


Introduction
Larks in the genus Heteromirafra are of particular interest to conservationists because the genus may be ancient and basal to all larks (de Juana et al. Reference de Juana, Suárez, Ryan, del Hoyo, Elliott and Christie2004) and because all the species in this tiny genus are globally threatened. Rudd's Lark H. ruddi (‘Vulnerable’) is endemic to a small area of upland grassland in South Africa, while Archer's Lark H. archeri (‘Critically Endangered’) is known with certainty only from a single site in north-west Somalia, where it has not been seen since 1922 (BirdLife International 2009, Maphisa et al. Reference Maphisa, Donald, Buchanan and Ryan2009).
Liben Lark H. sidamoensis is also listed by IUCN as ‘Critically Endangered’ and is known only from a single site in south-east Ethiopia, the Liben Plain, where the first specimen was collected in 1968 (Erard Reference Erard1975, Collar et al. Reference Collar, Dellelegn Abebe, Fishpool, Gabremichael, Spottiswoode and Wondafrash2008, BirdLife International 2009). Formerly known as Sidamo Lark, the common name of the species was changed by BirdLife International to Liben Lark in 2009, to reflect more accurately the species's distribution (the Liben Plain no longer falls within the administrative region of Sidamo) and to help engender local support for its conservation (Collar Reference Collar2009). The Liben Plain forms part of a well-defined plateau within the Borana rangelands, once one of the most productive pastures in Africa. One of the main changes in the management of the Liben Plain in recent years has been the loss of the traditional ‘geda’ pattern of seasonal grazing, which has been implicated in sharp declines in wildlife elsewhere in eastern Africa (Western et al. Reference Western, Groom and Worden2009). Geda management involved transhumance, the use of wet- and dry-season pasturing and, since the loss of natural browsers, the control of encroaching scrub by fire. Instead, government initiatives have encouraged permanent settlement and the loss of shifting grazing.
When the Liben Lark was recognised as a new species, it was considered threatened, but its precise status was unclear and it was given the old IUCN category ‘Indeterminate’ (Collar and Stuart Reference Collar and Stuart1985, Collar Reference Collar1997). Evidence that habitat on the Liben Plain had been affected by human activity (Ash and Gullick Reference Ash and Gullick1989) led to the listing in 1994 of Liben Lark in the new IUCN category ‘Endangered’ (Collar et al. Reference Collar, Crosby and Stattersfield1994). This coincided with the first-ever observation of the species alive in the field, along with some reassuring evidence that the human pressures were no longer so pressing (Robertson Reference Robertson1995). Thereafter, the species became one of many for which little information existed but where no particularly strong threat was recognised, and it was down-listed to ‘Vulnerable’ (Stattersfield and Capper Reference Stattersfield and Capper2000).
Only following visits in 2006 by CNS did it become apparent that the species required serious study and rapid re-evaluation, leading to a series of surveys in 2007 and 2008 (Collar et al. Reference Collar, Dellelegn Abebe, Fishpool, Gabremichael, Spottiswoode and Wondafrash2008, Spottiswoode et al. Reference Spottiswoode, Wondafrash, Gabremichael, Dellelegn Abebe, Mwangi, Collar and Dolman2009) and 2009 (this paper). What little is known of the species history, ecology and behaviour was clarified and summarised by Collar et al. (Reference Collar, Dellelegn Abebe, Fishpool, Gabremichael, Spottiswoode and Wondafrash2008). Recent assessments based on a transect survey in 2007, supplemented by observations in 2008, suggested that the Liben Plain then held 90–250 individuals, perhaps with a strongly male-skewed adult sex ratio (Spottiswoode et al. Reference Spottiswoode, Wondafrash, Gabremichael, Dellelegn Abebe, Mwangi, Collar and Dolman2009). Birds selected areas with higher than average grass cover and height, and the main threats identified were overgrazing and encroachment by scrub and cropland (Spottiswoode et al. Reference Spottiswoode, Wondafrash, Gabremichael, Dellelegn Abebe, Mwangi, Collar and Dolman2009). Because of its small and declining population and range, the species was duly listed as ‘Critically Endangered’ in 2009 (BirdLife International 2009).
