


Introduction
The poorly known Black-fronted Francolin Pternistis atrifrons (“Black-fronted Spurfowl” following Crowe & Little Reference Crowe and Little2004), treated as a subspecies of the Chestnut-naped Francolin P. castaneicollis for a long time, is currently considered a species in its own right based on morphological and genetic characteristics (del Hoyo et al. Reference del Hoyo, Collar, Christie, Elliott and Fishpool2014, Töpfer et al. Reference Töpfer, Podsiadlowski and Gedeon2014). This classification was adopted in the most recent IUCN Red List of Threatened Species (BirdLife International 2016). The range of the Black-fronted Francolin is restricted to a small area of the Oromia Regional State in southern Ethiopia, with only one historic record from neighbouring Kenya (Töpfer et al. Reference Töpfer, Podsiadlowski and Gedeon2014). In the absence of adequate information on the geographic range and the population size, it was placed in the category Least Concern (BirdLife International 2016). Our own field studies in 2012 and 2013 (Töpfer and Gedeon Reference Töpfer and Gedeon2013, Töpfer et al. Reference Töpfer, Podsiadlowski and Gedeon2014, Gedeon et al. Reference Gedeon, Zewdie and Töpfer2017), however, have already indicated major threats (range and habitat losses due to agricultural expansion, grazing pressure, commercial firewood and timber exploitation, and hunting) but lacked more precise information on range and population size. Therefore, a new survey was performed in 2014 to collect additional data to evaluate the Black-fronted Francolin’s current conservation status and to develop proposals for conservation actions, including the expansion of the currently projected Borana National Park.
Methods
The present paper is based mainly on field data collected from 5 to 25 June 2014 by Kai Gedeon but also includes information (12 records) from our previous studies in 2012 and 2013 (Töpfer et al. Reference Töpfer, Podsiadlowski and Gedeon2014). In 2014, evidence for the occurrence of francolins (21 records) was collected along a 1,128-km track in the mountainous area between Yabello and Moyale, using an off-road vehicle and walking (Figure 1). These records comprise our own sight records and documentation of species-specific calls, pieces of characteristically shaped droppings or typically patterned moulted feathers (cf. Töpfer et al. Reference Töpfer, Podsiadlowski and Gedeon2014) as well as credible information by local villagers. The latter was obtained by asking for the occurrence of any francolin species in the area by presenting plates in a field guide (Redman et al. Reference Redman, Stevenson and Fanshawe2009) without prior mention of the target species. We considered such information credible if villagers could identify Black-fronted Francolins unequivocally, particularly with respect to its closest relative, the Chestnut-naped Francolin depicted on the same plate (Töpfer et al. Reference Töpfer, Podsiadlowski and Gedeon2014). In addition, permanent sound recording devices (Wildlife Acoustics SongMeter SM2+) were set up at different subjectively chosen sites.
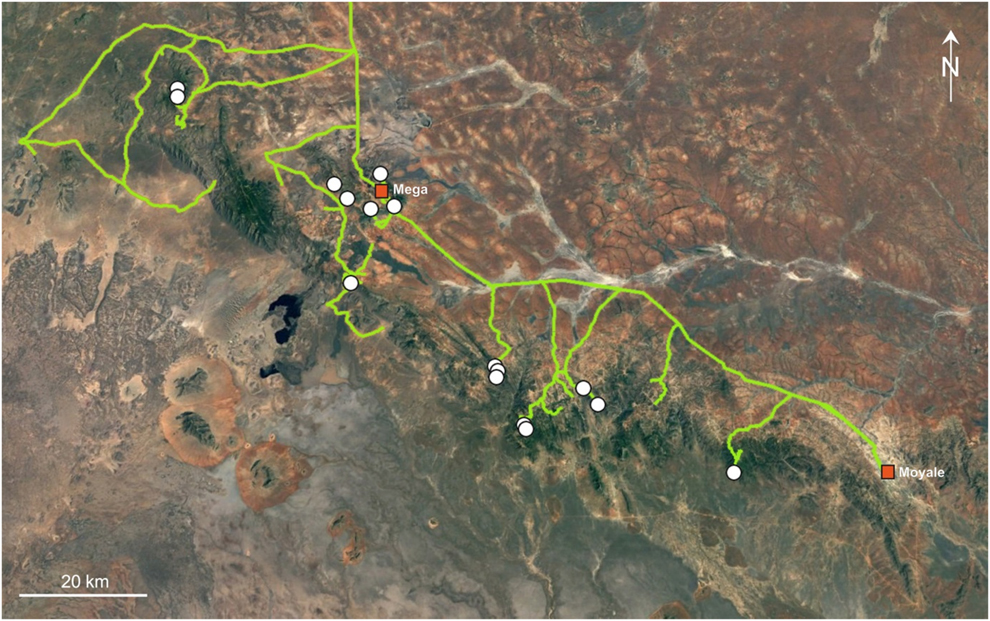
Figure 1. The 1,128-km survey track (light line) and Black-fronted Francolin records (white dots) within and around the mountains between Dilo, Mega and Moyale in Southern Ethiopia (June 2014). Source: Google Earth.
