase (ACP), antibacterial peptides, complement and cytokines( Reference Gomez, Sunyer and Salinas 5 ). It has been reported that the expression of cytokines could be regulated by the signalling molecule intracellular NF-κB p65 and the inhibitor of κBα (IκBα) in rats( Reference Sun, Feng and Li 6 ) as well as by the signalling molecule p38 mitogen-activated protein kinase (p38MAPK) in humans( Reference Epstein, Docena and MacDonald 7 ). To date, a limited number of studies have explored the effects of vitamin A on animal intestinal immune function, which focus on only cytokines( Reference Lessard, Hutchings and Cave 8 – Reference Dalloul, Lillehoj and Shellem 10 ), secretory IgA and Toll-like receptors (TLR)2, TLR4 and myeloid differentiation primary response gene (88) (MyD88) mRNA levels( Reference Yang, Yuan and Tao 9 ). Nevertheless, no studies have explored the mechanism of the effects of vitamin A on inflammatory cytokines production through NF-κB (including p65/p52 and c-Rel), IκBα, IKK (including IKKα, IKKβ and IKKγ) and p38MAPK signalling in animals. It has been reported that vitamin A can inhibit polysaccharide synthesis in chick embryo cells( Reference Solursh and Meier 11 ). Liu et al.( Reference Liu, Zhang and You 12 ) showed that polysaccharides can suppress the expression of the antibacterial peptide hepcidin in rat liver. Meanwhile, an available study showed that vitamin A could strengthen the phagocytic ability of neutrophils in rats( Reference Twining, Schulte and Wilson 13 ). In humans, neutrophils were able to generate LA for defence against pathogenic micro-organisms( Reference Rasool, Freer and Moreno 14 ). Moreover, in the rat, vitamin A supplementation could decrease glutamate uptake in hippocampus slices( Reference de Oliveira, Rocha and Stertz 15 ). One study showed that high glutamate concentrations could up-regulate cytokine TNF-α mRNA levels in rats( Reference Chaparro-Huerta, Rivera-Cervantes and Flores-Soto 16 ). In addition, Zorn & Sauro demonstrated that vitamin A could activate protein kinase C (PKC) in rat splenic cells( Reference Zorn and Sauro 17 ). Some studies have shown that PKC is an important intermediary in the NF-κB pathway in drosophila cells( Reference Moscat, Rennert and Diaz-Meco 18 ) and in the p38MAPK pathway in chicken chondrocytes( Reference Zhen, Wei and Wu 19 ). These observations implied a connection between vitamin A and innate humoral immune components, inflammatory cytokines as well as the series of NF-κB and p38MAPK signalling molecules in the animal intestine. Thus, further studies to systematically evaluate the effects of vitamin A on animal intestinal immune function will be of great value.
The fish intestine shows regional immune specialisation in different segments. In Atlantic salmon, higher levels of major histocompatibility complex class II( Reference Harstad, Lukacs and Bakke 20 , Reference Løkka, Austbø and Falk 21 ) and T-cell markers( Reference Løkka, Austbø and Falk 21 ) in the hindgut v. the foregut have been reported. Moreover, Rombout & Berg( Reference Rombout and Berg 22 ) reported that in the second gut segment of carp more resident intraepithelial macrophages were found compared with the first segment. To our knowledge, vitamin A (retinol) can be metabolised to retinoic acid (RA) to exert most of its functions including its immunological function in specific subsets of intestinal dendritic cells (DC) and macrophages of animals( Reference Iwata, Hirakiyama and Eshima 23 – Reference Cassani, Villablanca and De Calisto 25 ). Thus, it is valuable to investigate the effects of vitamin A on fish immune function in different intestinal segments.
Grass carp (Ctenopharyngodon idellus) is one of the most important freshwater aquaculture species worldwide( 26 ). At present, the vitamin A requirement of grass carp has been determined in only juveniles, based on the percentage of WG (PWG)( Reference Ming 27 ). However, the nutrient requirements of fish may vary with different growth stages( Reference Campeche, Catharino and Godoy 28 , Reference Guimarães, Lim and Yildirim-Aksoy 29 ) and different physiological indicators( Reference Zhou, Wang and Wang 30 ). Therefore, it is worthy to investigate the dietary vitamin A requirements of grass carp in different growth stages and based on different indicators.
In the present study, we proposed a hypothesis that dietary vitamin A deficiency could impair fish immune function in different intestinal segments. To test this hypothesis, we first systemically investigated the effects of vitamin A on fish enteritis morbidity, innate humoral immune-related components, inflammatory cytokines and the related signalling molecules in the proximal intestine (PI), mid intestine (MI) and distal intestine (DI). Meanwhile, we determined the vitamin A requirements of young grass carp based on different indicators, which may provide a partial basis for formulating the most appropriate feed for grass carp.
Methods
Experimental diet preparation
The experimental diet formulation is given in Table 1. Fishmeal, soyabean protein concentrate and gelatin were offered as dietary protein sources, and soyabean oil was provided as the dietary lipid source. The dietary protein level was fixed at 30 %, which has been shown to be adequate for grass carp as described by Khan et al.( Reference Khan, Jafri and Chadha 31 ). Different concentrations of retinyl acetate (172mg/g) were added to a basal diet constituting six different diets at 0 (un-supplemented control), 0·206, 0·413, 0·619, 0·963 and 1·307 mg/kg diet, and the amount of maize starch was reduced to compensate. The final vitamin A concentrations in each experimental diet were analysed by HPLC assay according to Moren et al.( Reference Moren, Opstad and Berntssen 32 ) to be 0·006 (un-supplemented control), 0·209, 0·416, 0·618, 0·965 and 1·306 mg/kg diet, respectively. All ingredients were mixed, pelleted and stored at −20°C until use, which was similar to the method described by Guimarães et al.( Reference Guimarães, Lim and Yildirim-Aksoy 29 ).
Table 1 Formulation and nutrient content of the basal diet

* Crude protein and crude lipid contents were measured values.
† Available P, n-3 and n-6 contents were calculated according to NRC (2011).
‡ Per kg of vitamin premix (g/kg): cholecalciferol (172mg/g), 0·40; dl-α-tocopherol acetate (50 %), 12·58; menadione (22·9 %), 0·83; cyanocobalamin (1 %), 0·94; d-biotin (2 %), 0·75; folic acid (95 %), 0·42; thiamine nitrate (98 %), 0·11; ascorbyl acetate (95 %), 4·31; niacin (99 %), 2·58; meso-inositol (98 %), 19·39; calcium-d-pantothenate (98 %), 2·56; riboflavin (80 %), 0·63; pyridoxine hydrochloride (98 %), 0·62. All ingredients were diluted with maize starch to 1 kg.
