


Primary liver cancer (PLC) is the sixth most commonly diagnosed cancer and the third leading cause of cancer death worldwide(Reference Sung, Ferlay and Siegel1). China alone accounts for over half of the new cases and deaths, with 410 038 new cases and 391 152 deaths in 2020(Reference Sung, Ferlay and Siegel1). Hepatocellular carcinoma (HCC) is the most predominant type of PLC. The prognosis of HCC is generally poor, with 5-year net survival ranging from 5 to 30 %(Reference Shao, Hu and Wang2). In addition to the established prognostic factors of underlying liver function, tumour stage, performance status and treatments(Reference Gilles, Garbutt and Landrum3), and other factors may also affect HCC prognosis.
Impaired folate-mediated one-carbon metabolism (FOCM) could contribute to cancer development and progression due to the crucial role of folate-mediated one-carbon metabolism in DNA synthesis, repair and methylation(Reference Newman and Maddocks4). Our previous study has associated lower serum folate concentrations at diagnosis with worse HCC survival(Reference Fang, Liu and Liao5). Methylenetetrahydrofolate reductase (MTHFR), 5-methyltetrahydrofolate-homocysteine methyltransferase (MTR) and 5-methyltetrahydrofolate-homocysteine methyltransferase reductase (MTRR) are three key enzymes involved in the folate-mediated one-carbon metabolism (Fig. 1). MTHFR irreversibly catalyses the conversion of 5,10-methylenetetrahydrofolate (5,10-methylene THF) to 5-methyltetrahydrofolate (5-methyl THF), the dominant circulating form of folate. Two common polymorphisms in the MTHFR gene, rs1801133 and rs1801131, reduce enzyme activity and lead to lower levels of 5-methyl-THF(Reference Weisberg, Tran and Christensen6,Reference Rozen7) . Previous genome-wide association studies have identified rs1801133 as the gene locus associated with serum folate levels(Reference Grarup, Sulem and Sandholt8). MTR and MTRR are responsible for the biosynthesis of methionine and the regeneration of THF for nucleotide biosynthesis(Reference Leclerc, Wilson and Dumas9). Gene variants of MTR rs1805087 and MTRR rs1801394 may cause decreased activity of the MTR enzyme(Reference Jokić, Brčić-Kostić and Stefulj10,Reference Wang, Li and Wang11) . Previous animal experiments have shown that MTR maintains the tumour tetrahydrofolate pool to drive nucleotide synthesis and cell proliferation in cancer cells(Reference Ghergurovich, Xu and Wang12,Reference Sullivan, Darnell and Reilly13) . Therefore, genetic polymorphisms in the genes encoding folate metabolism-related enzymes may influence enzyme activity and interact with folate status, ultimately affecting cancer survival.

Fig. 1. Overview of folate-mediated one-carbon metabolism (OCM) and related enzymes. 5,10-methylene THF, 5,10methylenetetrahydrofolate; 5-methyl-THF, 5-methyltetrahydrofolate; DHF, dihydrofolate; dTMP, deoxythymidine; MTHFR, methylenetetrahydrofolate reductase; MTR, 5-methyltetrahydrofolate-homocysteine methyltransferase reductase; MTRR, 5-methyltetrahydrofolate-homocysteine methyltransferase reductase; SAM, S-adenosylmethionine; THF, tetrahydrofolate.
To our knowledge, only six small studies including 71–244 cases have previously investigated the association between several polymorphisms in folate-metabolising genes (MTHFR rs1801133, MTHFR rs1801131, MTR rs1805087 or MTRR rs1801394) and the prognosis of HCC and produced inconsistent results(Reference Wang, Lu and Ling14–Reference Peres, Galbiatti-Dias and Castanhole-Nunes19). Of note, most studies were limited to specific patients, such as patients with chronic hepatitis B-related liver cancer and liver transplant recipients, which may underrepresent the overall patients with HCC. Additionally, most studies solely focused on the individual role of genetic variants in the candidate genes and neglected the potential interaction between these polymorphisms and folate status on the association with HCC survival. To date, only a Taiwanese cohort study suggested that HCC patients carrying the MTHFR rs1801133 CC genotype and with high erythrocytes folate levels had worse survival compared with the same genotype(Reference Kuo, Huang and Kuo15). However, apart from the MTHFR rs1801133 polymorphism, the role of other genetic variants in folate-metabolising genes and their interactions with folate status in HCC prognosis remain largely unexplored.
Therefore, our aims were (1) to investigate the association between folate-metabolising gene (MTHFR, MTR or MTRR) polymorphisms and the prognosis of HCC and (2) to explore whether these genetic variants modify the association of serum folate concentrations with HCC survival in the Guangdong Liver Cancer Cohort study (GLCC).
Materials and methods
Study population
Our study participants were from the GLCC, an ongoing prospective cohort study initiated in 2013 to investigate factors affecting PLC progression and survival. The study design has been described in detail elsewhere(Reference Fang, Chen and Wang20). In brief, we recruited untreated patients aged 18–80 years who were newly diagnosed with PLC within 1 month at the Sun Yat-sen University Cancer Center, China. A total of 1359 PLC patients were enrolled in the GLCC between September 2013 and April 2017. After excluding fifty-seven cases with a confirmed diagnosis of PLC other than HCC (e.g. intrahepatic cholangiocarcinoma and HCC-ICC) and 332 cases who had no available blood samples for SNP genotyping, a total of 970 eligible patients were included in this study. Among them, 864 patients also had serum folate measurements. The selection of the study participants is presented in online Supplementary Fig. 1.
