



Obesity represents a growing public health challenge(1), and it is particularly detrimental during early young adulthood as it increases the risk of developing chronic diseases later in life(Reference Ellison-Barnes, Johnson and Gudzune2). Obesity is typically defined through global metrics such as BMI, however, is a non-specific marker of body fat and provides no insight in body fat distribution(Reference Ahima and Lazar3). Specifically, Asians generally have a higher body fat than white people of the same age, sex and BMI category(4). Efforts to combat the obesity pandemic and its related metabolic diseases have mainly targeted traditional risk factors, e.g. excess energy intake and insufficient physical activity. However, traditional risk factors cannot fully explain the rapid rise of obesity during the past decades. Over the past 10 years, many causes of obesity and metabolic diseases have been identified, such as the timing of food intake(Reference Beccuti, Monagheddu and Evangelista5–Reference St-Onge, Ard and Baskin7).
Evidence for the importance of food timing for obesity comes from animal models(Reference Arble, Bass and Laposky8–Reference Fonken, Workman and Walton10) as well as observational(Reference Bernardes, Teixeira and Madalena11,Reference Dashti, Gomez-Abellan and Qian12) and experimental(Reference Allison, Hopkins and Ruggieri13,Reference Chellappa, Qian and Vujovic14) studies in humans, although not all studies are consistent(Reference Dote-Montero, Acosta and Sanchez-Delgado15,Reference Fiore, Scapaticci and Neri16) . Previous observational studies linking late eating with higher obesity risk revealed that food timing per se might alter body weight without a significant difference in energy intake and expenditure(Reference Ruiz-Lozano, Vidal and de Hollanda17,Reference Xiao, Garaulet and Scheer18) . However, most studies have utilised clock time to illustrate food timing(Reference St-Onge, Ard and Baskin7), which fails to accurately characterise food timing in the context of the circadian timing system. It is well established that there are individual differences in endogenous circadian timing relative to clock hour(Reference Horne and Ostberg19,Reference Roenneberg, Kuehnle and Juda20) , and that can vary by up to 10 h among college students(Reference McHill, Phillips and Czeisler21). This potential mismatch between clock and circadian timing may account for the contradictory associations between food timing and obesity in human studies.
Experimental evidence suggests the alignment between food timing and circadian rhythms, as with some forms of time-restricted eating or feeding, may prevent weight gain and promote metabolic health(Reference Manoogian, Chow and Taub22). However, food timing relative to endogenous circadian timing has not yet been thoroughly examined in human studies. McHill and colleagues(Reference McHill, Phillips and Czeisler21) examined the association between food timing relative to objective markers of circadian rhythms with adiposity in USA college students. The available evidence emphasises that it may be more physiologically relevant to consider the timing of food intake relative to endogenous circadian timing instead of only as clock time(Reference McHill, Phillips and Czeisler21).
Sex differences in the circadian system have been well characterised. For example, females have a shorter intrinsic circadian period and an earlier circadian phase than males(Reference Cain, Dennison and Zeitzer23,Reference Duffy, Cain and Chang24) . Given previous studies indicated that metabolic response to a meal depends on the timing of food intake relative to the individual’s circadian rhythm(Reference McHill, Phillips and Czeisler21,Reference Asher and Sassone-Corsi25,Reference Sofer, Stark and Madar26) , it is essential to explore the different metabolic responses to later circadian timing of food intake in males and females, which has not been investigated in previous research.
Hence, the objective of the present study was twofold: First, to examine the associations of food timing relative to clock time and endogenous circadian timing with general and visceral adiposity among Chinese college students and second to explore whether these associations differed by sex.
Methods
Participants
Participants were recruited from undergraduate students at local universities in Hefei, Anhui Province (China), using a multimethod approach spanning paper posters, flyers around campus and online advertisement (e.g. social media). The inclusion criteria included the following: (i) having the ability to download phone applications, (ii) not being engaged in a weight loss program, (iii) not taking any medication and (iv) not being engaged in night work within a 12-month period or traveling across time zones within a 3-month period before and during the procedure(Reference Phillips, Vidafar and Burns27). This study was conducted according to the guidelines laid down in the Declaration of Helsinki, and all procedures involving human subjects were approved by the Ethics Committee of the Anhui Medical University (No. 20180082). Written informed consent was obtained from all subjects.