§ Per kg of mineral premix (g/kg): MnSO4.H2O (31·8 % Mn), 1·8900; MgSO4·H2O (15·0 % Mg), 200·0000; FeSO4.H2O (30·0 % Fe), 24·5700; ZnSO4.H2O (34·5 % Zn), 8·2500; CuSO4.5H2O (25·0 % Cu), 0·9600; KI (76·9 % I), 0·0668 g; Na2SeO3 (44·7 % Se), 0·0168. All ingredients were diluted with maize starch to 1 kg.
|| Vitamin A premix: premix was added to obtain graded level of vitamin A and the amount of maize starch was reduced to compensate.
Feeding trial and experimental management
After an acclimation period of 4 weeks as described by Shaik Mohamed et al.( Reference Shaik Mohamed, Sivaram and Christopher Roy 33 ), fish were fed a vitamin A-deficient diet for 2 weeks to diminish the body storage of vitamin A according to Campeche et al.( Reference Campeche, Catharino and Godoy 28 ). Thereafter, a total of 540 fish with an average initial weight of 262·02 (sd 0·45) g were randomly assigned to eighteen experimental cages (1·4 length×1·4 width×1·4 height in metres) in three tanks – that is, six cages with thirty fish per cage in each tank. Each tank was randomly assigned to one of three replicates of the six experimental dietary treatments. Each cage was equipped with a 100-cm-diameter disc in the bottom to collect uneaten food according to the method described by Luo et al.( Reference Luo, Feng and Jiang 34 ). The experimental design included six treatments with three replicates; the respective diet was fed four times per day for 70 d. The fish were fed to apparent satiation in the feeding trial, just as several previous studies have reported( Reference Jiang, Tang and Liu 35 , Reference Feng, Li and Liu 36 ). After feeding for 30 min, the uneaten food was collected, dried and weighed to calculate the feed intake (FI) according to the method of Mundheim et al.( Reference Mundheim, Aksnes and Hope 37 ). Water temperature was set at 28 (sd 2)°C throughout the experiment. The pH and dissolved oxygen levels were maintained at 7·0 (sd 0·2) and not <6·0 mg/l, respectively, and the experimental units were under a natural light and dark cycle, similar to that described by Luo et al.( Reference Luo, Feng and Jiang 34 ). Fish from each cage were weighed at the beginning and at the end of the 70-d feeding trial to determine the PWG, SGR and feed efficiency (FE). The formulae used for calculation are described below.
Challenge test and sample collection
The challenge trial was performed in a similar manner to the trial described by Kuang et al.( Reference Kuang, Xiao and Feng 38 ). After analysing fish growth performance at the end of the feeding trial, fifteen fish from each treatment group were randomly collected with similar body weights and moved into labelled cages according to Liu et al.( Reference Liu, Xu and Ge 39 ). As described by Wu et al.( Reference Wu, Jiang and Liu 40 ), the fish were acclimated in the cages for 5 d. Next, all fish were intraperitoneally injected with Aeromonas hydrophila (College of Veterinary Medicine, Sichuan Agricultural University) at a dose rate of 2·5×108 colony-forming unit/ml per g fish body weight to begin the challenge test. The challenge dose was selected as an appropriate dose that could effectively induce inflammation and consequently enable the investigation on fish reactivity against a threatening disease without causing death according to our preliminary test (data not shown). The challenge test was conducted for 14 d according to Nya & Austin( Reference Nya and Austin 41 ). During the infectious trial, the experimental conditions were the same as the feeding trial. After completion of the challenge experiment, all fish from each treatment were anaesthetised in a benzocaine bath as described by Chen et al.( Reference Chen, Feng and Jiang 42 ). A scoring system was designed to evaluate the severity of fish intestinal inflammation, which was similar to the results of Song et al.( Reference Song, Zhao and Bo 43 ) and Knudsen et al.( Reference Knudsen, Urán and Arnous 44 ). Subsequently, the intestines of the fish were quickly removed and classified according to the position of the turns in the intestine: the PI was anterior to the first turn, the MI was the region located between the first turn and the last turn and the DI was posterior to the last turn( Reference Yaotong and Yongjian 45 , Reference Li, Feng and Jiang 46 ). Thereafter, the intestinal samples were frozen in N2 and stored at −80°C until subsequent analysis of immune parameters.
Intestinal immune parameters assay
The intestinal samples of six fish in each treatment were homogenised on ice in 10 volumes (w/v) of ice-cold physiological saline and centrifuged at 6000 g at 4°C for 20 min; next, the collected supernatant was stored at −20°C as described by Kuang et al.( Reference Kuang, Xiao and Feng 38 ) for subsequent analyses. ACP activity was determined according to the method of Zhao et al.( Reference Zhao, Zhang and Xu 47 ). LA activity was analysed according to Chi et al.( Reference Chi, Liu and Fei 48 ). Complement 3 (C3) and complement 4 (C4) components were determined according to the method of Liu et al.( Reference Liu, Zhao and Brown 49 ).
Real-time quantitative PCR
The procedures of RNA isolation, RT and quantitative real-time PCR were similar to the description from a previous study in our laboratory(
Reference Jiang, Liu and Jiang
50
). In brief, total RNA samples of six fish per treatment were isolated from the intestines using RNAiso Plus (Takara) according to the manufacturer’s instructions, followed by DNAse I treatment. RNA quality and quantity were assessed by agarose gel (1 %) electrophoresis and spectrophotometric (A260:280 nm ratio) analysis, respectively. Next, RNA was reverse transcribed into complementary DNA (cDNA) using the PrimeScript™ RT Reagent Kit (Takara) according to the manufacturer’s instructions. For quantitative real-time PCR, specific primers were designed according to the sequences cloned in our laboratory or the published sequences for grass carp (Table 2). All real-time PCR analyses were performed in six biological replicates, and two technical replicates were analysed for each biological replicate. A total of 15 μl of the PCR reaction system consisted of 7·5 μl of SYBR Premix Ex Taq II (2×), 0·6 μl of forward primers (10 μm), 0·6 μl of reverse primers (10 μm), 2 μl of cDNA and 4·3 μl of DNase-free dH2O. To detect any possible DNA contamination, real-time quantitative PCR was also performed in RNA control samples in the absence of RT during cDNA synthesis. A no-template control of nuclease-free water was also included in each run. All the real-time PCR reactions were performed on a CFX96™ Real-Time PCR Detection System (Bio-Rad, Laboratories, Inc.) using a SYBR® PrimeScript RT-PCR Kit II. According to the results of our preliminary experiment concerning the evaluation of internal control genes, β-actin was used as a reference gene to normalise cDNA loading, which was stably expressed in the present experiment (data not shown). After the final PCR cycle, melting curves were systematically monitored to ensure that only one fragment was amplified. The amplification efficiency of the housekeeping and target genes was calculated using the standard curves of the specific gene generated from 10-fold serial dilutions, as described by Jiang et al.(
Reference Jiang, Hu and Zhang
51
). Pooled cDNA samples of the intestine were used to generate the calibration curves in this study. After verification that the primers amplified with an efficiency of approximately 100 %, the
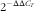 $2^{{{\minus}\Delta \Delta C_{T} }} $
method was used to calculate the relative mRNA levels of all the genes according to Livak & Schmittgen(
Reference Livak and Schmittgen
52
).