This paper presents the results of an expedition to the Liben Plain in late May 2009 to undertake a repeat of the transect survey of June 2007 and thus to assess changes in numbers and habitat. It also presents an assessment of the potential range of the species elsewhere in the Horn of Africa using spatial modelling based on the location of birds recorded in 2007.
Methods
Field transects
For the 2007 survey (11–16 June), GPS waypoints were taken at the start and end of each of 12 transects covering the grassland parts of the Liben Plain and every 250 m along them, to guide fieldworkers along each transect in a straight line and to provide sampling points for field measurements of vegetation (Figure 1). The transects totalled 60 km in length (Spottiswoode et al. Reference Spottiswoode, Wondafrash, Gabremichael, Dellelegn Abebe, Mwangi, Collar and Dolman2009). GPS locations were also taken of the location of each bird encountered in 2007, and vegetation measurements were taken at those locations in the same way as at transect waypoints. When a bird was sighted during a transect, observers left the transect and walked to where the bird was first seen to record its position with a GPS and take habitat measurements. The perpendicular distance of each bird from the transect was estimated from GPS locations. The behaviour of each bird was assessed at the time of first observation and all birds seen in song were assumed to be males.

Figure 1. Map of the Liben Plain, southern Ethiopia, showing the distribution of Liben Lark sightings in 2007 and 2009, transect waypoints and the main habitat types. This map is adapted from Figure 1 of Spottiswoode et al. (Reference Spottiswoode, Wondafrash, Gabremichael, Dellelegn Abebe, Mwangi, Collar and Dolman2009), in which the area of grassland was shown incorrectly due to a printing error.
In 2009, we repeated all transects walked in 2007 and took vegetation measurements in exactly the same way at (i) all vegetation-recording waypoints along each transect, (ii) all locations where birds had been recorded in 2007 and (iii) all locations where birds were recorded in 2009. Within 25 m of each point, counts were made of trees, fennel Ferula communis plants, paths or tracks and the number of nests (indicated by large, roughly circular patches of bare ground) of the Harvester Ant Messor cephalotes. The latter measurements were taken because the number of animal tracks and ant nests might be expected to influence grass height. Then the cover of shrubs, cow dung and bare ground was estimated within two 5-m radius circles located 10 m each side of the point and perpendicular to the axis of the transect. The height of the grass in these areas was estimated as the percentage cover of grass less than 5 cm tall, grass 5–15 cm tall, grass 15–40 cm tall and grass over 40 cm tall. These percentages summed to 100. Spottiswoode et al. (Reference Spottiswoode, Wondafrash, Gabremichael, Dellelegn Abebe, Mwangi, Collar and Dolman2009) contains further details. A single index of grass height was then estimated as the sum of the percentage covers for each grass height category multiplied by the mid-point of that grass height range (or in the case of the > 40 cm category, a value of 40). Because of changes in the way that trees and shrubs were counted in 2009, it was not possible to make a statistical comparison of changes in their occurrence between surveys, although photographic evidence suggested little difference.
Fieldwork in 2009 was undertaken by all the authors except GMB and CNS. Five of the seven fieldworkers in 2009 were also part of the 2007 expedition, ensuring a high degree of consistency in field recording methods between the two surveys. Fieldwork was carried out between 19 and 23 May 2009 by up to four teams of two or three observers working simultaneously. The drought that affected parts of eastern Africa in 2009 did not affect this region, and indeed heavy rain was recorded several times during the surveys. Transects started at around 06h30 and ended usually well before 11h00.
Sign tests were used to assess changes in other habitat variables between 2007 and 2009 at transect waypoints and at locations where the bird was recorded in 2007. Binary logistic regression was used to model differences in vegetation between locations where birds were recorded in 2009 and transect waypoints. Univariate analyses identified a set of candidate variables for entry into a multivariate model. Backwards deletion was used to derive the Minimum Adequate Model.