Procedure
Pre-Laboratory procedures
Participants wore a wrist actigraphy and documented sleep and food schedules in real -time for 7 consecutive days.
Laboratory procedures
For calculation of circadian timing, participants were admitted to the laboratory for 1 night, arriving at approximately 14.00 h. After admission, participants completed the bioelectrical impedance analysis to assess adiposity. The lights were dimmed (< 10 lux) at 18.00 h. Salivary melatonin was sampled hourly from 19.00 h until 01.00 h. The following morning, participants were asked to void, and then their height and weight were measured in light clothing without shoes before eating or drinking anything. After completion of the laboratory procedures, participants were given breakfast and transported to school.
Measures
Demographics
At inclusion, participants filled in a set of questionnaires on socio-demographics and lifestyle, including age, sex, smoking status (current, former and never), alcohol status (current, former and never), consumption of sugar-sweetened beverages (never, 1–3 times/week, 4–6 times/week and ≥ 1 time/day) and screen time (< 2 h/d, 2–5 h/d and ≥ 5 h/d).
Dim-Light melatonin onset
Melatonin concentrations were determined from serial saliva samples (approximately 2 ml) collected hourly using Salivettes (Starstedt, Nümbrecht), beginning at 19.00 and ending at 01.00 h (seven measurements per subject) under dim-light conditions (< 10 lux), as verified using a portable illuminance meter (TES-1339R; Taishi Corp.). During the sample period, participants were seated in comfortable chairs, watched a dimmed TV (< 10 lux), talked to each other, had access to toilet facilities (also < 10 lux) and a research assistant was present at all times to ensure they did not fall asleep. Snacks and water were offered following each sampling, and participants rinsed their mouths with water 20 min before each sample collection. The saliva sample was centrifuged at 3300 g for 5 min. If there was less than 2 ml of saliva after centrifugation, the sample was centrifuged again at 3500 g for 5 min. Samples were frozen and stored at −80°C before being analysed. Melatonin levels were determined using a commercially available enzyme-linked immunosorbent assay kit from IBL (IBL International GmbH). Dim-light melatonin onset (DLMO) was calculated as the linear interpolated clock time at which evening salivary melatonin concentrations crossed and maintained above a 5 pg/ml threshold(Reference Phillips, Vidafar and Burns27).
Clock time of food intake
Each participant was asked by Tencent Document (https://docs.qq.com/mall/index) to complete a 7-day (5 weekdays and 2 weekend days) ecological momentary assessment of food timing, based on a signal-contingent approach at semifixed intervals (i.e., 07.00–10.00, 10.00–13.00, 13.00–16.00, 16.00–19.00 and 19.00–22.00)(Reference Barchitta, Maugeri and Favara28). To synchronise participants’ food timing information while minimising the burden on participants, students received notifications five times a day via Tencent Document, and each data entry took approximately 1–2 min. Participants reported the start timing of breakfast, lunch, dinner and first and last eating occasion in real-time, with an eating occasion (EO) defined as food or drink containing any amount of calories(Reference Dashti, Gomez-Abellan and Qian12). Eating midpoint was defined as the midpoint between the first and last EO(Reference Zeron-Rugerio, Hernaez and Porras-Loaiza29). Eating jetlag was calculated in hours as follows: Eating midpoint on weekends − Eating midpoint on weekdays, and all analyses were conducted using the absolute value of the estimated eating jetlag(Reference Zeron-Rugerio, Hernaez and Porras-Loaiza29). An experienced research staff instructed study participants on how to accurately complete the food records at the start of the study and reviewed these data with the participants every day to ensure the correctness of self-reported data.
Circadian timing of food intake
The duration between the average clock time of the last EO and DLMO (last EO-DLMO) was used to calculate the circadian timing of food intake. DLMO is a marker of the circadian phase and the beginning of an individual’s biological night(Reference Benloucif, Guico and Reid30). Therefore, the last EO-DLMO was conducted in an attempt to capture the EO that may have occurred during the biological night when energy expenditure was lower(Reference Zitting, Vujovic and Yuan31) and diet-induced thermogenesis tended to be lower as well(Reference Morris, Garcia and Myers32).