$2^{{{\minus}\Delta \Delta C_{T} }} $
method was used to calculate the relative mRNA levels of all the genes according to Livak & Schmittgen(
Reference Livak and Schmittgen
52
).
Table 2 Real-time PCR primer sequences

LEAP-2, liver-expressed antimicrobial peptide 2; TNF-α, tumour necrosis factor α; IFN-γ2, interferon γ2; TGF-β, transforming growth factor β; IκBα, inhibitor of kBα; IKK, IκB kinase; p38MAPK, p38 mitogen-activated protein kinase.
Calculations and statistical analysis
Growth performance parameters were calculated on the basis of the following formulae: growth performance was assessed in terms of PWG, SGR, FI and FE.
 $$\eqalignno{ & {\rm PWG}\,\left( {\rm \%} \right){\rm \,{\equals}\, \cr& 100{\times}{\!}}\left( {{\rm final}\,{\rm weight}\,{\minus}\,{\rm initial}\,{\rm weight}} \right){\rm /initial}\,{\rm weight}{\!}: \cr & {\rm SGR}\,\left( {\rm \%} \right){\rm \,{\equals}\,100{\times}{\!}}\left( {{\rm ln}\,\left( {{\rm final}\,{\rm weight}} \right)\,{\minus}\,{\rm ln}\,\left( {{\rm initial}\,{\rm weight}} \right)} \right){\rm /d{\!}:} \cr & {\rm FE}\,\left( {\rm \,\%\,} \right){\rm \,{\equals}\,100{\times}{\!}}\left( {{\rm final}\,{\rm weight}\,{\minus}\,{\rm initial}\,{\rm weight}} \right){\rm \,/\,FI}{\rm .} $$
$$\eqalignno{ & {\rm PWG}\,\left( {\rm \%} \right){\rm \,{\equals}\, \cr& 100{\times}{\!}}\left( {{\rm final}\,{\rm weight}\,{\minus}\,{\rm initial}\,{\rm weight}} \right){\rm /initial}\,{\rm weight}{\!}: \cr & {\rm SGR}\,\left( {\rm \%} \right){\rm \,{\equals}\,100{\times}{\!}}\left( {{\rm ln}\,\left( {{\rm final}\,{\rm weight}} \right)\,{\minus}\,{\rm ln}\,\left( {{\rm initial}\,{\rm weight}} \right)} \right){\rm /d{\!}:} \cr & {\rm FE}\,\left( {\rm \,\%\,} \right){\rm \,{\equals}\,100{\times}{\!}}\left( {{\rm final}\,{\rm weight}\,{\minus}\,{\rm initial}\,{\rm weight}} \right){\rm \,/\,FI}{\rm .} $$
All data were subjected to a one-way ANOVA followed by Duncan’s multiple-range test to evaluate significant differences among treatments at P<0·05 with SPSS 20.0 (SPSS Inc.), similar to the method of Li et al.( Reference Li, Feng and Jiang 46 ). The requirements of dietary vitamin A based on PWG and intestinal health indicators were estimated by a broken-line or quadratic regression model according to the method of Chen et al.( Reference Chen, Feng and Jiang 42 ).
Results
Growth performance parameters of the fish
The effects of graded levels of dietary vitamin A on fish growth parameters are shown in Table 3. The initial body weight of fish was not significantly different among the groups. The final body weight, PWG, SGR and FI were significantly improved with dietary vitamin A levels up to 0·618 mg/kg and were significantly decreased with higher vitamin A levels. Fish fed the vitamin A 0·618 mg/kg diet showed significantly higher FE than fish fed the vitamin A-deficient control diet and the vitamin A 1·306 mg/kg diet.
Table 3 Initial body weight (IBW, g/fish), final body weight (FBW, g/fish), percentage of weight gain (PWG, %), specific growth rate (SGR, %/d), feed intake (FI, g/fish) and feed efficiency (FE, %) of young grass carp (Ctenopharyngodon idella) fed diets containing graded levels of vitamin A for 10 weeks (Mean values and standard deviations of three replicate groups, with thirty fish in each group)

a,b,c,d,e,f Mean values within a row with unlike superscript letters were significantly different (P<0·05; one-way ANOVA and Duncan’s multiple-range tests).
Enteritis and enteritis morbidity
Fish fed the vitamin A-deficient control diet showed noticeable hypertrophy and haemorrhages after being challenged with A. hydrophila, as shown in Fig. 1. Meanwhile, the effect of vitamin A on fish enteritis morbidity is shown in Fig. 2. The highest enteritis morbidity (65·33 %) was observed in fish fed the vitamin A-deficient control diet, and the morbidity decreased to a minimum (21·33 %) with dietary vitamin A levels up to 0·618mg/kg and then significantly increased with higher vitamin A levels.

Fig. 1 Dietary vitamin A deficiency led to obvious enteritis after infection with Aeromonas hydrophila in young grass carp (Ctenopharyngodon idella). Noticeable hypertrophy and haemorrhages were observed in fish fed vitamin A-deficient diets after infection with A. hydrophila.

Fig. 2 Effects of dietary vitamin A on enteritis morbidity in fish fed diets containing different vitamin A levels for 10 weeks, and then challenged with Aeromonas hydrophila for 14 d. a,b,c,d,e Mean values with different letters were significantly different (P<0·05; one-way ANOVA and Duncan’s multiple-range tests).
Immune parameters in the three intestinal segments of fish
Significant effects of the vitamin A-deficient diet on fish LA and ACP activities as well as the C3 and C4 contents were observed in the PI, MI and DI of fish, as shown in Table 4.