Ethical approval for this study was obtained from the Ethics Committee of the School of Public Health at Sun Yat-sen University, and the study was conducted according to the Helsinki Declaration of Ethics. All participants provided informed consent at the time of recruitment.
Laboratory assays
Fasting venous blood was collected before anticancer treatment and stored at –80°C after centrifugation. Serum folate concentrations were quantified in batches using a chemiluminescent microparticle immunoassay (ARCHITECT Folate assay, Abbott Diagnostics) at the KingMed Diagnostics Laboratory (Guangzhou, China)(Reference Fang, Liu and Liao5).
Routine laboratory parameters, including α-fetoprotein, alanine aminotransferase, aspartate aminotransferase, γ-glutamyltransferase, alkaline phosphatase, albumin and total bilirubin and C-reactive protein, were analysed according to a standardised protocol at the Clinical Laboratory of Sun Yat-sen University Cancer Center. We constructed a liver damage score (ranging from 0 to 6) by summing the number of abnormal laboratory-defined values for six hepatic function tests (ALT > 50 μ/l, aspartate aminotransferase > 40 μ/l, γ-glutamyltransferase > 60 μ/l, alkaline phosphatase > 150 μ/l, albumin < 40 g/l and total bilirubin > 20·5 μmol/l) to assess preexisting chronic liver diseases(Reference Fedirko, Duarte-Salles and Bamia21). A score of 0 indicated no liver injury, 1–2 represented possible minor liver injury and ≥ 3 suggested possible liver injury.
Clinical and lifestyle data collection
Demographic characteristics (e.g. age and sex) and diagnostic and treatment information were obtained from the Sun Yat-sen University Cancer Center electronic management system. The Barcelona Clinic Liver Cancer stage was chosen to assess tumour severity, which comprehensively considers tumour number and size, Child-Pugh score (an indicator of the severity of liver dysfunction and hepatic functional reserve)(Reference Pugh, Murray-Lyon and Dawson22) and performance status of the patients(Reference Llovet, Brú and Bruix23). We recorded the primary cancer treatment that patients received after diagnosis.
Information on lifestyles was obtained through baseline interviews using a structured questionnaire. Participants were classified into three groups according to smoking status: never smokers, former smokers and current smokers. Current smokers were defined as those who had smoked at least one cigarette per day for at least 6 months and former smokers were defined as those who had quit smoking for at least 1 year. Weight and height were measured following a standard procedure. BMI was calculated by dividing weight in kilograms (kg) by height in metres squared (m2).
DNA extraction, genotyping and single nucleotide polymorphisms selection
Genomic DNA was extracted from blood clots using TIANGEN blood clot genomic DNA extraction kits (Tiangen Biochemical Technology (Beijing) Co., Ltd., DP335-02). The purity and concentration of the extracted DNA samples were determined using a Nanodrop one ultra-micro spectrophotometer (Thermo Fisher Scientific), and the ratio of absorbance at 260 and 280 nm (A260/A280) ranged between 1·8 and 2·0 was acceptable. We selected six candidate single nucleotide polymorphisms (SNP) in folate-metabolising genes, including MTHFR rs1801133, MTHFR rs1801131, MTHFR rs2274976, MTR rs1805087, MTRR rs1801394 and MTRR rs10380. We chose SNP with an expected minor allele frequency > 5 % in the Chinese population, which have been previously shown to be associated with HCC or have potential functional significance. These loci were genotyped using the Kompetitive allele specific polymerase chain reaction assay(Reference Kalendar, Shustov and Akhmetollayev24). We designed three primers for each SNP, including two forward-specific primers and one reverse universal primer. Two forward primers corresponded to two kinds of fluorescence signals. After PCR amplification, we measured the fluorescent values of the two signals to determine the genotype of the samples. Strict quality controls were performed by setting up negative and positive controls during the genotyping. The genotype missing rate of MTHFR rs1801133, MTHFR rs1801131, MTHFR rs2274976, MTR rs1805087, MTRR rs1801394 and MTRR rs10380 was 3·4 %, 1·9 %, 1·4 %, 5·8 %, 3·8 % and 3·5 %, respectively.
Survival outcomes
Participants were followed from the date of blood donation until the date of death or the last date known alive whichever occurred first. The last outcome ascertainment was carried out on 22 February 2019. Survival outcomes assessed included overall survival (OS) and liver cancer-specific survival (LCSS). The outcome event was all-cause death for OS and death from HCC for LCSS. The date and cause of death were ascertained by referring to the death registration and reporting system of the Guangdong Provincial Center for Disease Control and Prevention, combined with the inpatient and outpatient medical system of the Sun Yat-sen University Cancer Center. In addition, we conducted telephone-based interviews with the patients or their next-of-kin every 6–12 months to confirm their survival status.
