


Obesity is a multifactorial disease with epidemic proportions. Its causes are mostly attributed to an excessive intake of energy together with low energy expenditure(Reference Jequier1). However, it is becoming increasingly clear that perinatal nutrition is also a key factor involved in the programming of later body weight. While moderate maternal energy restriction during lactation in rats has been associated in the offspring with lower body weight in adulthood(Reference Palou, Priego and Sanchez2, Reference Palou, Torrens and Priego3), severe maternal undernutrition during gestation, in animal models and also in humans, has been related to a greater propensity to develop obesity and related metabolic alterations, such as CVD or diabetes(Reference McMillen, MacLaughlin and Muhlhausler4, Reference Wells5). Therefore, it becomes interesting to investigate the mechanisms occurring in early postnatal life that could underlie these substantial differences, even before the appearance of differences in the phenotype, which may also be of great importance in the prevention of future health problems. In contrast to severe energy restriction, a moderate reduction of energy intake reflects better a more frequent condition in humans.
Concerning energy restriction during gestation and later obesity, we have described that moderate (20 %) maternal energy restriction during the first half of pregnancy in rats results in greater food intake in their offspring and this concludes in higher body weight in males but not in females(Reference Palou, Priego and Sanchez6). These animals, at the early age of 25 d, showed alterations in the structure and function of the hypothalamus, associated with an impairment of the central insulin and leptin action which could, at least, be in part responsible for the dysregulation of energy balance, leading to hyperphagia(Reference Garcia, Palou and Priego7, Reference Palou, Konieczna and Torrens8). However, which mechanisms account for the different outcomes between males and females of this prenatal condition are not known, beyond the presence of reduced sympathetic innervation in the inguinal adipose tissue, which occurred only in male animals, and could be responsible, at least in part, for the greater body weight and adiposity displayed in adulthood(Reference Garcia, Palou and Sanchez9).
Obesity is associated with an increased risk of morbidity and mortality as well as reduced lifespan(Reference Palou, Serra and Bonet10). The sirtuin family of NAD+-dependent protein deacetylases and ADP-ribosyltransferases are one of the most potential molecular targets for having an impact on ageing events(Reference Bishop and Guarente11, Reference Borradaile and Pickering12). Sirtuin 1 (SIRT1) has acquired great importance due to its participation in the control of energy metabolism-related genes and its ubiquitous distribution in the organism; higher expression of SIRT1 has been described to be advantageous as it activates catabolic metabolic pathways and increases longevity(Reference Pfluger, Herranz and Velasco-Miguel13, Reference Lagouge, Argmann and Gerhart-Hines14). As with SIRT1, sirtuin 2 (SIRT2) is an ubiquitous, nuclear and cytoplasmic protein deacetylase which was first involved in the prevention of the instability of the chromosomes during mitosis division(Reference Inoue, Hiratsuka and Osaki15). Later on, it has been shown to inhibit adipocyte differentiation, but its functions in the liver or muscle are not clearly elucidated(Reference North and Verdin16). All in all, the actions of SIRT1 in different tissues together with the actions of SIRT2 in adipose tissue might be able to stand up to obesity and the metabolic syndrome.
Considering the potential involvement of SIRT1 in the protection from the metabolic syndrome, the aim of the present study was to ascertain whether the detrimental effects of gestational maternal energy restriction on later metabolic health and body weight maintenance also involve changes in sirtuin expression in different tissues involved in energy homeostasis, and contribute to explaining the sex-different outcome on adult body weight.
Methods
Animals and experimental design
The study was performed in male and female rats from twelve different litters, following the protocol below, as described previously(Reference Palou, Priego and Sanchez6, Reference Garcia, Palou and Priego7, Reference Garcia, Palou and Sanchez9). All rats were housed under controlled temperature (22°C) and a 12 h light–dark cycle (lights on from 08.00 to 20.00 hours), and had unlimited access to tap water and a standard chow diet (13 kJ/g (3 kcal/g), with 2·9 % energy from fat; Panlab) unless mentioned otherwise. Briefly, virgin female Wistar rats weighing between 200 and 225 g were mated with male rats (Charles River Laboratories). Day of conception (day 0 of pregnancy) was determined by examination of vaginal smears for the presence of sperm, and then female rats were single caged. Pregnant rats were divided into two groups: one with free access to a standard chow diet and the other one underwent a 20 % restriction of energy intake from day 1 to day 12 of pregnancy. Energy restriction was performed by offering each dam a daily amount of food corresponding to 80 % of the energy that should be eaten according to body weight and to the day of pregnancy. This amount was calculated considering the energy daily consumed by their pregnant controls under ad libitum feeding conditions. After the energy restriction period, rats were allowed to eat ad libitum, and food intake was measured. On day 1 after delivery, excess pups in each litter were removed to keep ten pups per dam (five males and five females, when possible). Weaning was conducted at 21 d of life.