Table 4 Lysozyme activity (LA, U/mg protein), acid phosphatase activity (ACP, U/g protein), complement 3 (C3, mg/g protein) and complement 4 (C4, mg/g protein) contents in the proximal intestine (PI), mid intestine (MI) and distal intestine (DI) of young grass carp (Ctenopharyngodon idella) fed diets containing different vitamin A levels for 10 weeks, and then challenged with Aeromonas hydrophila for 14 days(Mean values and standard deviations of six replicates)

a,b,c,d,e Mean values within a row with unlike superscript letters were significantly different (P<0·05; one-way ANOVA and Duncan’s multiple-range tests).
In the PI, LA and ACP activities as well as C3 and C4 contents were all significantly lower in the vitamin A-deficient control group than in the other vitamin A-supplemented groups. Fish fed the vitamin A 0·618mg/kg diet had significantly higher LA activity than the other groups. ACP activity was significantly increased with dietary vitamin A levels up to 0·416mg/kg and plateaued with higher vitamin A levels. The C3 content gradually increased with dietary vitamin A levels up to 0·618 mg/kg and decreased thereafter. Fish fed the vitamin A 0·416 mg/kg diet had a significantly higher C4 content than the other groups.
In the MI, LA and ACP activities as well as C3 content were all lower in fish fed the control diet compared with fish fed the vitamin A-supplemented diets. The C3 content was significantly increased with dietary vitamin A levels up to 0·416mg/kg and gradually decreased with higher vitamin A levels. The C4 content was significantly increased with dietary vitamin A levels up to 0·416mg/kg and then decreased with further increases in vitamin A levels.
In the DI, fish fed the control diet and the vitamin A 0·209 mg/kg diet showed significantly lower LA activity than fish in the other groups. ACP activity and C3 and C4 contents were all significantly decreased in the control group compared with fish fed the other vitamin A-supplemented diets. Fish fed the vitamin A 0·618, 0·965 and 1·306mg/kg diets had significantly higher ACP activity than the other groups. The C3 content was significantly improved in fish fed the vitamin A 0·618 mg/kg diet and significantly decreased with higher vitamin A levels. Fish fed the vitamin A 0·416 mg/kg diet had significantly higher C4 content than fish in the other groups.
In fact, except for LA activity in the DI as well as the C4 content in the MI, the vitamin A-deficient group showed the lowest values for all parameters in all intestinal sections, which suggested that vitamin A deficiency impaired immune function in fish intestine.
Gene expression in the intestine
mRNA expression of inflammatory cytokines and antimicrobial peptides (β-defensin-1, hepcidin, liver-expressed antimicrobial peptide 2A and liver-expressed antimicrobial peptide 2B) in the proximal intestine, mid intestine and distal intestine
In the PI, as shown in Fig. 3(A1) (pro-inflammatory cytokines) and (A2) (antimicrobial peptides and anti-inflammatory cytokines), TNF-α and IL-15 mRNA levels were all significantly down-regulated in the vitamin A-supplemented diet groups compared with those in the control diet group, except for those in the vitamin A 0·209 mg/kg diet group. The interferon γ2 (IFN-γ2) and IL-8 mRNA levels were significantly down-regulated in the vitamin A-supplemented diet groups compared with those in the control group, except for those in the vitamin A 1·306mg/kg diet group. Fish fed the vitamin A 0·416 and 0·618 mg/kg diets had significantly lower IL-1β mRNA levels than fish in the control group. Compared with fish fed the control diet, the relative mRNA expression of IL-12p35 was significantly down-regulated in fish fed the vitamin A-supplemented diet. In addition, β-defensin-1 and IL-4/13B mRNA levels were all significantly up-regulated in fish fed the vitamin A-supplemented diet compared with those of fish fed the control diet, except for those in the vitamin A 0·209mg/kg diet group. Fish fed the vitamin A 0·416, 0·618 and 0·965mg/kg diets showed significantly higher hepcidin mRNA levels compared with the fish fed the control diet. Compared with fish fed the control diet, the liver-expressed antimicrobial peptide (LEAP)-2A and LEAP-2B mRNA levels were significantly up-regulated, except for those in fish in the vitamin A 1·306 mg/kg diet group. The relative mRNA expressions of IL-11 and IL-4/13A were significantly higher in fish fed the vitamin A 0·618 mg/kg diet than those in fish fed the control diet. However, the relative mRNA expressions of IL-6, IL-17D, IL-12p40, IL-10, transforming growth factor (TGF)-β1 and TGF-β2 were not influenced by vitamin A levels in the PI.

Fig. 3 Effects of dietary vitamin A on relative mRNA levels of pro-inflammatory cytokines TNF-α, IFN-γ2, IL-1β, IL-6, IL-8, IL-15, IL-17D, IL-12p35 and IL-12p40 in the proximal intestine (PI) (A1), mid intestine (MI) (B1) and distal intestine (DI) (C1), and antibacterial peptides β-defensin-1, hepcidin, LEAP-2A and LEAP-2B as well as anti-inflammatory cytokines IL-10, IL-11, TGF-β1, TGF-β2, IL-4/13A and IL-4/13B in the PI (A2), MI (B2) and DI (C2) of fish fed diets containing different vitamin A levels for 10 weeks, and then challenged with Aeromonas hydrophila for 14 d. Expression results were normalised to β-actin expression, which did not change with treatment. Values are means (n 6 fish per treatment), with their standard errors represented by vertical bars. a,b,c,d Mean values were significantly different among the groups (P<0·05; one-way ANOVA and Duncan’s multiple-range tests). IFN-γ2, interferon γ2; LEAP, liver-expressed antimicrobial peptide; TGF, transforming growth factor; VA, vitamin A. ![]() , Control;
, Control; ![]() , VA 0·209;
, VA 0·209; ![]() , VA 0·416;
, VA 0·416; ![]() , VA 0·618;
, VA 0·618; ![]() , VA 0·965;
, VA 0·965; ![]() , VA 1·306 mg/kg diet.
, VA 1·306 mg/kg diet.