Statistical analysis
We compared clinical and nonclinical characteristics between eligible and ineligible participants, as well as among eligible participants with different genotypes of the selected SNP. Differences among the groups were analysed using one-way ANOVA, Wilcoxon rank sum test or Kruskal–Wallis rank sum test for continuous variables and Pearson’s χ 2 test for categorical variables.
Three genetic models (co-dominant (wild-type v. heterozygote v. homozygote), dominant (wild-type v. heterozygote + homozygote) and additive (per variant allele)) were applied to assess the association between genetic polymorphisms in folate-metabolising genes and LCSS and OS. Cox proportional hazards models were used to calculate hazard ratios (HR) and 95 % CI. Model 1 was a crude model. Model 2 was adjusted for non-clinical factors, including age at diagnosis (continuous), sex (women, men), BMI (< 18·5, 18·5–24·0, 24·0∼28·0 and ≥ 28·0 kg/m2) and smoking status (never, former and current). Model 3 was further adjusted for clinical factors, including α-fetoprotein levels (≤ 400 ng/ml, > 400 ng/ml), C-reactive protein levels (≤ 3·0 mg/l, > 3·0 mg/l), liver damage score (0, 1∼2 and ≥ 3), Barcelona Clinic Liver Cancer stage (0, A, B and ≥C) and cancer treatment (hepatectomy/liver transplantation, local ablation, hepatic arterial intervention and other treatments). Since only a few covariates were missing with a small proportion, BMI, C-reactive protein levels and α-fetoprotein levels with missing data (n 3) were automatically excluded from the multivariable models. The proportional hazards assumption was verified using the global Schoenfeld residual test. Sensitivity analyses were conducted by excluding female participants to minimise the impact of gender differences.
Joint associations of folate metabolism-related gene polymorphisms with LCSS and OS were analysed based on the number of protective alleles from the six polymorphisms, where the C allele of the rs1801133 polymorphism, the A allele of the rs1801131 polymorphism, the G allele of the rs2274976 polymorphism, the A allele of the rs1805087 polymorphism, the G allele of the rs1801394 polymorphism and the C allele of the rs10380 polymorphism were considered protective.
We evaluated whether the association between sex-specific quartiles of serum folate concentrations and survival outcomes in HCC patients was modified by the genotype of MTHFR, MTR and MTRR, as well as the number of protective alleles, on both multiplicative and additive scales. The multiplicative interaction was assessed by comparing the −2 log-likelihood of the fully-adjusted models with and without the cross-product interaction term of the sex-specific quartiles of serum folate levels and the SNP genotype (i.e. all tests for interaction are 1 degree of freedom). Stratified analyses by the genotypes were subsequently performed. Linear trends were tested by entering the median value of sex-specific quartiles of serum folate levels as a continuous variable in the regression models. To assess the additive interaction, we treated serum folate concentrations (low-≤median and high->median) and the genotypes (wild-type, mutant) as dichotomised variables, The relative excess risk due to the interaction (RERI) and the attributable proportion due to the interaction (AP) were used to estimate the deviation from the additivity of the effect(Reference Knol and VanderWeele25), with the delta method to obtain CI for the indices(Reference Hosmer and Lemeshow26).
Statistical analyses were performed using SPSS version 26·0 (IBM Corp.) and R software version 4·2·3 (R Foundation for Statistical Computing, Vienna, Austria). All P values were two-sided, and P < 0·05 was considered statistically significant.
Results
Baseline characteristics
No significant differences in clinical and nonclinical characteristics were observed between eligible and ineligible participants except for cancer treatment received (online Supplementary Table 1). A lower proportion of eligible participants underwent hepatic resection or liver transplantation compared with ineligible participants. Of the 970 patients included, 857 (88·4 %) were men. The mean age at diagnosis was 53·0 (sd 11·9) years. Among the 864 patients who had serum folate measurements, the median serum folate concentration was 6·90 (25th–75th percentile: 5·22–9·20) ng/ml. Nearly half of the patients (46·3 %) were at an advanced stage (i.e. Barcelona Clinic Liver Cancer stage ≥C). Hepatic resection/liver transplantation was the most common tumour treatment (44·0 %), followed by hepatic artery intervention (39·6 %) and local ablation (11·3 %) (Table 1). The distribution of the six selected SNP in the folate metabolism pathway genes is presented in Table 2. The minor allele frequencies of these six SNP ranged between 12·1 % and 28·4 % in HCC patients, which were similar to those in the general population except for the T allele frequency of MTHFR rs1801133 (25·2 % v. 38·6 %).
Table 1. Baseline characteristics of the included patients with hepatocellular carcinoma in the Guangdong liver cancer cohort study

Abbreviations: AFP, α-fetoprotein; BCLC, Barcelona Clinic Liver Cancer; BMI, body mass index; CRP, C-reactive protein; P25, the 25th percentile; P75, the 75th percentile, sd: standard deviation.
* A summary score of the number of abnormal laboratory-defined values for six liver function tests: alanine aminotransferase > 50 μ/l, aspartate aminotransferase > 40 μ/l, γ-glutaryl-transferase > 60 μ/l, alkaline phosphatase > 150 μ/l, albumin < 40 g/l and total bilirubin > 20.5 μmol/l, ranging from 0 to 6.