Assessing changes in NDVI on the Liben Plain
To assess whether any changes in grassland structure or bird numbers could be explained by changes in growing season or drought, the Normalised Difference Vegetation Index (NDVI) was extracted for the area of the Liben Plain for 1998 to 2009. On grasslands, variation in NDVI primarily reflects growing season, which depends largely on variation in rainfall. The original data were downloaded from http://free.vgt.vito.be and processed in Erdas Imagine and ArcMap. Average NDVI for the period April to June across the Liben Plain was calculated to describe inter-annual variation during the main rainfall and growing season and what is thought to be the breeding season of Liben Lark.
Range modelling with remote sensing
Maximum entropy modelling was used to predict the likelihood of occurrence of Liben Lark in Ethiopia as a function of land cover and altitude using Maxent 2.3 (Phillips et al. Reference Phillips, Anderson and Schapire2006). Maximum entropy modelling produces an output map indicating the likelihood of occupancy in each cell (higher is more likely). This is derived from a probability distribution produced using environmental predictor variables and presence-only data. It generates a user-defined number of pseudo-absences: 10,000 in this study. Probability distributions are produced from functions of the input variables, with the probability distribution that is most spread out or closest to uniform being selected at each stage. Through a process of machine learning, the final probability distribution that is closest to uniform (maximum entropy) is estimated. Land cover characteristics were derived from information collected by the SPOT–Vegetation sensor. This covers the planet daily, and collects data at 1-km resolution. The maximal NDVI values over 10-day (dekad) periods for 2007 were extracted from http://free.vgt.vito.be. The 36 images that cover the entire area were reduced using Principal Components Analysis (PCA) in Erdas Imagine 8.7, with the top eight components being used in further analysis. Topographic data were derived from 90 m Shuttle Radar Topography Mission SRTM (USGS 2004), 30 arc seconds (Global Land Cover Facility, University of Maryland, College Park, Maryland, February 2000; www.landcover.org). The first eight PCA components explained some 88% of the variation in the data. Together with altitude, these variables were entered into a maximum entropy model covering some 320,000 km2 area around the neighbourhood of the 1-km grid squares which contained at least one sighting of a Liben Lark. The model with the greatest gain was then identified (6.59) and this was used to predict the likelihood of occurrence of the species across a much wider area covering the whole of Ethiopia, clamping values in the new area to values within the model development area. The exercise was repeated without altitude in the model to assess whether the effect of elevation merely reflected variation in climate or whether it had additional explanatory power.
Results
Changes in vegetation since 2007
Of the 252 transect waypoints at which habitat measurements were taken in 2007, 21 (8.3%) were lost to agriculture by 2009. Changes in vegetation at the remaining 231 waypoints within the area of grassland are summarised in Table 1. There was a significant shortening of the average grass height between 2007 and 2009 and an increase in grass in the smallest height category (Table 1, Figure 2). The significant increase in the number of fennel plants resulted from both an increase of 9% (to 53%) in the number of points at which fennel was present and an increase in the number of plants counted at waypoints where fennel was already present in 2007 (sign test, P < 0.001).
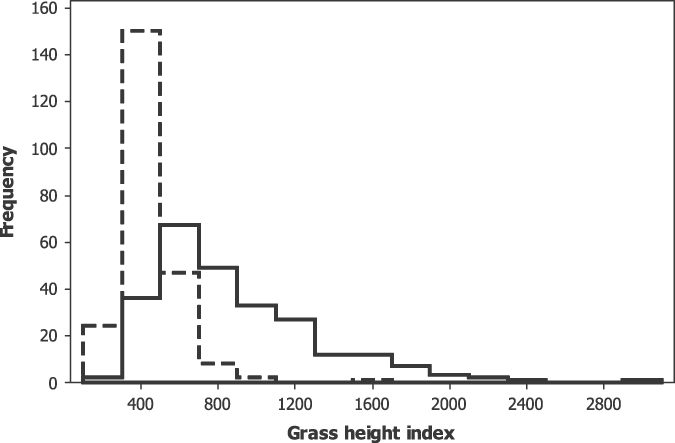
Figure 2. Frequency histograms of grass height index on transect waypoints. Dashed line, 2009; solid line, 2007.