In the MI, as shown in Fig. 3(B1) (pro-inflammatory cytokines) and (B2) (antimicrobial peptides and anti-inflammatory cytokines), a significant decrease in the mRNA levels of TNF-α, IFN-γ2 and IL-6 was observed in the vitamin A 0·416, 0·618 and 0·965 mg/kg diet groups compared with those of the control diet. Except for the vitamin A 1·306 mg/kg group, IL-1β and IL-15 mRNA levels were significantly decreased compared with those of the control diet. Fish fed the vitamin A-supplemented diets exhibited a significant decrease in the mRNA levels of IL-8 compared with fish fed the control diet. Fish fed the vitamin A 1209 and 0·618 mg/kg diets had significantly lower IL-17D and IL-12p35 mRNA levels than fish of the control group. In addition, except for the vitamin A 1·306 mg/kg diet group, all the vitamin A-supplemented treatment groups had significantly higher mRNA levels of β-defensin-1 than the control group. Compared with fish in the control group, fish fed the vitamin A 0·618 mg/kg diet exhibited significantly higher hepcidin and TGF-β1 mRNA levels. The LEAP-2A and IL-10 mRNA levels were all significantly up-regulated in fish fed the vitamin A 0·416, 0·618 and 0·965mg/kg diets compared with fish in the control group. The relative mRNA levels of LEAP-2B and TGF-β2 were significantly higher in fish fed the vitamin A 0·416 and 0·618 mg/kg diets compared with fish in the control group. All the vitamin A-supplemented diet groups had significantly higher mRNA levels of IL-11 than the control groups. Compared with fish in the control group, fish fed the vitamin A 0·618, 0·965 and 1·306 mg/kg diets had higher IL-4/13A and IL-4/13B mRNA levels. However, vitamin A had no significant effects on IL-12p40 mRNA levels in the MI.
In the DI, as shown in Fig. 3(C1) (pro-inflammatory cytokines) and (C2) (antimicrobial peptides and anti-inflammatory cytokines), TNF-α mRNA levels were significantly down-regulated in fish fed the vitamin A-supplemented diets compared with fish fed the control diet, except for those in the vitamin A 1·306mg/kg diet group. Compared with fish in the control diet group, IFN-γ2, IL-6 and IL-17D mRNA levels were significantly lower in fish fed the vitamin A 0·416 and 0·618mg/kg diets. Fish fed the vitamin A 0·416, 0·618 mg and 0·965 mg/kg diets had significantly lower IL-1β mRNA levels than fish fed the control diet. IL-8 mRNA level was significantly down-regulated in fish fed the vitamin A 0·618 and 0·965mg/kg diets compared with that of fish fed the control diet. Except for those in the vitamin A 0·209mg/kg diet group, fish in the vitamin A-supplemented groups had significantly higher IL-15 mRNA levels than fish in the control group. Compared with fish in the control group, IL-12p35 mRNA levels were significantly down-regulated in fish fed the vitamin A-supplemented diets. In addition, compared with the mRNA levels of fish in the control diet group, β-defensin-1, TGF-β2 and IL-4/13A were significantly improved in fish in all other groups, except for those in the vitamin A 0·209mg/kg diet group. The mRNA levels of hepcidin were significantly up-regulated in fish fed vitamin A 0·416, 0·618 and 0·965 mg/kg diets compared with fish in the control group. Fish fed the vitamin A 0·416 and 0·618 mg/kg diets exhibited significant increases in the mRNA level of IL-10. Except for those in the vitamin A 1·306 mg/kg diet group, fish fed the vitamin A-supplemented diets had significantly higher IL-11 and TGF-β1 mRNA levels than fish in the control diet group. Compared with the control group, all the vitamin A-supplemented diet groups showed significantly higher IL-4/13B mRNA levels. However, the IL-12p40, LEAP-2A and LEAP-2B mRNA levels in the DI were not affected by dietary vitamin A levels.
mRNA levels of immune-related signalling molecules in the proximal intestine, mid intestine and distal intestine
The effects of dietary vitamin A on NF-κB p65, NF-κB p52, c-Rel, IκBα, IκB kinase α (IKKα), IKKβ, IKKγ and p38MAPK gene expression levels are presented in Fig. 4. To the best of our knowledge, NF-κB p52, NF-κB p65 and c-Rel are three members of the NF-κB family that play a central role in regulating cytokines during inflammation in mammals( Reference Ghosh, May and Kopp 53 ). The results showed that the up-regulation of NF-κB p65 could up-regulate the mRNA levels of pro-inflammatory cytokines IL-8 and IFN-γ2 as well as down-regulate the mRNA levels of anti-inflammatory cytokines IL-10 and TGF-β1 in the intestine of grass carp( Reference Chen, Feng and Jiang 42 ). Meanwhile, Lui et al.( Reference Lui, Lee and Cheng 54 ) reported that the up-regulation of p38MAPK could increase pro-inflammatory cytokines TNF-α and IL-6 in rats.

Fig. 4 Effects of dietary vitamin A on relative mRNA levels of signalling molecules NF-κB p65, NF-κB p52, c-Rel, IκBα, IKKα, IKKβ, KKγ and p38MAPK in the proximal intestine (PI) (A), mid intestine (MI) (B) and distal intestine (DI) (C) of fish fed diets containing different vitamin A levels for 10 weeks, and then challenged with Aeromonas hydrophila for 14 d. Expression results were normalised to β-actin expression, which did not change with treatment. Values are means (n 6 fish per treatment), with their standard errors represented by vertical bars. a,b,c Mean values were significantly different among the groups (P<0·05; one-way ANOVA and Duncan’s multiple-range tests). IκB, inhibitor κB; IKK, IκB kinase; p38MAPK, p38 mitogen-activated protein kinase; VA, vitamin A. ![]() , Control;
, Control; ![]() , VA 0·209;
, VA 0·209; ![]() , VA 0·416;
, VA 0·416; ![]() , VA 0·618;
, VA 0·618; ![]() , VA 0·965;
, VA 0·965; ![]() , VA 1·306 mg/kg diet.
, VA 1·306 mg/kg diet.
In the present study, as shown in Fig. 4(a), in the PI, c-Rel and IKKγ mRNA levels were significantly down-regulated in fish fed the vitamin A-supplemented diets compared with fish fed the control diet, except for those fed the vitamin A 0·209 mg/kg diet. Fish fed the vitamin A 0·618 mg/kg diet had significantly higher IκBα mRNA levels than the control group. Compared with fish in the control group, IKKβ mRNA levels were significantly down-regulated in fish fed the vitamin A 0·416 and 0·618 mg/kg diets. However, NF-κB p65, NF-κB p52, IKKα and p38MAPK mRNA levels in the PI were not significantly affected by dietary vitamin A levels.