† Including radiation therapy and systemic treatment (e.g. molecular targeted therapy, systemic chemotherapy, traditional Chinese medication).
Table 2. Distribution of the selected SNPs in folate metabolism-related genes in the Guangdong liver cancer cohort study

Abbreviations: M, major allele; m, minor allele; MAF, minor allele frequency; MTHFR, methylenetetrahydrofolate reductase; MTR, methionine synthase; MTRR, methionine synthase reductase; SNP, single nucleotide polymorphism.
* Data were extracted from the National Center for Biotechnology Information (NCBI) Allele Frequency Aggregator (ALFA) for the East Asian population.
Baseline characteristics of HCC patients by the selected MTHFR, MTR and MTRR genotypes are shown in online Supplementary Tables 2 and 3, respectively. The MTHFR rs1801131 CC and MTHFR rs2274976 AA genotypes were more prevalent in women than in men. Patients carrying MTHFR rs1801133 TT, MTHFR rs1801131 CC and MTRR rs10380 TT genotypes tended to have lower serum folate levels, whereas patients with MTHFR rs2274976 AA, MTR rs1805087 GG and MTRR rs1801394 GG genotypes tended to have higher serum folate levels compared with their counterparts, although not statistically significant.
Methylenetetrahydrofolate reductase, 5-methyltetrahydrofolate-homocysteine methyltransferase reductase and 5-methyltetrahy hepatocellular carcinoma survival
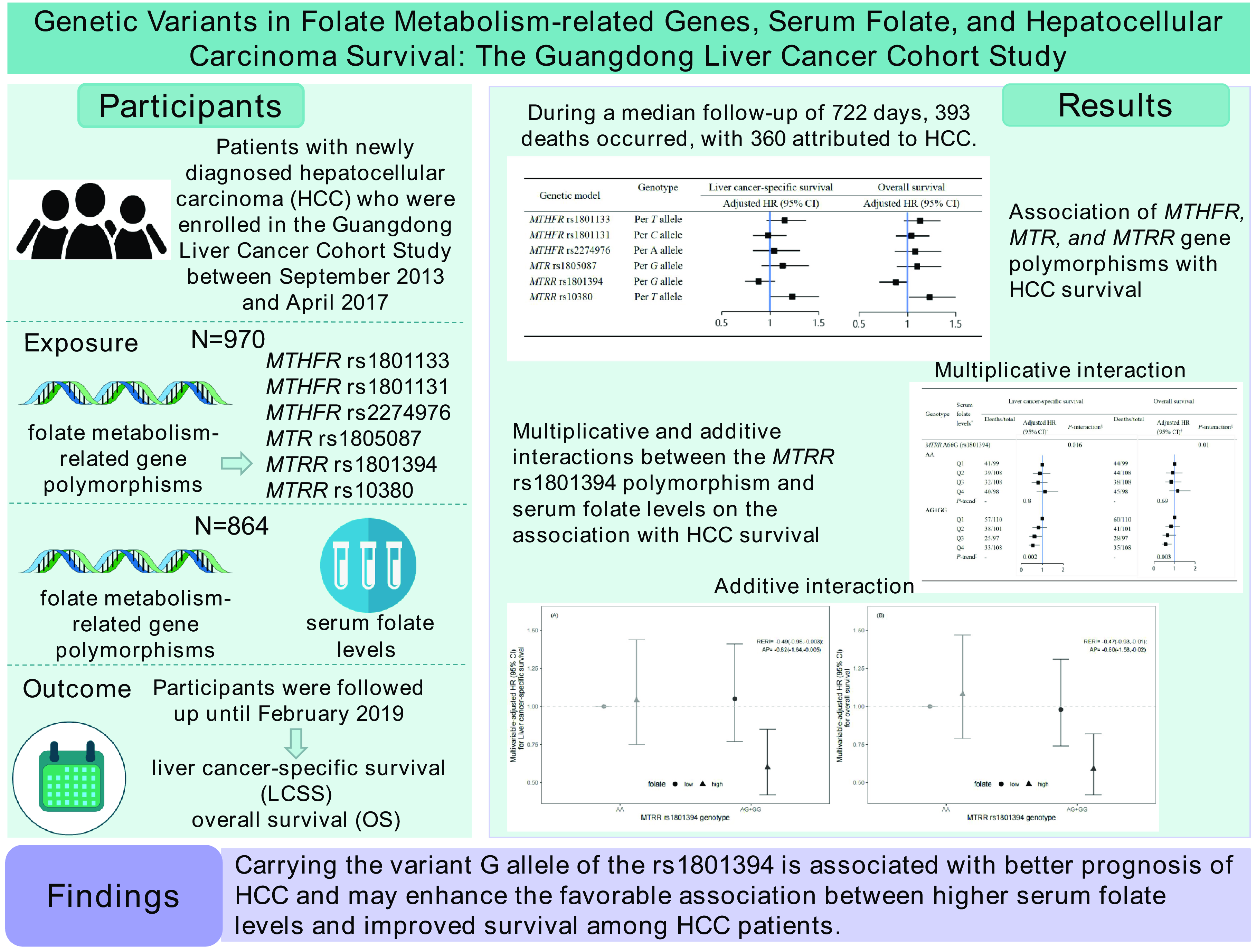
During a median follow-up of 722 (25th–75th percentile: 320–1077) days, 393 (40·5 %) patients were deceased and 360 (91·6 %) of them died from HCC. The associations of MTHFR, MTR and MTRR genetic polymorphisms with HCC survival are shown in Table 3. After adjustment for non-clinical and clinical prognostic factors, MTRR rs1801394 was associated with OS in HCC patients in the additive, co-dominant and dominant models. Compared with HCC patients with wild-type genotype (AA), HCC patients with GA (GA v. AA: HR = 0·77; 95 % CI: 0·62, 0·96) genotype exhibited improved OS. In the dominant model, the mutant patients with HCC (AG + GG) had better OS than patients with the wild-type genotype (AA) (HR = 0·78; 95 % CI: 0·62, 0·96). In addition, an increased number of the G allele was associated with improved OS (per G allele: HR = 0·84; 95 % CI: 0·71, 0·99), under the additive model. However, each additional mutant allele T of MTRR rs10380 was associated with worse OS among HCC patients (per T allele: HR = 1·23; 95 % CI: 1·01, 1·50) under the additive model after full adjustments. The other four SNP (MTHFR rs1801133, MTHFR rs1801131, MTHFR rs2274976 and MTR rs1805087) did not show significant associations with the survival outcomes of HCC patients. In sensitivity analyses, after restricting our analyses to male participants, the results were similar to those from the main analyses (online Supplementary Table 4).