On day 25 of life, animals from control dams and from energy-restricted (ER) dams (n 6–9 animals per group) were killed by decapitation under fed conditions, during the first 2 h at the beginning of the light cycle. Blood samples were collected in heparinised containers, then centrifuged at 700 g for 10 min to obtain the plasma, and stored at − 20°C until analysis. Retroperitoneal and inguinal white adipose tissue (rWAT and iWAT, respectively), liver, skeletal muscle and hypothalamus were rapidly removed and frozen in liquid N2 and stored at − 80°C until ulterior studies. rWAT, iWAT and liver were weighed before freezing.
Although the retroperitoneal and the inguinal fat depots were sampled to be weighed, the retroperitoneal depot was selected as representative to be analysed for gene expression, based on the literature showing that this depot seems to be more sensitive to nutritional status, compared with other depots(Reference Palou, Sanchez and Priego17).
The animal protocol followed in the present study was reviewed and approved by the Bioethical Committee of our University and guidelines for the use and care of laboratory animals of the University were followed.
Measurement of circulating parameters
Blood glucose concentration was measured using an Accu-Chek Glucometer (Roche Diagnostics). Plasma insulin concentration was determined using a rat insulin ELISA kit (Mercodia AB) following standard procedures. Plasma leptin concentration was measured using a mouse leptin ELISA kit (R&D Systems).
RNA extraction
Total RNA was extracted from rWAT, iWAT, liver, muscle and hypothalamus by Tripure Reagent (Roche Diagnostic GmbH) according to the manufacturer's instructions. Isolated RNA was quantified using the NanoDrop ND-1000 spectrophotometer (NanoDrop Technologies, Inc.) and its integrity confirmed using 1 % agarose gel electrophoresis.
Real-time quantitative PCR analysis
Real-time PCR was used to measure mRNA expression levels of SIRT1 in rWAT, iWAT, liver, skeletal muscle and hypothalamus; SIRT2 in rWAT; 5′ adenosine monophosphate-activated protein kinase (AMPK) and adipose TAG lipase (ATGL) in rWAT and liver; uncoupling protein 2 (UCP2) in rWAT; and sterol response element binding protein 1c (SREBP1c), stearoyl-CoA desaturase-1 (SCD1) and the free fatty acid transporter CD36 in the liver. Briefly, 0·25 μg of total RNA (in a final volume of 5 μl) were denatured at 65°C for 10 min and then reverse transcribed to complementary DNA using MuLV reverse transcriptase (Applied Biosystems) at 20°C for 15 min, 42°C for 30 min, with a final step of 5 min at 95°C in an Applied Biosystems 2720 Thermal Cycler (Applied Biosystems). Each PCR was performed from diluted complementary DNA template, forward and reverse primers (1 μm each), and Power SYBER Green PCR Master Mix (Applied Biosystems). Primers were obtained from Sigma and sequences are described in Table 1. Real-time PCR was performed using the Applied Biosystems StepOnePlus™ Real-Time PCR Systems (Applied Biosystems) with the following profile: 10 min at 95°C, followed by a total of forty-two temperature cycles (15 s at 95°C and 1 min at 60°C). In order to verify the purity of the products, a melting curve was produced after each run according to the manufacturer's instructions. The threshold cycle (C t) was calculated by the instrument's software (StepOne Software version 2.1) and the relative expression of each mRNA was calculated as a percentage of male control rats, using the 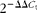 $2^{ - \Delta \Delta C _{t}}$ method(Reference Pfaffl18); β-actin and guanosine diphosphate dissociation inhibitor 1 (GDI1) were used as reference genes depending on the tissue, according to their better suitability.