In the MI, NF-κB p65 and p38MAPK mRNA levels were all significantly down-regulated in fish fed the vitamin A 0·416, 0·618 and 0·965mg/kg diets compared with fish fed the control diet. Except for those fed the vitamin A 0·209 mg/kg diet, fish fed the vitamin A-supplemented diets had significantly lower c-Rel mRNA levels than fish in the control group. IκBα mRNA levels were significantly enhanced in fish fed the vitamin A 0·618 mg/kg diet compared with fish fed the control diet. Both IKKβ and IKKγ mRNA levels were significantly down-regulated in fish fed the vitamin A-supplemented diets compared with fish fed the control diet, except for those fed the vitamin A 1·306 mg/kg diet. However, NF-κB p52 and IKKα mRNA levels in the MI were not affected by dietary vitamin A levels.
In the DI, compared with fish fed the control diet, NF-κB p65 mRNA levels were significantly down-regulated in fish fed the vitamin A 0·416 and 0·618mg/kg diets. Fish fed the vitamin A-supplemented diets had significantly lower c-Rel mRNA levels than fish in the control group, except for fish fed the vitamin A 0·209 mg/kg diet. IκBα mRNA level was significantly up-regulated in fish fed the vitamin A 0·209, 0·416 and 0·618 mg/kg diets and significantly down-regulated in fish fed the vitamin A 1·306 mg/kg diet compared with fish in the control group. Except for fish fed the vitamin A 1·306mg/kg diet, fish fed the vitamin A-supplemented diets had significant lower IKKβ mRNA levels than fish in the control group. Compared with fish fed the control diet, IKKγ mRNA levels were significantly down-regulated in fish fed the vitamin A 0·209, 0·416 and 0·618mg/kg diets. p38MAPK mRNA levels were significantly lower in fish fed the vitamin A 0·416, 0·618 and 0·965mg/kg diets compared with the control group. However, NF-κB p52 and IKKα mRNA levels in the DI were not affected by dietary vitamin A levels.
Furthermore, the correlation of inflammatory cytokines with NF-κB and p38MAPK signalling in the fish PI, MI and DI are presented in Table 5. The mRNA levels of the pro-inflammatory cytokines TNF-α, IFN-γ2, IL-1β, IL-6, IL-8, IL-15, IL-17D and IL-12p35 were positively related to NF-κB p65, c-Rel and p38MAPK mRNA levels, whereas the mRNA levels of the anti-inflammatory cytokines IL-10, IL-11, TGF-β1, TGF-β2, IL-4/13A and IL-4/13B were negatively related to NF-κB p65, c-Rel and p38MAPK mRNA levels in the different intestinal segments of fish. In addition, NF-κB p65 mRNA levels were negatively related to IκBα mRNA levels in the MI and DI, and c-Rel mRNA levels were negatively related to IκBα mRNA levels in the PI and MI, respectively, whereas IκBα mRNA levels were negatively related to IKKβ mRNA levels in the MI and to IKKγ mRNA levels in the PI, MI and DI.
Table 5 Correlation coefficients of TNF- α, interferon γ2 (IFN- γ2), IL-1 β, IL-6, IL-8, IL-15, IL-17D, IL-12p35, IL-10, IL-11, transforming growth factor (TGF)-β1, TGF-β2, IL-4/13A and IL-4/13B with NF-κ B p65, c-Rel and p38 mitogen-activated protein kinase (p38MAPK); NF-κ B p65 and c-Rel with the inhibitor of kBα (I κ B α); I κ B α with IκB kinase (IKK) β and IKK γ in the proximal intestine (PI), mid intestine (MI) and distal intestine (DI) of fish

Discussion
Vitamin A deficiency decreased fish growth performance and resistance to enteritis
The present study showed that compared with the use of optimum vitamin A levels, vitamin A deficiency remarkably decreased the PWG, SGR, FI and FE of fish (Table 3), suggesting that vitamin A deficiency significantly decreased fish growth performance. On the basis of PWG, the optimum vitamin A requirement of fish was estimated to be 0·664mg/kg diet (y=−115·914x 2+154·054x + 216·598; R 2=0·799, P<0·01) (Fig. 5). Generally, it is a common method to conduct a challenge trial by infecting fish with a widespread pathogenic bacterium after the feeding experiment, which can assess the nutritional enhancement of the disease-resistance abilities of the fish, as several studies have indicated( Reference Peres, Lim and Klesius 55 , Reference Zhang, Li and Xu 56 ). To our knowledge, A. hydrophila is one of the most common pathogen causing bacterial enteritis in freshwater fish( Reference Song, Zhao and Bo 43 , Reference Xu, Ge and Xiong 57 ). Song et al.( Reference Song, Zhao and Bo 43 ) established an enteritis model in grass carp by anal intubation with A. hydrophila. Therefore, after the feeding trial, we infected fish with A. hydrophila to investigate the effect of vitamin A on fish resistance to enteritis in this study. The result showed that vitamin A deficiency caused the highest enteritis morbidity (65·33 %), whereas optimum vitamin A levels significantly decreased enteritis morbidity to 21·33 % (Fig. 2), suggesting that vitamin A deficiency could impair fish resistance to enteritis, whereas optimum vitamin A levels could block this impairment. Meanwhile, the optimum vitamin A level for protecting fish against enteritis morbidity was estimated to be 0·707mg/kg diet (y=78·309x 2−110·695x + 66·485, R 2=0·933, P<0·05). To our knowledge, the ability to resist disease is remarkably connected with immune function in fish( Reference Romarheim, Hetland and Skrede 58 ). Thus, we next investigated the effects of vitamin A on the intestinal immune function of fish.

Fig. 5 Quadratic regression analysis of percentage of weight gain (PWG) in fish fed diets containing different vitamin A levels for 10 weeks.
Vitamin A deficiency impaired fish innate immunity in the proximal intestine, mid intestine and distal intestine
Previous studies have shown that decreased innate humoral immune molecules such as complement (such as C3 and C4)( Reference Nakao, Tsujikura and Ichiki 59 ), LA( Reference Saurabh and Sahoo 60 ) and the antimicrobial peptide hepcidin( Reference Cuesta, Meseguer and Esteban 61 ) could impair fish innate immunity. In the present study, compared with optimum vitamin A levels, dietary vitamin A deficiency significantly decreased LA and ACP activities, C3 and C4 contents and mRNA levels of β-defensin-1 and hepcidin in the PI, MI and DI, as well as LEAP-2A and LEAP-2B in the PI and MI, suggesting that vitamin A deficiency could impair fish intestinal innate immunity. On the basis of ACP activity in the DI, the optimum dietary vitamin A level for the fish was estimated to be 0·722mg/kg (Fig. 6).

Fig. 6 Broken-line analysis of acid phosphatase (ACP) activity in fish fed diets containing different vitamin A levels for 10 weeks, and then challenged with Aeromonas hydrophila for 14 d. DI, distal intestine.