Table 3. Association of MTHFR, MTR, and MTRR genetic polymorphisms with survival outcomes among patients with hepatocellular carcinoma in the Guangdong liver cancer cohort study

Abbreviations: CI, confidence interval; HR, hazard ratio; MTHFR, methylenetetrahydrofolate reductase; MTR, methionine synthase; MTRR, methionine synthase reductase; ref, reference.
* Crude model.
† Adjusted for age at diagnosis (continuous), sex (women, men), BMI (< 18.5, 18.5∼24.0, 24.0∼28.0, ≥ 28.0 kg/m2), smoking status (never, former, current).
‡ Additionally adjusted for α-fetoprotein levels (≤ 400 ng/ml, > 400 ng/ml), C-reactive protein levels (≤ 3.0 mg/l, > 3.0 mg/l), liver damage score (0, 1∼2, ≥ 3), BCLC stage (0, A, B, ≥C), and cancer treatment (hepatectomy/liver transplantation, local ablation, hepatic arterial intervention, other treatments).
When we combined the number of protective alleles, we observed that compared with patients having two to six protective alleles, patients carrying ten to twelve protective alleles had better LCSS (HR = 0·70; 95 % CI: 0·49, 1·00) and OS (HR = 0·67; 95 % CI: 0·47, 0·95) in fully-adjusted models (Table 4).
Table 4. Association of combined folate metabolism-related gene polymorphisms with survival outcomes among patients with hepatocellular carcinoma in the Guangdong liver cancer cohort study

Abbreviations: CI, confidence interval; HR, hazard ratio; ref, reference.
* Crude model.
† Adjusted for age at diagnosis (continuous), sex (women, men), BMI (< 18.5, 18.5∼24.0, 24.0∼28.0, ≥ 28.0 kg/m2), smoking status (never, former, current).
‡ Additionally adjusted for α-fetoprotein levels (≤ 400 ng/ml, > 400 ng/ml), C-reactive protein levels (≤ 3.0 mg/l, > 3.0 mg/l), liver damage score (0, 1∼2, ≥ 3), BCLC stage (0, A, B, ≥C), and cancer treatment (hepatectomy/liver transplantation, local ablation, hepatic arterial intervention, other treatments).
§ The C allele of the rs1801133 polymorphism, the A allele of the rs1801131 polymorphism, the G allele of the rs2274976 polymorphism, the A allele of the rs1805087 polymorphism, the G allele of the rs1801394 polymorphism, and the C allele of the rs10380 polymorphism were considered protective.
|| Linear trend was tested by entering the number of protective alleles as a continuous variable in the regression models.
Interaction between serum folate levels and methylenetetrahydrofolate reductase, 5-methyltetrahydrofolate-homocysteine methyltransferase reductase and 5-methyltetrahydrofolate-homocysteine methyltransferase genetic polymorphisms
In our previous study, patients in the lowest quartile had significantly worse LCSS (HR = 1·48; 95 % CI: 1·05, 2·09) and OS (HR = 1·43; 95 % CI: 1·03, 1·99) after adjustment for non-clinical and clinical prognostic factors compared with those in the third quartile of serum folate(Reference Fang, Liu and Liao5). As shown in Table 5, we further observed significant multiplicative interactions between sex-specific quartiles of serum folate concentrations and MTRR rs1801394 genotype on the association with LCSS (Pinteraction = 0·016) and OS (Pinteraction = 0·010). When stratified by the genotype of MTRR rs1801394, higher serum folate concentrations were associated with better LCSS (Q4 v. Q1: HR = 0·55; 95% CI: 0·35, 0·86; P = 0·006 for trend) and OS (Q4 v. Q1: HR = 0·56; 95% CI: 0·36, 0·86; P = 0·006 for trend) among patients who carried mutant genotypes (AG and GG), but not among those with the wild-type genotype (AA). No significant multiplicative interaction was observed between other MTHFR, MTR and MTRR SNPs, as well as the number of protective alleles, and serum folate levels (online Supplementary Table 5).