Table 1. Changes in habitat variables between 2007 and 2009 at 231 transect waypoints within grassland habitat. Values shown are means.

Habitat use by birds
Points at which Liben Larks were recorded in 2007 differed from transect waypoints in having less bare ground and longer grass (Spottiswoode et al. Reference Spottiswoode, Wondafrash, Gabremichael, Dellelegn Abebe, Mwangi, Collar and Dolman2009). However, by 2009 these two sets of points were indistinguishable (Mann-Whitney U-tests; P > 0.5; Figure 2). Moreover, there was a significantly more rapid increase between 2007 and 2009 in cover of bare ground at sites occupied in 2007 than at transect waypoints (Mann-Whitney U-test, P < 0.001), indicating more rapid grassland degradation at sites occupied by birds in 2007. The results of logistic regression models are given in Table 2. In univariate analyses, only the amount of bare ground and the number of trees and bushes explained significant variation in the presence or absence of Liben Larks in 2009, both being negatively correlated with presence. When entered into a model simultaneously, both retained significant explanatory power (Table 2) and the entry of no other variable into this model improved its fit. The final model correctly assigned 75% of locations with respect to the presence or absence of birds.
Table 2. Univariate and multivariate binary logistic regression models of presence/absence of Liben Lark from 40 recorded locations of birds and 252 transect waypoints in May 2009.

Changes in number and distribution of birds
Transects in 2007 recorded Liben Larks at 69 point locations (Spottiswoode et al. Reference Spottiswoode, Wondafrash, Gabremichael, Dellelegn Abebe, Mwangi, Collar and Dolman2009). Of these, six (8.7%) locations had been lost to arable agriculture by 2009. In 2009, the number of sightings made along the same transects fell to 40 and the polygon encompassing all point locations fell from 44.6 km2 in 2007 to 27.8 km2 in 2009 (Figure 3). All but two encounters were with single singing males. On two occasions, two birds were seen together and in one of these, copulation was seen, confirming a breeding pair. No nests were found and no other evidence of breeding (e.g. carrying of food or nesting material) was seen.

Figure 3. Boxplots of percentage of bare ground recorded in June 2007 and May 2009 at points where birds were present in 2007 (‘Bird 2007’), points where birds were present in 2009 (‘Bird 2009’) and at the same set of vegetation sampling waypoints along transects in both years (‘Waypoint’). The horizontal line represents the median, the box the interquartile range and the vertical lines span the range of the values lying between the interquartile and 1.5 times the interquartile range. Outliers beyond this are represented as asterisks.
Recent changes in NDVI on the Liben Plain
There was no evidence of any change in the growing season on the Liben Plain between 1998 and 2009 (Figure 4), suggesting that the observed grassland deterioration could not be explained by patterns in rainfall. NDVI in 2009 was slightly higher than that in 2007.

Figure 4. Trends in average (with SD) April–June NDVI on the Liben Plain, 1998–2009.
Predicted distribution
The maximum entropy model predicting the probability of occupancy of 1-km cells by Liben Lark performed well, with an AUC/ROC of 1, indicating that all locations where birds were recorded and the 10,000 random points were correctly classified as occupied or unoccupied across the 320,000 km2 training area (Figure 4). This high ROC value will, to some extent, be an overestimate of the accuracy of the model due to the high degree of spatial autocorrelation of the points (Veloz Reference Veloz2009).
Using the equal sensitivity and selectivity threshold (37.6), only areas around the known distribution were classified as having a sufficiently high probability of being suitable for identification as occupied, indicating that there are no suitable areas near the existing population. It is worth noting, however, that all of the observations came from a very small area, meaning that spatial autocorrelation may be a problem. When the model was applied to the wider area, an area due east of Jijiga in eastern Ethiopia some 500 km north-east of the Liben Plain stood out as being the only extensive area of habitat that may be suitable, but the predicted probability of occupancy for this area was lower than the Liben Plain (Figure 5). The predicted distribution based on a model without altitude resulted in a projected range almost the same as that with altitude and the AUC was unchanged, suggesting that the reflectance characteristics of the Liben Plain are not found elsewhere in southern Ethiopia at any altitude.