Interestingly, both LEAP-2A and LEAP-2B mRNA levels in the DI were not affected by vitamin A levels. Until now, no information is available regarding the effects of vitamin A on LEAP-2 in animals. The reasons for the negligible effects of vitamin A on LEAP-2A and LEAP-2B mRNA levels in the DI might be partially explained by two factors. First, it could be attributed to the decrease in another antimicrobial peptide in the DI, hepcidin (LEAP-1), caused by vitamin A deficiency. To our knowledge, hepcidin (LEAP-1) shares large structural characteristics with LEAP-2 in mammals( Reference Sang, Ramanathan and Minton 62 ). According to the study of Hocquellet et al.( Reference Hocquellet, Odaert and Cabanne 63 ), the antimicrobial activity of LEAP-2 may not be its main biological function. Thus, hepcidin (LEAP-1) might compensate for LEAP-2 in the defence against pathogens. The present study showed that compared with the optimum vitamin A level vitamin A deficiency significantly decreased the mRNA levels of hepcidin (LEAP-1) in fish DI. Therefore, vitamin A might be operating through hepcidin (LEAP-1) rather than LEAP-2 to defend against pathogens in the DI. Second, it could be partly associated with LEAP-2 expression in different intestinal segments of the fish. One study reported that LEAP-2B mRNA expression in the DI was significantly lower than that in the PI of common carp( Reference Yang, Guo and Li 64 ). These might be a few reasons for the down-regulation of LEAP-2A and LEAP-2B mRNA levels in the PI and MI caused by vitamin A deficiency, whereas both of them were not affected by vitamin A levels in the DI. However, the mechanisms remain unclear. It is well known that fish innate immunity is closely associated with inflammation, which is largely mediated by cytokines( Reference Wang and Secombes 65 ). Therefore, we next examined the effects of vitamin A on intestinal inflammation in the fish.
Vitamin A deficiency aggravated fish intestinal inflammation and was partly connected to the NF-κB canonical signalling pathway and the p38 mitogen-activated protein kinase signalling pathway
Available studies have shown that the inflammatory process is initiated upon the up-regulation of pro-inflammatory cytokines such as IFN-γ, TNF-α, IL-1β and IL-12 and the down-regulation of anti-inflammatory cytokines such as IL-4/13A, IL-4/13B, IL-10 and TGF-β1 in fish( Reference Wang and Secombes 65 ). In the present study, compared with optimum vitamin A levels, vitamin A deficiency significantly up-regulated the mRNA levels of pro-inflammatory cytokines TNF-α, IFN-γ2, IL-1β, IL-8, IL-15 and IL-12p35 and down-regulated the mRNA levels of anti-inflammatory cytokines IL-11, IL-4/13A and IL-4/13B in the PI, MI and DI, indicating that vitamin A deficiency aggravated fish intestinal inflammation. Previous studies have reported that inflammatory cytokines could be regulated by the signalling molecule NF-κB in Jian carp (Cyprinus carpio var. Jian)( Reference Jiang, Shi and Zhou 66 ) and the signalling molecule p38MAPK in rats( Reference Liu, Pan and Liu 67 ). Thus, we next investigated the effect of vitamin A on NF-κB and p38MAPK signalling pathways in the three intestinal segments of fish.
To the best of our knowledge, NF-κB p52, NF-κB p65 and c-Rel are three members of the NF-κB family that play a central role in regulating cytokines during inflammation in mammals( Reference Ghosh, May and Kopp 53 ). The results showed that the up-regulation of NF-κB p65 could up-regulate pro-inflammatory cytokines IL-8 and IFN-γ2 as well as down-regulate the mRNA levels of anti-inflammatory cytokines IL-10 and TGF-β1 in the intestine of grass carp( Reference Chen, Feng and Jiang 42 ). In the present study, compared with the optimum vitamin A levels, vitamin A deficiency significantly up-regulated c-Rel mRNA levels in the PI, MI and DI and NF-κB p65 mRNA levels in the MI and DI, whereas NF-κB p52 mRNA levels were not affected by vitamin A. The analysis showed that the mRNA levels of the pro-inflammatory cytokines TNF-α, IFN-γ2, IL-1β, IL-6, IL-8, IL-15, IL-17D and IL-12p35 were positively correlated with NF-κB p65 or c-Rel mRNA levels, whereas the mRNA levels of the anti-inflammatory cytokines IL-10, IL-11, TGF-β1, TGF-β2, IL-4/13A and IL-4/13B were negatively correlated with NF-κB p65 or c-Rel mRNA levels in the different intestine segments of fish (Table 5), indicating that vitamin A deficiency might enhance NF-κB p65 or c-Rel (rather than NF-κB p52) mRNA levels to aggravate fish intestinal inflammation. The reason for the negligible effect of vitamin A on NF-κB p52 mRNA might be partially associated with the different effects of vitamin A on different NF-κB signalling pathways. To our knowledge, NF-κB signalling can be elicited in two activation pathways: the canonical signalling pathway and the non-canonical signalling pathway. Canonical signalling depends on the activation of IκB kinase (IKKα, IKKβ, IKKγ) and the degradation of the inhibitory protein IκBα, which leads to the translocation of NF-κBs (p65/p50/c-Rel) to the nucleus. In contrast, non-canonical NF-κB activation is dependent on the activation of IKKα (but not IKKβ and IKKγ) to induce the processing of p52/RelB dimers in cancer cells( Reference Bollrath and Greten 68 ). The present study showed that compared with optimum vitamin A levels, vitamin A deficiency significantly down-regulated IκBα mRNA levels and up-regulated IKKβ and IKKγ mRNA levels in the PI, MI and DI. However, IKKα mRNA level was not affected by vitamin A levels. Correlation analysis showed that NF-κB p65 mRNA levels were negatively related to IκBα mRNA levels in the MI and DI, and that c-Rel mRNA levels were negatively related to IκBα mRNA levels in the PI and MI, whereas IκBα mRNA levels were negatively related to IKKβ in the MI and to IKKγ mRNA levels in the PI, MI and DI (Table 5). These results suggested that vitamin A regulated these cytokines might be partly through the NF-κB canonical signalling pathway rather than the non-canonical signalling pathway in the PI, MI and DI. However, the underlying mechanism requires further investigation. Furthermore, Liu et al.( Reference Liu, Pan and Liu 67 ) reported that the up-regulation of p38MAPK could increase the pro-inflammatory cytokines TNF-α and IL-6 in rats. In the present study, compared with the optimum vitamin A levels, vitamin A deficiency caused significant up-regulation of p38MAPK mRNA levels in the MI and DI. Further correlation analysis indicated that the mRNA levels of the pro-inflammatory cytokines TNF-α, IFN-γ2, IL-1β, IL-8, IL-15, IL-17D and IL-12p35 in the MI and DI and IL-6 in the MI were positively correlated with p38MAPK mRNA levels. In addition, the mRNA levels of the anti-inflammatory cytokines IL-10 and IL-11 in the MI and TGF-β1, TGF-β2 and IL-4/13B in the MI and DI were negatively related to p38MAPK mRNA levels (Table 5), suggesting that p38MAPK also participated in regulating cytokine production in the MI and DI of fish.