$2^{ - \Delta \Delta C _{t}}$ method(Reference Pfaffl18); β-actin and guanosine diphosphate dissociation inhibitor 1 (GDI1) were used as reference genes depending on the tissue, according to their better suitability.
Table 1 Nucleotide sequences of primers used for PCR amplification

GDI1, guanosine diphosphate dissociation inhibitor 1; SIRT1, sirtuin 1; SIRT2, sirtuin 2; AMPK, 5′ AMP-activated protein kinase; ATGL, adipose TAG lipase; UCP2, uncoupling protein 2; SREBP1c, sterol response element binding protein 1c; SCD1, stearoyl-CoA desaturase 1; CD36, free fatty acid transporter CD36.
Statistical analysis
Data are expressed as means with their standard errors (n 6–9). Two-way ANOVA with the factors of sex (S) and energy restriction (R) was performed to assess statistical significances. Individual means were compared with a Student's t test. The analyses were performed with SPSS for Windows (SPSS). Threshold of significance was defined at P< 0·05. Data were log-transformed for statistical analysis if they were found not to be normally distributed, but were expressed as original values in the text and tables for ease of interpretation.
Results
Weight-related parameters and blood parameters
As previously described in the same cohort of animals(Reference Garcia, Palou and Priego7, Reference Garcia, Palou and Sanchez9), control and ER animals exhibited no significant differences in their body weight at the age of 25 d, either in the weight of rWAT, iWAT or liver (Table 2). Blood glucose and plasma insulin levels were not significantly different in ER animals compared with their controls. Circulating leptin concentration was lower in ER male rats, but not in females, with respect to their controls (P< 0·05, Student's t test; Table 2).
Table 2 Weight-related and blood parameters at the age of 25 d† (Mean values with their standard errors)

ER, energy-restricted dams during gestation; rWAT, retroperitoneal WAT; iWAT, inguinal WAT.
* Mean value was significantly different from its respective control group (P< 0·05; Student's t test). No significant differences were found by two-way ANOVA.
† Body weight, rWAT, iWAT and liver weight, and circulating glucose, insulin and leptin levels at 25 d of life (n 6–9) of male and female offspring from controls and ER animals, under ad libitum feeding conditions.
Sirtuin 1 mRNA expression in white adipose tissue, liver, hypothalamus and muscle
Fig. 1 shows mRNA expression levels of SIRT1 in the rWAT, liver, skeletal muscle and hypothalamus of controls and ER animals. Interestingly, a similar pattern of SIRT1 mRNA expression was found in the tissues studied. In rWAT, liver and skeletal muscle, ER male rats showed lower SIRT1 mRNA levels with respect to their controls (P< 0·05, Student's t test), whereas no significant differences were observed in females, and an interaction between the effect of energy restriction during gestation and sex was found in rWAT and skeletal muscle (P< 0·05, two-way ANOVA). An interaction between these factors was also found in the hypothalamus (P< 0·05, two-way ANOVA), but was attributed in this case to a tendency to higher mRNA levels in ER females compared with the controls (P= 0·067, Student's t test), with no differences in males.

Fig. 1 Sirtuin 1 mRNA expression levels in the retroperitoneal WAT (rWAT), liver, skeletal muscle and hypothalamus of 25-d-old male and female offspring of controls and energy-restricted dams during gestation (ER). mRNA levels were measured by real-time PCR and expressed as a percentage of the value of control male rats. Values are means, with their standard errors represented by vertical bars (n 6–9). R × S, interactive effect between energy restriction and sex (P< 0·05; two-way ANOVA). * Mean value was significantly different for the controls from that of ER animals (P< 0·05; Student's t test). □, Control males; ■, ER males; ![]() , control females;
, control females; ![]() , ER females.
, ER females.
To ascertain whether changes in SIRT1 expression found in rWAT could be representative for the WAT in general, SIRT1 expression was also measured in the subcutaneous inguinal depot. Interestingly, this depot showed a similar pattern to that of the retroperitoneal one, with a significant decrease in ER males with respect to their controls (100 (sem 20) and 33·0 (sem 3·7), controls and ER animals, respectively, P< 0·05, Student's t test), and no changes in females (61·2 (sem 8·3) and 61·8 (sem 8·1), controls and ER animals, respectively) (interaction between the effect of energy restriction during gestation and sex; P< 0·05, two-way ANOVA).