Figure 5. Distribution of potential Liben Lark habitat, based on NDVI and altitude using maximum entropy modelling. Darker browns indicate greater probability of habitat being suitable. The 1,000 m (light grey), 1,500 m (grey) and 2,000 m (dark grey) contours are shown, along with country borders. The box indicates the area over which the model was built.
Discussion
The results suggest a rapid deterioration in habitat quality and population of the Liben Lark in just two years, confirming the diagnosis that the species is poised to become mainland Africa's first recorded bird extinction (Spottiswoode et al. Reference Spottiswoode, Wondafrash, Gabremichael, Dellelegn Abebe, Mwangi, Collar and Dolman2009). The results suggest that the continuing degradation of the grassland on the Liben Plain cannot be attributed to drought and that overgrazing is the most likely cause. There is a possibility that the population results are biased if more birds nest in May (month of the 2009 survey) than in June (2007 survey), since in other larks, males sing less when females are incubating and brooding (Donald Reference Donald2004). Nevertheless, the results in both years of study revealed a negative association between Liben Lark and the cover of bare ground, and the cover of bare ground increased significantly between 2007 and 2009.
The 2007 result showing a preference for taller grass was not replicated in 2009, possibly because the grass on the plain in 2009 was almost uniformly short (Figure 1). This cannot be attributed to drought, as in the months before the surveys, and indeed on several days during them, there was heavy rain, and NDVI changed little between 1998 and 2009. Early observations of the lark describe its habitat as ‘long grass’ or ‘waist-high grass’ (Collar et al. Reference Collar, Dellelegn Abebe, Fishpool, Gabremichael, Spottiswoode and Wondafrash2008), a habitat not seen anywhere on the Liben Plain in 2007–2009. Discussions with local people provided support for suggestions that the species prefers longer and denser grass than is currently available. Even relatively recently, the grassland on the plain was very much denser and taller than it is now; some local people reported that in their childhoods the grass was tall enough to bind together in situ to make shelters from the rain. The only site on the Liben Plain that was exclosed to grazers, and so where the structure of ungrazed grassland could be assessed, was a fenced army camp. In June 2008, this was unoccupied and the grass within it was considerably higher and more extensive than the grass on the common grazing land outside it (Figure 6).

Figure 6. Grassland structure on the Liben Plain in June 2008. Above, grazed common land showing the short sward height and patchy grass cover. Below, the taller and more continuous grassland cover of an exclosure (a fenced but at the time disused army camp). (Nigel J. Collar)
A further clue to the historically preferred habitat of the species lies in its morphology. It has a long hindclaw, a feature of larks using grassy habitats (Green et al. Reference Green, Barnes and Brooke2009), as well as long legs and a long neck. In contrast, the Somali Short-toed Lark Calandrella somalica, which is far more abundant than Liben Lark on the Liben Plain, has (as its name suggests) the short claws of species associated with bare ground. Grassland changes that have brought the Liben Lark to the verge of extinction may have favoured the far more common and widespread Somali Short-toed Lark.
Most of the bare ground recorded on the plain was not in large patches but interstitial within the grassland, which largely comprised a mosaic of clumps of grass in a matrix of bare ground. This is likely to be the result of surface erosion brought about by overgrazing. An increase in the extent of bare ground has been implicated in the local loss of the congeneric Rudd's Lark in South Africa (Maphisa et al. Reference Maphisa, Donald, Buchanan and Ryan2009). The loss of an annual burning regime might also have affected the Liben Lark, since Rudd's Lark shows a preference for lightly grazed grass that has been recently burned; nesting starts earlier and the breeding season is longer on grassland burned shortly before the start of the breeding season (Maphisa et al. Reference Maphisa, Donald, Buchanan and Ryan2009). Whether in Rudd's Lark this response is linked to habitat structure, to the control of ticks (Ixodidae) or to changes in other invertebrate populations is unclear. Burning encourages rapid regrowth and so might promote the development of longer grass at the start of the breeding season. It also creates a mosaic of burned and unburned patches, potentially offering a wider range of resources to grassland birds.