Sleep
For 7 consecutive days, participants were instructed to wear an actigraphy (ActiGraph GT3X+) on the non-dominant wrist for 24 h per day (except when bathing or swimming) and to complete an electronic sleep diary. Sleep onset, offset, duration and sleep fragmentation index were manually scored using the Sadeh algorithm in ActiLife software version 6·13·3 (Actigraph) from actigraphy and corroborated with sleep diary times(Reference Sadeh, Sharkey and Carskadon33). The midsleep point refers to the midpoint between actigraphy-derived sleep onset and offset.
BMI
BMI (kg/m2) was estimated using laboratory measurements: height and weight (without shoes and light clothing) were taken using standard protocols, without eating or drinking after the morning void.
Body composition
Body composition, including percent body fat (%), fat mass (FM, kg) and visceral fat area (VFA, cm2), were measured using an InBody S10 body composition analyser (Inbody Co., Seoul, Korea), a bioelectrical impedance analysis device that is portable, non-invasive and non-radiation. The VFA measured by bioelectrical impedance analysis using Inbody ® (Inbody Co., Seoul, Korea) demonstrated a good correlation with VFA observed by the gold standard method – computed tomography(Reference Lee, Park and Ahn34–Reference Nagai, Komiya and Mori36).
Analysis
Categorical variables were compared using the χ 2 test and continuous variables using paired or unpaired t tests. Possible linear and non-linear relationships between last EO (clock time/circadian timing) and adiposity were evaluated with multivariable-adjusted restricted cubic splines. Multiple linear regression models were used to test whether last EO (clock time/circadian timing) and sleep measures predicted adiposity. Further, sex-stratified analyses were performed to explore the sex differences of these associations. Scatter plots and Pearson correlations were used to examine the sex-specific association between later circadian timing of food intake with general and visceral adiposity. Age, sex, alcohol status, sugar-sweetened beverage consumption, screen time and sleep duration were included as covariates in multivariate linear regression models and restricted cubic splines models. Analyses were performed with R Software version 4·2·1 (R Foundation for Statistical Computing) and STATA 16 statistical software (Stata Corporation). Alpha < 0·05 (two-sided) was deemed statistically significant.
Results
General characteristics of the study participants
Of the 151 participants, 133 were included in the statistical analysis finally. Of the eighteen who were not included, five were not included because DLMO was not observed during the sampling period and thirteen did not have a complete 7-day food record (see Flow Chart, Fig. 1). The average age of the study population was 20·7 (s d, 0·8) years, 84 were females.

Fig. 1. Participant flow chart.
The main characteristics of study participants were presented by sex in Table 1. Compared with the male participants, female participants were more likely to have slightly earlier DLMO (female: 21:53, male: 22:06, P = 0·40) and last EO (clock time) of the day (19:40 v. 19:43, P = 0·83). When considering the circadian timing of last EO, females have the later circadian timing of last EO (i.e. last EO closer to DLMO) than males (–2·2 v. −2·4, P = 0·61). Female participants had higher percent body fat (28·0 v. 18·2, P < 0·001), FM (15·6 v. 12·8, P = 0·012) and VFA (65·9 v. 50·6, P = 0·004) than male participants, while male participants had higher BMI (20·9 v. 22·6, P = 0·003).
Table 1. Descriptive characteristics of participants by sex among 133 young adults (Numbers and percentages; mean values and standard deviations)

PBF, percent body fat; FM, fat mass; VFA, visceral fat area; DLMO, dim-light melatonin onset; EO, eating occasion; Last EO-DLMO, local time of the last EO minus DLMO.
Data are mean (sd) or n (%) for continuous and categorical variables, respectively. For last EO-DLMO, higher values denote last EO closer to, or after, DLMO.
Bold font is used to indicate statistically significant P value (P < 0·05).