5′ Adenosine monophosphate-activated protein kinase, adipose TAG lipase and uncoupling protein 2 mRNA expression in retroperitoneal white adipose tissue
mRNA levels of the selected genes related to lipid metabolism in rWAT are shown in Fig. 2. ER male rats showed lower mRNA expression of the lipolysis-related gene ATGL compared with the controls (P< 0·05, Student's t test), while a tendency to higher mRNA levels was found in ER females (P= 0·071, Student's t test) with respect to their controls (interaction between the effect of energy restriction during gestation and sex; P< 0·05, two-way ANOVA). A similar tendency, but not significant, was also found for AMPK. Higher mRNA expression levels of AMPK were observed in females compared with males (P< 0·05, two-way ANOVA).

Fig. 2 mRNA expression levels of the selected genes related to lipid metabolism in the retroperitoneal WAT of 25-d-old male and female offspring of controls and energy-restricted dams during gestation (ER). mRNA levels were measured by real-time PCR and expressed as a percentage of the value of control male rats. Values are means, with their standard errors represented by vertical bars (n 6–9). The genes determined were 5′ adenosine monophosphate-activated protein kinase (AMPK), adipose TAG lipase (ATGL) and the uncoupling protein 2 (UCP2). S, effect of sex; R × S, interactive effect between energy restriction and sex (P< 0·05; two-way ANOVA). * Mean value was significantly different for the controls from that of ER animals (P< 0·05; Student's t test). □, Control males; ■, ER males; ![]() , control females;
, control females; ![]() , ER females.
, ER females.
Of interest, UCP2 mRNA expression was also decreased in ER males with respect to their controls (P< 0·05, Student's t test), whereas females did not show differences between the groups (interaction between the effect of energy restriction during gestation and sex; P< 0·05, two-way ANOVA).
5′ Adenosine monophosphate-activated protein kinase, adipose TAG lipase, sterol response element binding protein 1c, stearoyl-CoA desaturase-1 and CD36 mRNA expression in liver
Fig. 3 shows the mRNA expression levels of the selected genes related to lipid metabolism in the liver. Both male and female ER rats showed lower mRNA levels of AMPK and ATGL in comparison with the controls (P< 0·05, two-way ANOVA). It was observed that ER male animals exhibited lower mRNA levels of SREBP1c and SCD1 than the controls, unlike female ER rats which showed higher mRNA levels compared with the controls (interaction between the effect of energy restriction during gestation and sex; P< 0·05, two-way ANOVA). A similar pattern to that of SREBP1c and SCD1 was also observed concerning CD36 mRNA expression levels, although differences did not reach statistical significance.

Fig. 3 mRNA expression levels of the selected genes related to lipid metabolism in the liver of 25-d-old male and female offspring of controls and energy-restricted dams during gestation (ER). mRNA levels were measured by real-time PCR and expressed as a percentage of the value of control male rats. Values are means, with their standard errors represented by vertical bars (n 6–9). The genes determined were 5′ adenosine monophosphate-activated protein kinase (AMPK), adipose TAG lipase (ATGL), sterol regulatory element binding protein 1c (SREBP1c), stearoyl-CoA desaturase-1 (SCD1) and the free fatty acid transporter CD36 (CD36). R, effect of energy restriction; R × S, interactive effect between energy restriction and sex (P< 0·05; two-way ANOVA). * Mean value was significantly different for the controls from that of ER animals (P< 0·05; Student's t test). □, Control males; ■, ER males; ![]() , control females;
, control females; ![]() , ER females.
, ER females.
Sirtuin 2 mRNA expression in retroperitoneal white adipose tissue
Fig. 4 shows mRNA expression levels of SIRT2 in rWAT. ER animals showed a tendency to lower SIRT2 mRNA levels compared with their controls (P= 0·072, two-way ANOVA), but differences were more marked and statistically significant only in females (P< 0·05, Student's t test).

Fig. 4 Sirtuin 2 mRNA expression levels in the retroperitoneal white adipose tissue of 25-d-old male and female offspring of controls and energy-restricted dams during gestation (ER). mRNA levels were measured by real-time PCR and expressed as a percentage of the value of control male rats. Values are means, with their standard errors represented by vertical bars (n 6–9). * Mean value was significantly different for the controls from that of ER animals (P< 0·05; Student's t test). A tendency to lower sirtuin 2 mRNA levels in ER rats was found with two-way ANOVA (P =0·072). □, Control males; ■, ER males; ![]() , control females;
, control females; ![]() , ER females.