As with the 2007 survey, the great majority of birds encountered were males in song; indeed only one certain female was seen in 2009. It is inevitably the case that singing males are more detectable to observers than females, and particularly so in larks (Suarez et al. Reference Suarez, Garcia, Carriles, Calero-Riestra, Agirre, Justribo and Garza2009). However, observers followed a number of male birds for long periods without seeing them encounter a female. It is possible that males greatly outnumber females, a common pattern in endangered species (Donald Reference Donald2007), including other threatened larks (Donald et al. Reference Donald, de Ponte, Pitta Groz and Taylor2003, Suarez et al. Reference Brooke, Flower and Mainwaring2009, Brooke et al. Reference Suarez, Garcia, Carriles, Calero-Riestra, Agirre, Justribo and Garza2010). This means that the functional population is likely to be well below that assessed from the number of singing males.
The modelled prediction of suitable habitat in eastern Ethiopia (Figure 5) is remarkable for two reasons. First, the area in question is adjacent to the only certain locality for Archer's Lark (albeit not seen there since 1922), on the border of Somalia and Ethiopia. Second, Heteromirafra larks have twice been seen in a small area of grassland east of Jijiga in recent years (H. Shirihai in litt., MNG pers. obs.) some 30 km from the area predicted by the model. The possibility that Archer's Lark and Liben Lark are conspecific is currently being investigated; the fact that the Jijiga birds sang in flight like Liben Larks (MNG pers. obs.) tends to reinforce this possibility. Unfortunately, this area is now extremely dangerous even for Ethiopians and is likely to remain so for many years, so the determination of the identity of the larks there may not come soon. In the meantime, it is possible that the same problems of habitat degradation in play on the Liben Plain will eliminate this population. Conservationists therefore have no option but to treat the Liben Plain as though it were the only site in the world for the Liben Lark (as it is likely to be). In this regard, the predictive map reinforces the urgency of the situation, since it has been validated by the Jijiga sightings and it identifies no other places where suitable conditions exist.
Very rapid action is therefore required to prevent mainland Africa's first known bird extinction for at least five centuries. The results of this research suggest that increasing the cover of grass might be more important than increasing its height, although methods to increase one are likely also to increase the other. As a priority, substantial areas of the Liben Plain need to be closed off to grazing to promote grassland recovery, provide suitable feeding and breeding habitat for the lark and reduce surface soil erosion. Exclosures will need to be monitored carefully to assess whether exclosure alone is sufficient to restore the grassland, or whether ploughing to reduce soil compaction and re-seeding are also necessary. This measure might require temporary compensation for local grazers for the pastures lost but already enjoys strong local support from a committee of local stakeholders, who in May 2009 unanimously requested the help of international conservation NGOs to achieve this aim. They are concerned that further grassland degradation will result in the loss of the traditional pastoralist way of life, a process already started in the spread of arable agriculture onto the Liben Plain. A second aim of a conservation strategy for this species is to clear encroaching acacia thorn scrub and so increase the area of land available for both birds and grazing livestock. Recent scrub clearance near the plain suggests that large-scale scrub clearance is likely to be practical and relatively inexpensive.
Acknowledgements
We thank BirdLife International's Preventing Extinctions Programme for covering the costs of the 2009 travel and fieldwork, supplemented by the British Ecological Society (MAKM) and the Bromley Trust (NJC); Tony Hickey of Ethiopian Quadrants for generous support; Abiy Dagne and Tewabe Ashenafi for excellent service as our drivers; Paul Dolman for advice on fieldwork; and Hadoram Shirihai for kindly sharing information. We are very grateful to Ian May for producing Figure 1 and for estimating the areas of the two polygons of occupancy. For helpful comments on earlier drafts we are most grateful to Rhys Green, Nathan Gregory, Paola Laiolo and an anonymous referee.