Linear and nonlinear associations between last eating occasion (clock time/circadian timing) with general and visceral adiposity
Multivariable-adjusted restricted cubic splines demonstrated significant positive linear associations between last EO-DLMO with adiposity (BMI: P overall = 0·03, P nonlinear = 0·08; percent body fat: P overall = 0·02, P nonlinear = 0·35; FM: P overall = 0·03, P nonlinear = 0·21; VFA: P overall = 0·02, P nonlinear = 0·14) (Fig. 2 (e)–(h)). In addition, sex-stratified analyses revealed that these linear associations were significant in females but not males (online Supplementary Fig. 1 (a)–(h)).

Fig. 2. Restricted cubic splines representing the association between last EO (clock time/circadian timing) with general and visceral adiposity among 133 young adults. Heavy central line represents the estimated adjusted beta, with shaded ribbons denoting 95 % confidence interval. For last EO-DLMO, higher values denote last EO closer to, or after, DLMO. Splines were adjusted for age, sex (female, male), alcohol status (current, former and never), sugar-sweetened beverages consumption (never, 1–3 times/week, 4–6 times/week and ≥ 1 time/d), screen time (< 2 h/d, 2–5 h/d and ≥ 5 h/d) and sleep duration (h). DLMO, dim-light melatonin onset; EO, eating occasion; FM, fat mass; Last EO-DLMO, local time of the last EO minus DLMO; PBF, percent body fat; VFA, visceral fat area.
Association between last eating occasion (clock time/circadian timing) with general and visceral adiposity by sex
As shown in Table 2, the results demonstrated no association between the clock time of last EO with general and visceral adiposity, subanalyses after stratification for sex also revealed similar trends. However, when considering the circadian timing of last EO, significant associations were observed, such that individuals with their last EO closer to DLMO had higher BMI, percent body fat, FM and VFA (Table 2). Furthermore, sex-stratified analyses revealed that these associations were significant in females but not males (Fig. 3, Table 2). For females, each hour increase in last EO-DLMO was associated with higher BMI by 0·51 kg/m2 (95 % CI: 0·12, 0·91; P = 0·01), higher percent body fat by 1·05 % (95 % CI: 0·30, 1·80; P = 0·007), higher FM by 0·99 kg (95 % CI: 0·21, 1·77; P = 0·01) and higher VFA by 4·75 cm2 (95 % CI: 0·84, 8·65; P = 0·02), whereas no similar associations were observed among males.
Table 2. Association between last EO (clock time/circadian time) with general and visceral adiposity by sex among 133 young adults (Unstandardised beta and 95 % confidence intervals)
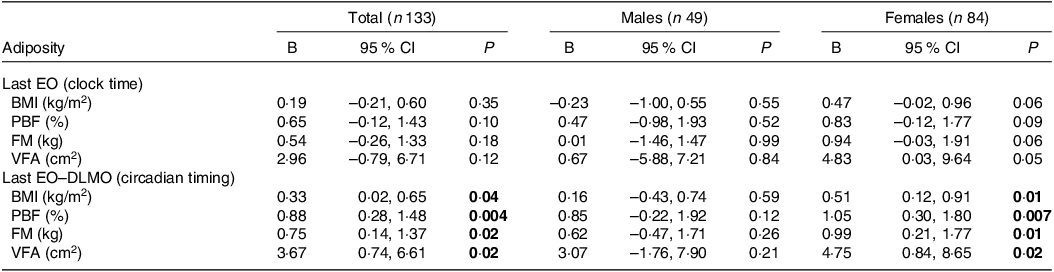
PBF, percent body fat; FM, fat mass; VFA, visceral fat area; DLMO, dim-light melatonin onset; EO, eating occasion; Last EO-DLMO, local time of the last EO minus DLMO; B = unstandardised beta.
Adjusted for age, sex (female, male), alcohol status (current, former and, never), sugar-sweetened beverages consumption (never, 1–3 times/week, 4–6 times/week and ≥ 1 time/day), screen time (< 2 h/d, 2–5 h/d and ≥ 5 h/d) and sleep duration (h). For last EO-DLMO, higher values denote last EO closer to, or after, DLMO.
Bold font is used to indicate statistically significant P value (P < 0·05).