, ER females.
Discussion
Here we show that a moderate maternal energy restriction of 20 % during the first half of pregnancy programmes a characteristic profile of SIRT1 mRNA expression in different key tissues involved in the control of energy metabolism in male and female offspring, which may be related to the different predisposition of these animals to overweight development in later life(Reference Palou, Priego and Sanchez6, Reference Palou, Konieczna and Torrens8).
The thrifty phenotype hypothesis proposes that undernutrition during early life may induce lasting alterations in tissue and organ function to increase efficiency using available nutrients when they are in short supply, preserving glucose for the development of main organs in the fetus, particularly the brain(Reference Hales and Barker19). These prenatal alterations to a poor nourishment may be detrimental in a positive nutritional environment, because the balance setpoint for nutrient availability has been changed, leading to storage at the expenses of energy expenditure, therefore increasing the risk of obesity development and associated diseases in adulthood(Reference McMillen, MacLaughlin and Muhlhausler4, Reference Wells5, Reference Hales and Barker19).
In accordance with this hypothesis, we have previously described that moderate undernutrition during fetal life (achieved by 20 % maternal energy restriction during the first part of gestation) was associated with hyperphagia in postnatal life, which resulted in higher body weight in adulthood in males but not in females(Reference Palou, Priego and Sanchez6, Reference Palou, Konieczna and Torrens8). The hyperphagia could, in part, be explained by alterations in the hypothalamic structures related to the control of food intake(Reference Garcia, Palou and Priego7), and by a lower capacity to respond to insulin and leptin action at the central level, which was already present at the premature age of 25 d of life(Reference Palou, Konieczna and Torrens8). However, the reasons for the sex-dependent consequences in later body weight have not yet been clearly elucidated.
The sirtuin family of NAD-dependent protein deacetylases and ADP-ribosyltransferases regulates the activities of a variety of transcription factors, co-regulators and enzymes improving obesity resistance and expanding lifespan(Reference Holness, Caton and Sugden20). Here we found that ER male animals, but not females, displayed lower mRNA expression levels of SIRT1 in comparison with the controls in WAT, liver and skeletal muscle, before any difference in body weight, but in accordance with their later overweight when adults(Reference Palou, Priego and Sanchez6). Hence, it could be hypothesised that programmed changes in SIRT1 expression may, to a certain extent, account for the sex-dependent outcomes on the body weight of adult ER animals. Holness et al. (Reference Holness, Caton and Sugden20) have also recently propounded a link between the thrifty phenotype, programmed changes in SIRT1 expression and energy supply. In agreement, other studies have also described changes in SIRT1 expression depending on the perinatal nutritional conditions; while maternal protein restriction during the lactation period resulted in increased SIRT1 mRNA expression levels in the kidney of the offspring, together with improved insulin sensitivity(Reference Martin-Gronert, Tarry-Adkins and Cripps21), the offspring of protein-restricted mice during gestation showed lower SIRT1 protein expression in the skeletal muscle(Reference Chen, Martin-Gronert and Tarry-Adkins22). Therefore, the sex-related differences found here in ER animals concerning SIRT1 mRNA levels and the similar expression profile found in different tissues, namely WAT (including one internal and one subcutaneous depot), liver and skeletal muscle, increase the power of these findings suggesting a main role of SIRT1 expression in peripheral tissues in the programming of adult body weight. It could be speculated that the maintenance of SIRT1 expression in peripheral tissues of ER females could explain or contribute, at least in part, to the capacity of these animals to maintain their body weight in adulthood despite their higher energy intake.