For model building we compiled a comprehensive set of species records from our own field surveys. In total, 26 unique georeferenced localities were available. We obtained 19 bioclimatic variables from the Worldclim database (Hijmans et al. Reference Hijmans, Cameron, Parra, Jones and Jarvis2005) with a spatial resolution of 30 arc seconds. The temporal resolution of the Worldclim data corresponds to monthly averages between 1950 and 2000. Additionally, as remote sensing variables may allow incorporation of habitat availability in SDMs, we used three variables obtained via a tasseled cap transformation (Crist and Cicone Reference Crist and Cicone1984) provided by the United States Geological Survey (USGS) in cooperation with the National Aeronautics and Space Administration (NASA), and ESRI product Tasseled Cap – 2010 Mature Support (ESRI 2016). This variable set originally had a resolution of 0.00026949459 by 0.00026949459 decimal degrees and was resampled to 30 arc seconds to match the resolution of the climate data. The tasseled cap transformation allows the computation of the three indices, brightness, greenness and wetness, which differentiate between open landscape, vegetation cover and moisture. The final set of variables was selected based on a matrix of pairwise Spearman rank correlations, wherein only variables with an inter-correlation of R2 < 0.75 were selected (i.e. in highly correlated pairs of variables only one variable was selected). These comprised BIO1 = Annual Mean Temperature, BIO3 = Isothermality, BIO4 = Temperature Seasonality, BIO7 = Temperature Annual Range, BIO12 = Annual Precipitation, BIO13 = Precipitation of Wettest Month, BIO15 = Precipitation Seasonality, BIO18 = Precipitation of Warmest Quarter, TC1 = brightness, TC2 = greenness and TC3 = wetness. For visualizing the final results we obtained a Landsat GLS picture with a spatial resolution of 30 m from the ESRI image service. WGS84 was used as the coordinate system.
For SDM computation we selected Maxent 3.3.3k (Phillips et al. Reference Phillips, Anderson and Schapire2006; Elith et al. Reference Elith, Phillips, Hastie, Dudik, Chee and Yates2011) as this algorithm performs well even if the number of records is rather restricted (Elith et al. Reference Elith, Graham, Anderson, Dudik, Ferrier, Guisan, Hijmans, Huettmann, Leathwick, Lehmann, Li, Lohmann, Loiselle, Manion, Moritz, Nakamura, Nakazawa, Overton, Perterson, Phillips, Richardson, Scachetti-Pereira, Shapire, Soberón, Williams, Wisz and Zimmermann2006). Maxent is a machine learning algorithm which estimates the probability of environmental suitability of a given area based on comparisons between the conditions at actual species occurrences and random background data (Elith et al. Reference Elith, Phillips, Hastie, Dudik, Chee and Yates2011). As background we selected the general area of occurrence of the species within a rectangular area. To avoid over-complex models we restricted the feature types to linear, quadratic and hinge features, while using the default settings for the other options. As output we chose the logistic output format. For model testing via the area under the receiver operating characteristic curve (AUC; Swets Reference Swets1988), we randomly split the species records 100 times into 80% used for model training and 20% used for model evaluation. The average prediction across all replicates was used for final processing, which was reclassified omitting all grid cells with probabilities below the average minimum training presence as non-fixed threshold as recommended by Liu et al. (Reference Liu, Berry, Dawson and Pearson2005).
Results
Geographic range
The data confirm that the range of the Black-fronted Francolin is restricted to a very small area in the extreme south of Ethiopia. The northern- and westernmost record is at 4°18´49´´N 37°58´50´´, the southern- and easternmost record at 3°33´38´´N 38°49´47´´E (Figure 2). Thus, the north-south extent of the area is about 90 km, and east-west about 100 km. The modelled distribution of the Black-fronted Francolin indicates its close dependency on the mountain range running from Dilo and Mega in Ethiopia up to the Kenyan border near Moyale.