Interestingly, the effects of vitamin A on inflammation in the PI showed some differences compared with the MI and DI in this study. Our results showed that compared with optimum vitamin A levels, vitamin A deficiency caused significant up-regulation in the mRNA levels of the pro-inflammatory cytokines IL-6 and IL-17D and the down-regulation of the mRNA levels of the anti-inflammatory cytokines IL-10, TGF-β1 and TGF-β2 in the MI and DI, whereas all these cytokines were not affected by dietary vitamin A levels in the PI. The reason might be partly associated with NF-κB p65 and p38MAPK signalling pathways. The present study showed that vitamin A deficiency could significantly up-regulate NF-κB p65 and p38MAPK mRNA in the MI and DI. Correlation analysis suggested that IL-6, IL-17D, IL-10, TGF-β1 and TGF-β2 mRNA levels in the MI and DI were significantly correlated with NF-κB p65 or p38MAPK mRNA levels (Table 5), suggesting that IL-6, IL-17D, IL-10, TGF-β1 and TGF-β2 mRNA levels could be regulated by NF-κB p65 or p38MAPK in the MI and DI. However, both NF-κBp65 and p38MAPK were not affected by vitamin A levels in the PI, which could be a potential reason for the negligible effects of dietary vitamin A on IL-6, IL-17D, IL-10, TGF-β1 and TGF-β2 mRNA levels in the PI. Furthermore, the reason for the non-significant impact of vitamin A on NF-κB p65 and p38MAPK mRNA levels in the PI may be partly associated with the different effects of vitamin A on the different intestinal segments in fish. In general, specific subsets of intestinal DC and macrophages could constitutively express RA-synthesising enzymes, in which vitamin A (retinol) could be metabolised to RA to exert most of its functions including immunological function in animals( Reference Iwata, Hirakiyama and Eshima 23 , Reference Manicassamy and Pulendran 24 ). To the best of our knowledge, the PI is the main nutrient-absorbing region and contains typically lower numbers of bacteria, which showed a relatively weaker inflammatory response than the posterior intestine in teleosts( Reference Marel, Pröpsting and Battermann 69 ). Thus, vitamin A may play a greater immune function in the posterior intestine than in the PI. This might be a part of the reason why vitamin A did not affect NF-κB p65 and p38MAPK mRNA levels in the PI. However, the underlying mechanisms deserve further investigation.
Comparison of the optimal vitamin A levels depending on different indicators
According to our study, the optimum dietary vitamin A level was effective in promoting growth and enhancing intestinal immune function in fish, which clearly varied with different indicators. The optimum vitamin A levels for PWG and protection against enteritis morbidity in grass carp (262·02–996·67 g) were estimated to be 0·664 and 0·707mg/kg in the diet, respectively. In addition, based on ACP activity, the optimum vitamin A levels were estimated to be 0·722 mg/kg in the diet. Apparently, the intestinal immune requirements of fish were slightly higher than the growth requirement, indicating that more vitamin A might be required for the development of fish intestinal immune response and to enhance disease resistance. The results were similar to Zhou et al.( Reference Zhou, Wang and Wang 30 ), who observed that the requirement of vitamin E based on plasma LA activity was higher than the requirement based on WG in the cobia (Rachycentron canadum).
Conclusion
The results of this study demonstrated for the first time that vitamin A deficiency could decrease fish growth, resistance to enteritis and immune function in the PI, MI and DI, and different immune responses in the PI, MI and DI were mediated at least in part by the NF-κB canonical signalling pathway rather than the NF-κB non-canonical signalling pathway. Meanwhile, vitamin A deficiency might also mediate p38MAPK signalling to disturb the inflammatory cytokines in the MI and DI, but it did not appear to regulate inflammatory cytokines in the PI (see in Fig. 7 for details). In addition, based on the PWG, to protect fish against enteritis morbidity and ACP activity, the optimum dietary vitamin A levels required for optimal young grass carp growth, enteritis resistance and immune function were estimated to be 0·664, 0·707 and 0·722 mg/kg, respectively.

Fig. 7 General summary of the effects of vitamin A on immune function and its potential signalling pathways in the proximal intestine (PI) (a), mid intestine (MI) (b) and distal intestine (DI) (c) of young grass carp (Ctenopharyngodon idella). LA, lysozyme; ACP, acid phosphatase; C3, complement 3; C4, complement 4; LEAP, liver-expressed antimicrobial peptide; IKK, IκB kinase; I
κ
B, inhibitor κB; p38MAPK, p38 mitogen-activated protein kinase; TGF, transforming growth factor. ![]() , Not through;
, Not through; ![]() , inhibit;
, inhibit; ![]() , might through.
, might through.
Acknowledgements
The present study was jointly supported by the National Basic Research Program of China (973 Programme) (2014CB138600), the Specialized Research Fund for the Doctoral Program of Higher Education of China (20135103110001), Science and Technology Support Program of Sichuan Province of China (2014NZ0003), Major Scientific and Technological Achievement Transformation Project of Sichuan Province of China (2012NC0007; 2013NC0045), the Demonstration of Major Scientific and Technological Achievement Transformation Project of Sichuan Province of China (2015CC0011), Natural Science Foundation for Young Scientists of Sichuan Province (2014JQ0007) and Sichuan Province Research Foundation for Basic Research (2013JY0082). The authors thank the personnel of these teams for their kind assistance.
The author’s contributions are as follows: X.-Q. Z. and L. F. designed the study; L. Z. and L. F. conducted the study and analysed the data; Y. L., P. W., S.-Y. K., L. T., W.-N. T. and Y.-A. Z. participated in the interpretation of the results; L. Z. and W.-D. J. wrote the manuscript; X.-Q. Z. had primary responsibility for the final content of the manuscript. All the authors read and approved the final version of the manuscript.
The authors declare that there are no conflicts of interest.