Unlike the potential anti-obesogenic actions of SIRT1 in peripheral tissues, hypothalamic SIRT1 expression increases by fasting in rats(Reference Cakir, Perello and Lansari23) and mice(Reference Ramadori, Lee and Bookout24), and fasting-induced hyperphagia has been shown to require hypothalamic SIRT1 activity(Reference Cakir, Perello and Lansari23). SIRT1 at the central level has been proposed to sense the nutritional status of the body and regulate the hypothalamic central melanocortin signalling and S6K pathway to govern food intake and body weight(Reference Cakir, Perello and Lansari23). Therefore, it is interesting to highlight that SIRT1 mRNA levels in the hypothalamus were not different between the control and ER animals; in fact, an opposite tendency (P= 0·067, Student's t test) to higher mRNA levels was even found in ER females with respect to their controls, which is in accordance with their higher food intake. However, we have to take into account that the results presented here are based on gene expression and further analyses are needed to determine whether changes in SIRT1 mRNA levels are reflecting changes in its protein amount or activity.
SIRT1 is frequently defined as a master metabolic regulator since it is able to influence several transcription factors involved in energy homeostasis; in fact, it has been described to regulate glucose and lipid metabolism in the liver, WAT, pancreas and muscle, increasing lipid catabolism and energy expenditure(Reference Schug and Li25). Thus, to ascertain whether changes in SIRT1 mRNA expression could be accompanied with changes in lipid metabolism and energy expenditure, and in order to explain the sex differences in adult body weight, the expression of the selected genes in rWAT and liver was also studied. Lower mRNA levels of AMPK in the liver of both male and female ER rats and a tendency to lower levels in the rWAT of ER males were found compared with the controls. AMPK stimulates catabolic pathways in order to generate ATP and inhibits non-essential anabolic processes that consume ATP when energy status is decreased(Reference Ruderman, Xu and Nelson26), and it has recently been described that SIRT1 and AMPK regulate each other and share several common target molecules(Reference Ruderman, Xu and Nelson26). In fact, AMPK activity is reduced in most genetic models of rodent obesity in peripheral tissues such as heart, skeletal muscle and liver(Reference Steinberg and Kemp27). Interestingly, the changes observed here in AMPK mRNA levels at this early stage of life are early events before changes in body weight. Leptin has been described to activate AMPK expression(Reference Minokoshi, Kim and Peroni28, Reference Pico, Jilkova and Kus29), therefore changes in AMPK mRNA levels could be partially explained by changes in circulating leptin levels, which were lower in ER male controls compared with their controls. However, the decrease in AMPK mRNA levels also occurring in the liver of ER female animals does not appear to be explained by changes in circulating leptin levels, which remained unchanged in these animals. Nevertheless, considering that leptin present in breast milk can be absorbed by the immature stomach of suckling rats and affect later development(Reference Sanchez, Oliver and Miralles30–Reference Palou, Pico and McKay32), the mentioned changes in AMPK expression levels could be reflecting changes in the amount of leptin ingested during lactation, more than a direct effect of their circulating levels after weaning. In fact, as previously described in the same model of animals(Reference Palou, Konieczna and Torrens8), the offspring of 20 % ER animals during gestation did not show the expected peak in circulating leptin levels during the neonatal period, a process termed as ‘neonatal leptin surge’(Reference Pico, Jilkova and Kus29), which occurred in control animals at the age of 9 d.
AMPK has been described to activate the lipolytic pathway by regulating hormone-sensitive lipase (HSL) activity(Reference Steinberg and Kemp27). However, ATGL has recently been identified as an important TAG lipase that regulates lipid hydrolysis upstream of HSL (Reference Steinberg and Kemp27). Here, both male and female ER animals displayed a lower mRNA expression of ATGL in the liver, in accordance with the lower AMPK mRNA levels; yet only ER males showed lower ATGL mRNA levels in rWAT, a similar pattern to that found for AMPK in this tissue. It could be suggested that reduced lipid mobilisation in adipose tissue is early programmed only in males, whereas hepatic lipid metabolism, particularly catabolic pathways, is altered in both male and female ER animals. In fact, dysregulation of adipose tissue and hepatic lipid metabolism has been described in the same cohort of animals in adulthood at the age of 6 months(Reference Palou, Konieczna and Torrens8). On the other hand, ER males, but not females, also showed lower UCP2 mRNA levels in rWAT with respect to their controls. Although the primary role of UCP2 is not clearly established, considerable evidence suggests that UCP2 plays a physiological role in regulating fatty acid oxidation, mitochondrial biogenesis, substrate utilisation and reactive oxygen species elimination(Reference Pecqueur, Bui and Gelly33). Hence, the lower expression levels found in ER male animals could be related to lower lipid mobilisation and fatty acid oxidation. Moreover, although a direct association between UCP2 and obesity or energy expenditure is not well established(Reference Schrauwen and Hesselink34), higher mRNA levels of UCP2 have been described in the WAT of obesity-resistant mice compared with obesity-prone mice(Reference Fleury, Neverova and Collins35), and lower expression levels have been reported in the WAT of obese human subjects(Reference Oberkofler, Liu and Esterbauer36); therefore, the decrease in the expression of this gene occurring only in male ER animals and not in females, as occurring with AMPK and ATGL, may also be related to the different outcomes of these prenatal conditions on later body weight between males and females.