Table 5. Association between sex-specific quartiles of serum folate levels and liver cancer-specific and overall survival stratified by MTHFR, MTR and MTRR genetic polymorphisms

HR, hazard ratio; MTHFR, methylenetetrahydrofolate reductase; MTR, methionine synthase; MTRR, methionine synthase reductase; ref, reference; Q1, first quartile; Q2, second quartile; Q3, third quartile; Q4, fourth quartile.
* Sex-specific quartiles of serum folate levels: women: Q1: ≤ 6.65 ng/ml, Q2:6.65∼8.97 ng/ml, Q3:8.97∼11.00 ng/ml, Q4: > 11.00 ng/ml; men: Q1: ≤ 5.03 ng/ml, Q2:5.03∼6.70 ng/ml, Q3:6.70∼8.88 ng/ml, Q4: >8.88ng/ml.
† Adjusted for age at diagnosis (continuous), sex (women, men), BMI (< 18.5, 18.5∼24.0, 24.0∼28.0, ≥ 28.0 kg/m2), smoking status (never, former, current), α-fetoprotein level (≤ 400 ng/ml, > 400 ng/ml), C-reactive protein level (≤ 3.0 mg/l, > 3.0 mg/l), liver damage score (0, 1∼2, ≥3), BCLC stage (0, A, B and ≥C), and cancer treatment (hepatectomy/liver transplantation, local ablation, hepatic arterial intervention and other treatments).
‡ Test the trend against a variable containing the median of each interquartile.
§ The likelihood ratio test was used to evaluate the interaction term.
We documented significant additive interaction between serum folate levels and the MTRR rs1801394 genotype on the association with LCSS (RERI = −0·49; 95 % CI: −0·98, −0·003; AP = −0·82; 95 % CI: −1·64, −0·005) and OS (RERI = −0·47; 95 % CI: −0·93, −0·01; AP = −0·80; 95 % CI: −1·58, −0·02). The joint associations of serum folate levels and MTRR rs1801394 genotype on survival outcomes of HCC patients are shown in Fig. 2. After adjustment for non-clinical and clinical factors, patients with high serum folate levels (> 6·91 ng/ml) and heterozygous or homozygous variants (AG and GG) had better LCSS (HR = 0·60, 95 % CI: 0·42, 0·85) and OS (HR = 0·59, 95 % CI: 0·42, 0·82), compared with patients with low serum folate levels (≤ 6·91 ng/ml) and the wild-type genotype (AA).

Fig. 2. Joint effects of serum folate levels and genotype of MTRR rs1801394 on survival outcomes in the Guangdong Liver Cancer Cohort study. (A) Liver cancer-specific survival and (B) overall survival. AP, attributable proportion due to interaction; HR, hazard ratio; MTRR, 5-methyltetrahydrofolate-homocysteine methyltransferase reductase; RERI, relative excess risk due to interaction. The MTRR rs1801394 genotype was divided into wild-type (AA) or mutant (AG + GG) groups. Serum folate levels were classified into low (≤ 6·91 ng/ml) and high (> 6·91 ng/ml) based on the median, represented respectively by solid circles and triangles. Data were analysed by Cox proportional hazards models, adjusted for age at diagnosis (continuous), sex (women, men), BMI (<18·5, 18·5∼24·0, 24·0∼28·0, ≥ 28·0 kg/m2), smoking status (never, former, current), α-fetoprotein level (≤ 400 ng/ml, > 400 ng/ml), C-reactive protein level (≤ 3·0 mg/l, > 3·0 mg/l), liver damage score (0, 1∼2, ≥ 3), BCLC stage (0, A, B, ≥C), and cancer treatment (hepatectomy/liver transplantation, local ablation, hepatic arterial intervention, other treatments). Measures for additive interaction and the corresponding 95 % CIs were estimated using the delta method.
Discussion
We prospectively examined the association of six polymorphisms in three genes encoding folate metabolising enzymes (MTHFR, MTR or MTRR) and their interactions with serum folate concentrations with HCC survival in the GLCC study. Carrying the variant G allele of the MTRR rs1801394 polymorphism was related to improved OS among patients with HCC after adjusting for potential confounders. However, the MTHFR and MTR gene polymorphisms did not manifest any association with HCC survival. When combined with the effect of the six SNP effects, patients carrying more protective alleles had better survival. Additionally, we observed multiplicative and additive interactions between serum folate levels and the MTRR rs1801394 polymorphism. The association between serum folate concentrations and HCC survival differed by the genotypes of MTRR rs1801394 and was only evident among patients who carried mutant genotypes (AG and GG). Compared with patients with low serum folate levels and the wild-type genotype (AA), patients with high serum folate levels and mutant genotypes (AG and GG) had improved LCSS and OS.