Fig. 3. Scatter plots showing correlation between last EO-DLMO with BMI (a), PBF (b), FM (c) and VFA (d) by sex among 133 young adults. Pearson’s correlation coefficients and P value for each analysis are shown in the corresponding graph. For last EO-DLMO, higher values denote last EO closer to, or after, DLMO. DLMO, dim-light melatonin onset; EO, eating occasion; FM, fat mass; Last EODLMO, local time of the last EO minus DLMO; PBF, percent body fat; VFA, visceral fat area.
Association between sleep duration and sleep quality with general and visceral adiposity by sex
The results of the multiple linear regression models examining the associations of sleep duration and sleep quality with general and visceral adiposity are displayed in online Supplementary Tables 1 and 2. There were no significant associations between sleep duration and sleep quality with adiposity and results of stratified analyses by sex were consistent with overall results (both P > 0·05).
Discussion
The findings demonstrated that the timing of food intake relative to endogenous circadian timing rather than clock time was associated with general and visceral adiposity among young adults living in real-world settings. Strong linear associations between later circadian timing of food intake and adiposity were found among young females but not males. The findings highlight the importance of considering the timing of food intake relative to endogenous circadian timing instead of only as clock time. Nutrition strategies aimed to improve metabolic health should consider the circadian timing of food intake, particularly as our society moves toward personalised healthcare strategy.
Research on associations of food timing with adiposity is scant among young adults and has produced mixed results(Reference Fong, Caterson and Madigan37). One possible reason is that the clock time of food intake depends more on cultural and environmental factors such as work and school schedules, which may not accurately reflect the physiological impact of food timing on health. The present study indicated that later circadian timing of food intake (i.e. the timing of food intake relative to DLMO) was linked with higher general and visceral adiposity, while there were no differences when expressed in clock hours. These findings are consistent with a recent study conducted by McHill et al. (Reference McHill, Phillips and Czeisler21) and support the notion that examining the circadian timing of food intake rather than clock time could be more relevant when investigating associations with adiposity in young adults.
One potential mechanism for increased adiposity in response to later circadian timing of food intake is through effects on the thermic effect of food. Thermic effect of food is the energy expenditure in response to food intake(Reference Morris, Garcia and Myers32,Reference Bo, Fadda and Castiglione38,Reference Romon, Edme and Boulenguez39) , which is lower in the biological evening than morning due to the influence of the circadian system(Reference Morris, Garcia and Myers32). Given the association of adiposity with food timing relative to circadian timing rather than clock time, food intake closer to or after DLMO is a decrease in thermic effect of food, contributing to a positive energy balance and weight gain over time.
Another explanatory hypothesis is that the concurrence of food intake with elevated melatonin concentrations increases the probability of impaired glucose tolerance(Reference Garaulet, Lopez-Minguez and Dashti40,Reference Lopez-Minguez, Saxena and Bandin41) . It is known that postprandial glucose concentrations are higher in the biological evening than in the biological morning(Reference Bandin, Scheer and Luque42,Reference Morris, Yang and Garcia43) , and melatonin may further suppress the release of insulin in the biological evening(Reference Rubio-Sastre, Scheer and Gomez-Abellan44). Hence, eating earlier in the daytime to align with circadian rhythms may be preferred for improved blood glucose management, body weight control and related health outcomes, which will limit the concurrence of elevated glucose levels (due to postprandial state) and melatonin concentrations. Furthermore, later circadian timing of food intake may lead to a desynchronisation between the central and peripheral clocks and possibly result in metabolic disturbances(Reference Hernandez-Garcia, Navas-Carrillo and Orenes-Pinero45,Reference Scheer, Hilton and Mantzoros46) . In addition to the function of the circadian system, pathway analyses of the adipose tissue gene expression profiles revealed that late (circadian) eating modified lipid metabolic pathways in a direction consistent with increased adipogenesis and reduced lipolysis, contributing to increased lipid storage(Reference Vujovic, Piron and Qian47).
An interesting observation of the present study is the sex-specific association between later circadian timing of food intake with higher general and visceral adiposity in females but not males. One possible explanation for this observation is that the different metabolic response depends on the timing of food intake relative to an individual’s circadian system. Duffy and colleagues(Reference Duffy, Cain and Chang24) precisely examined the intrinsic circadian period in 157 healthy adults (33·1 % female) who underwent the gold-standard forced desynchrony protocol, and the results demonstrated that females have shorter intrinsic circadian periods than males and a greater proportion of females had intrinsic circadian periods shorter than 24 h. These sex-related circadian timing may explain the differential metabolic effects of food schedules among females v. males. However, whether and how sex modulates the adverse metabolic effects of later circadian timing of food intake is far less investigated. Further studies are needed to confirm these findings and investigate the underlying mechanism.