In addition to AMPK and ATGL, the expression of the other genes involved in lipid metabolism, in particular lipogenic genes such as SREBP1c, SCD1 and CD36, was also studied in the liver. Of interest, energy restriction during gestation resulted in a different pattern of expression between males and females concerning SREBP1c and SCD1. Their expression levels decreased in ER males but increased in ER females. A similar pattern was also found for CD36, although differences did not reach statistical significance. These changes could be reflecting impaired insulin signalling, as the expression of SREBP1c is up-regulated by insulin(Reference Cagen, Deng and Wilcox37), and SREBP1c regulates fatty acid biosynthetic genes such as SCD1 (Reference Horton, Goldstein and Brown38). In this sense, we have previously described that this condition during gestation programmes the offspring, both males and females, for a lower capacity to respond to insulin action, which is already present at early ages(Reference Palou, Konieczna and Torrens8). Therefore, although insulin resistance in the liver seems to be early programmed in both male and female animals, other factors occurring in females may protect and counterbalance this malprogramming effect due to energy restriction on energy homeostasis. The results of the present study support a potential role of SIRT1 in these differences.
To explore other members of the sirtuin family as potential markers of later obesity, mRNA expression levels of SIRT2 in rWAT were analysed. Unlike SIRT1, the role of SIRT2 has not been explored as much and its action in some tissues needs to be clarified. Nevertheless, in WAT, SIRT2 is known to inhibit adipocyte differentiation by regulating Forkhead box protein O1 (FOXO1) acetylation(Reference Jing, Gesta and Kahn39). Here, we found a tendency to lower SIRT2 mRNA levels in rWAT in ER animals compared with the controls (P= 0·072), although it was more pronounced and only significant by Student's t test in ER females. We previously described that the accumulation of an excessive amount of fat in ER males was related to hyperplasia in the iWAT and to hypertrophy in the rWAT(Reference Garcia, Palou and Sanchez9). Thus, the tendency to lower SIRT2 mRNA levels in ER males, together with the lower mRNA levels of SIRT1, ATGL and UCP2 in the rWAT, agrees with later hypertrophy development in this depot when rats were exposed to a high-fat diet in adulthood(Reference Garcia, Palou and Sanchez9). However, unlike males, changes occurring in SIRT2 mRNA levels in the rWAT of females do not directly agree with their capacity to maintain body weight in adult life, supporting a major role for SIRT1 in the regulation of energy homeostasis.
In conclusion, the present study shows that a moderate maternal energy restriction of 20 % during the first half of gestation programmes a particular, sex-dependent expression profile of SIRT1 in different peripheral tissues, which may be associated with obesity susceptibility and related metabolic disorders in adulthood; this suggests that SIRT1 mRNA expression levels in particular tissues may be considered as potential early biomarkers of obesity susceptibility.
Acknowledgements
This study was supported by the Spanish Government (grant AGL2009-11277), the European Union's Seventh Framework Programme FP7 2007-2013 under grant agreement no. 244995 (BIOCLAIMS Project) and the Instituto de Salud Carlos III, Centro de Investigación Biomédica en Red Fisiopatología de la Obesidad y Nutrición, CIBERobn. The Laboratory is a member of the European Research Network of Excellence NuGO (The European Nutrigenomics Organization, EU Contract no. FP6-506360). C. P., A. P. and M. P. designed the research. M. P. carried out the animal treatment, follow-up and sample collection and performed the analyses. T. P. and J. S. participated in the sample collection and in the discussion of the results. M. P., C. P. and A. P. carried out the discussion of the results and wrote the paper. All authors read and approved the final manuscript. The authors declare that they have no conflict of interest.