MTRR is one of the key regulatory enzymes involved in the folate metabolism pathway. It can catalyse the regeneration of methylcobalamin, which is a cofactor of MTR in the remethylation of homocysteine to methionine, and the regeneration of tetrahydrofolate for nucleotide biosynthesis(Reference Han, Shen and Meng27). Mutations in the MTRR gene may decrease the affinity between MTR and MTRR and reduce the activity of MTR, leading to abnormal DNA synthesis, repair and methylation(Reference Fang, Ji and Fan28). As well, animal studies have suggested that inhibition of MTR activity suppresses the proliferation of tumour cells(Reference Ghergurovich, Xu and Wang12,Reference Sullivan, Darnell and Reilly13) . rs1801394 is the most common polymorphism in the MTRR gene, which causes the substitution of isoleucine with methionine at codon 22 in the MTRR, yielding a variant protein exhibiting fourfold lower enzyme activity than the wild-type protein in vivo(Reference Gunathilake, Kim and Lee29). Our results showed that the heterozygous or homozygous mutations (AG and GG) of MTRR rs1801394 exhibited a protective effect on the prognosis of HCC. Similarly, AG and GG carriers had a lower risk of recurrence in colorectal adenoma(Reference Hubner, Muir and Liu30) and prostate cancer(Reference Collin, Metcalfe and Refsum31) and better survival of liver cancer(Reference Wang, Lu and Ling14) and gastric cancer(Reference Zhao, Xu and Gu32,Reference Zhao, Gu and Xu33) than wild-type (AA) patients. The rs10380 polymorphism is also common in the MTRR gene. A case–control study conducted in China showed that individuals carrying the MTRR rs10380 TT genotype had a higher incidence of HCC than wild-type (CC)(Reference Zhang, Liu and Han34). In our study, we additionally observed that each additional variant allele T of MTRR rs10380 was associated with worse OS in HCC patients. However, no such associations were found in patients with non-small cell lung cancer(Reference Jin, Huang and Hu35,Reference Do, Choi and Lee36) and gastrointestinal cancer(Reference Morishita, Hishida and Okugawa37). A common variant in MTR rs1805087 leads to the substitution of aspartic acid with glycine, which may decrease the activity of MTR. However, our study, along with previous studies on liver(Reference Wang, Lu and Ling14), lung(Reference Jin, Huang and Hu35), stomach(Reference Zhao, Xu and Gu32,Reference Zhao, Gu and Xu33) and ovarian(Reference Dixon, Ibiebele and Protani38) cancers, found null associations between the MTR rs1805087 polymorphism and cancer survival.
Multiple studies have examined the interactions between the MTRR rs1801394 polymorphism and folate intake or status on cancer risk, but there is limited evidence on cancer prognosis. For instance, a case–control study conducted in Thailand found a stronger association between lower serum folate levels and a higher risk of developing colorectal cancer in individuals with the G allele (AG and GG) of the MTRR rs1801394 than in wild-type individuals(Reference Panprathip, Petmitr and Tungtrongchitr39). Another case–control study demonstrated that women with wild-type (AA) of MTRR rs1801394 and low dietary folate intake have an elevated risk of developing colorectal cancer compared to women with homozygous mutant type (GG) and low dietary folate intake(Reference Liu, Scherer and Poole40). Similarly, in a cohort of Japanese postmenopausal women, the MTRR rs1801394 GG genotype and lower folate intake were associated with a higher risk of breast cancer compared with adequate folate intake and wild-type genotype (AA)(Reference Suzuki, Matsuo and Hirose41). In a randomised controlled trial conducted in 546 patients with colorectal adenoma, the risk of recurrence significantly decreased in MTRR rs1801394 heterozygotes and homozygotes (AG and GG) who were treated with folic acid (500 μg/d), but not in those who did not receive folic acid(Reference Hubner, Muir and Liu30). Consistent with previous results, we observed a significant interaction between the MTRR rs1801394 variant and serum folate levels on both additive and multiplicative scales. Compared with the AA genotype carriers with low serum folate levels, individuals carrying the G allele (AG and GG) of the MTRR rs1801394 and with high serum folate levels (> 6·91 ng/ml) had better HCC survival. The association between higher serum folate levels and improved LCSS/OS was only restricted to patients with the AG and GG genotypes of MTRR rs1801394, but not the wild-type (AA) patients with HCC. Our findings suggest that HCC patients with MTRR rs1801394 mutations may benefit more from optimising folate status through diet and supplements.