The absence of associations between sleep duration and sleep quality with adiposity in our results was unexpected, because insufficient sleep and poor sleep quality have been linked to higher weight(Reference Garaulet, Ortega and Ruiz48–Reference Zhao, Sun and He51). However, individuals with insufficient sleep may tend to increase food intake later in the day(Reference Markwald, Melanson and Smith52–Reference Spaeth, Dinges and Goel54) and later timing of food intake (closer to the biological night) is related to weight gain and obesity(Reference McHill, Phillips and Czeisler21).
Limitations of the study should be noted. There were no repeated assessments of circadian timing, thereby limiting interpretations of potential changes in circadian alignment with food timing, though DLMO is an extremely precise measure of circadian timing, and robust stability in DLMO time has been demonstrated in young adults living in real-world settings(Reference McHill, Sano and Hilditch55). A further key limitation of this study is the lack of data on dietary composition, which would have enhanced the analysis. Energy intake, macronutrient composition of the diet and overall diet quality may also interact synergistically with later circadian timing of food intake to influence adiposity(Reference Xiao, Garaulet and Scheer18), so that the observed effects may partly elated to the later circadian timing of food intake. However, the 7-day ecological momentary assessment of food records in this study provides more reliable and accurate data on food timing than questionnaire methods about usual mealtimes(Reference Maugeri and Barchitta56). In addition, we acknowledge that the population of university students may not be representative of the entire population in circadian timing of food intake. Older populations are more likely to have an earlier endogenous circadian phase and therefore may be more vulnerable to the effect of eating at a later clock time(Reference Knutson, Wu and Patel57) because it would be closer to their earlier DLMO. Other populations with known late eating at biological night are night shift workers, who have an increased risk of obesity(Reference Antunes, Levandovski and Dantas58) and metabolic syndrome(Reference De Bacquer, Van Risseghem and Clays59). Examination of the timing of food intake relative to DLMO in diverse populations is needed to extend our findings. Finally, the sample sizes differed between males and females in the current study, with males having a smaller sample size relative to females, which may affect the associations between later circadian timing of food intake and adiposity in males. Although the sex-specific association between later circadian timing of food intake with higher general and visceral adiposity is an important observation, comparable groups of males and females and longitudinal follow-up studies are needed to demonstrate the causality and potential mechanisms underlying this relationship.
Conclusions
In summary, the synthesis of real-world observations and precise laboratory data in the present study identified a new potential risk factor for weight gain and disease: food intake relative to endogenous circadian timing. The finding is particularly important for young adults and even teenagers, whose behaviour tends to be more evening-oriented. Future studies are needed to examine the interactions between circadian timing of food intake and energy intake, macronutrient profile and diet quality to influence weight trajectories, energy balance and metabolism over time. Furthermore, the sex-specific association observed in this study highlights an important, but under-explored, area of research – sex differences in later circadian timing of food intake effects, as well as the need to further investigate the underlying mechanisms.
Acknowledgements
The authors would like to thank all the volunteers for the participation and investigators for their contribution in the study.
This work was supported by the National Natural Science Foundation of China (Y.S., grant numbers 82173537) and Research Funds of Center for Big Data and Population Health of IHM (Y.S., grant numbers JKS2022008).
Y. H.: Conceptualisation, investigation, data curation, formal analysis, writing – review and editing. Y-x. X.: Methodology, data curation and writing – review and editing. Y-t. S.: Investigation and data curation. Y. Z.: Investigation and data curation. Y-h. W.: Formal analysis and supervision. F-b. T.: Conceptualisation and supervision. Y. S.: Conceptualisation, methodology, supervision and writing – review and editing. All authors approved the final submitted version.
The authors declared no conflict of interest.
Supplementary material
For supplementary material/s referred to in this article, please visit https://doi.org/10.1017/S0007114524001636