MTHFR is a key enzyme in the folate metabolism pathway that carries out the irreversible conversion of 5,10-methylene THF to 5-methyl THF, which in turn directs the folate pool towards remethylation of homocysteine to methionine. Two common polymorphisms in the MTHFR genes, rs1801133 and rs1801131, have been related to the activity of the enzyme MTHFR and altered levels of DNA methylation and synthesis(Reference Weisberg, Tran and Christensen6,Reference Rozen7) , but the function of MTHFR rs2274976 remains unknown(Reference Rady, Szucs and Grady42). To the best of our knowledge, the impact of the MTHFR rs2274976 polymorphism on HCC prognosis remains unexplored, while the association between MTHFR rs1801133 and rs1801131 polymorphisms and HCC survival has yielded inconsistent results. For example, a cohort study reported that HCC patients carrying the MTHFR rs1801133 TT and CT genotypes had a higher rate of HCC recurrence compared with those with the CC genotype(Reference Wang, Xie and Lu18). However, another cohort study found that MTHFR rs1801133 TT and CT genotypes were associated with favourable survival in HCC patients(Reference Kuo, Huang and Kuo15), but no association was found between MTHFR rs1801131 polymorphism and HCC survival(Reference Peres, Galbiatti-Dias and Castanhole-Nunes19). Previous cohort studies also revealed an interaction between folate intake and MTHFR rs1801131 polymorphism on survival among patients with ovarian cancer(Reference Zhang, Liu and Hao43) or esophageal cancer(Reference Lu, Xie and Wang44,Reference Jing, Huang and Duan45) , where the association between high folate intake and better prognosis was only evident in individuals with the MTHFR rs1801133 CC genotype. In contrast, rs1801133, rs1801131 and rs2274976 polymorphisms in the MTHFR gene were not associated with survival among patients with HCC in our study. Moreover, the present study did not support that the association between serum folate levels and HCC survival differed by MTHFR rs1801133 and the other two MTHFR gene polymorphisms (rs1801131 and rs2274976). In agreement with our results, a follow-up study of 232 patients with HCC reported no significant interaction between the MTHFR rs1801133 polymorphism and RBC folate levels on survival(Reference Kuo, Huang and Kuo15). The primary circulating form of folate is 5-methyl THF, which is synthesised through the catalytic action of the MTHFR enzyme. The enzyme is responsible for regulating the conversion of folate derived from dietary and/or supplemental sources, thereby MTHFR polymorphisms potentially interact with folate intake rather than circulating folate levels, which may explain the discrepancy between studies.
The combined effect of multiple SNP may be more informative than the individual effect of a single SNP. As expected, HCC patients with over seven protective alleles related to folate metabolism had better OS and LCSS compared with patients carrying 2–6 protective alleles. Consistently, in a cohort study of patients with gastric cancer, the combination of the MTRR rs1801394 GA and MTR rs1805087 AA genotypes prolonged patient survival over the combination of the MTRR rs1801394 AA and MTR rs1805087 AA genotypes. However, no significant protective effect was found in patients with the MTRR rs1801394 GA and MTR rs1805087 GA genotypes(Reference Zhao, Gu and Xu33).
Several strengths of this study lend credibility to its findings. First, we performed a large prospective cohort study and included only patients with newly diagnosed HCC to minimise potential confounding. Second, we genotyped six specific SNP, including SNP (MTHFR rs2274976 and MTRR rs10380), not previously addressed in prognostic studies of liver cancer. In addition, we measured serum folate levels in the same population, which enabled us to investigate the individual and combined association of folate-metabolising genes (MTHFR, MTR or MTRR) polymorphisms and serum folate levels with HCC prognosis. Third, we extensively collected information on covariates, including demographics, lifestyle factors, clinical characteristics and cancer treatment. Controlling for these key prognostic factors helps minimise confounding biases. Lastly, we used both OS and LCSS as endpoints in our analyses, despite HCC being a highly lethal cancer. This provides a more comprehensive view of HCC prognosis in our study.
We also acknowledge several limitations in our study. First, we only measured serum folate concentrations at diagnosis of HCC. Changes in diet, lifestyles, cancer progression and treatments such as chemotherapy, which can inhibit the folate cycle, may affect circulating folate levels after diagnosis. Additionally, we did not have information on biomarkers of other one-carbon nutrients such as vitamin B2, vitamin B6 and vitamin B12. Moreover, we only detected six common variants in the folate-metabolising genes. However, other genetic variants in these genes could potentially affect the activity of enzymes involved in folate metabolism. Finally, all of our participants were Asian, so it may be challenging to generalise our findings to patients of different genetic backgrounds.
In conclusion, our results showed that the heterozygous or homozygous mutant genotypes of MTRR rs1801394, individually or in combination with higher serum folate concentrations, were associated with improved survival among patients with HCC, supporting the role of gene–environment interactions in HCC prognosis. Our study provides the possibility of precision folate intervention to improve the prognosis of HCC with specific folate metabolism genotypes. Future randomised clinical trials and experiments are warranted to confirm our findings and decipher the underlying mechanisms.
Acknowledgements
We thank all the study participants and research staff for their contributions and commitment to the present study.
This work was supported by the grants from National Natural Science Foundation of China (Grant number 81803219); the Natural Science Foundation of Guangdong Province, China (Grant number 2022A1515011744) and the Science and Technology Program of Guangzhou, China (Grant number 202201011485).
The authors’ contributions were as follows – Y. Z., H. Z. and A. F.: conceived and designed the study; Y. L., X. D., M. Z., T. H., Z. Y., J. S., P. T., Z. L., Z. Z., Q. L., Y. X., X. X., T. P. and J. L.: collected and cleared the data; Y. L. and X. D.: analysed and interpreted the data; Y. L.: wrote the paper; Y. L., A. F.: revised the manuscript; X. D., M. Z., J. S., P. T., X. Z., E. L. G. and A. F.: critically reviewed the manuscript; Y. Z., H. Z. and A. F.: had primary responsibility for final content and all authors: read and approved the final manuscript.
All authors declare no potential conflicts of interest.
The data are available from the corresponding author on reasonable request.
Supplementary material
For supplementary material/s referred to in this article, please visit https://doi.org/10.1017/S0007114524001776