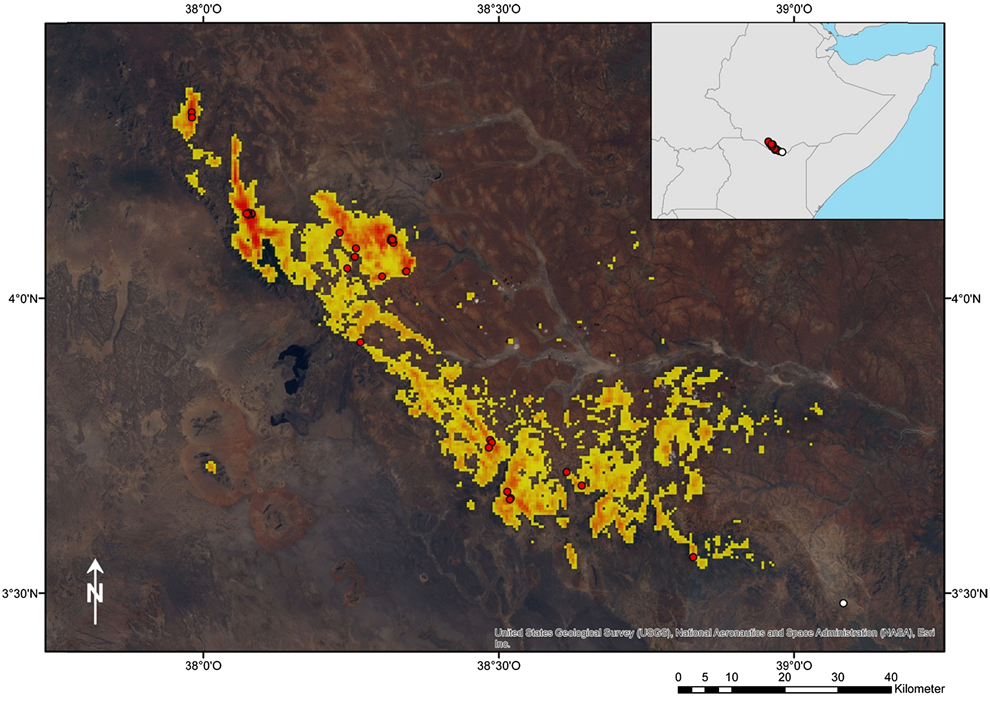
Figure 2. Projected distribution of the Black-fronted Francolin in South Ethiopia and North Kenya based on species distribution modelling. Actual records of occurrence (circles) are indicated. Warmer colours indicate high probabilities of occurrences, whereas cooler colours indicate lower probabilities (logistic output format of Maxent). The species appears closely bound to a mountain range running from Dilo over Mega in Ethiopia until the Moyale area in Kenya. Its occurrence is apparently defined by the special ecological conditions of this mountain chain in terms of being cooler, moister and less densely vegetated than the surrounding savannah habitats.
The modelled range (Figure 2) covers an area of 1,286 km2 (“extent of occurrence”, EOO, following IUCN 2001). However, in many parts of this range we did not find any signs of Black-fronted Francolins during fieldwork despite our focussed search. This applies particularly to the areas between the mountain ridges in the North and to the fragmented areas in the south-east. These areas are currently widely used for agriculture and/ or are spatially separated from areas with confirmed populations. Therefore, we estimate that only about one third of the modelled range is actually populated by the Black-fronted Francolin. This area, around 385 km2, corresponds to the “area of occupancy” (AOO) according to the IUCN Red List criteria (IUCN 2001).
When modelling the range, we received excellent AUC scores (AUCTraining = 0.99, AUCTest = 0.98) across the 100 Maxent replicates. Annual mean temperature (Figure 3A) had the highest variable contribution (45.9%), followed by precipitation of warmest quarter (Figure 3H; 15.8%), annual precipitation (Figure 3E; 15.6%), brightness (Figure 3I; 8.2%) and greenness (Figure 3J; 5.3%). The remaining variables contributed less than 5% to the final model (Figure 3). This indicates the Black-fronted Francolin’s specific preference for relatively cool (max. 16°C annual mean temperature) and moist conditions (> 700 mm annual precipitation and > 500 mm precipitation during warmest quarter, respectively) in relatively open and less densely vegetated habitats.
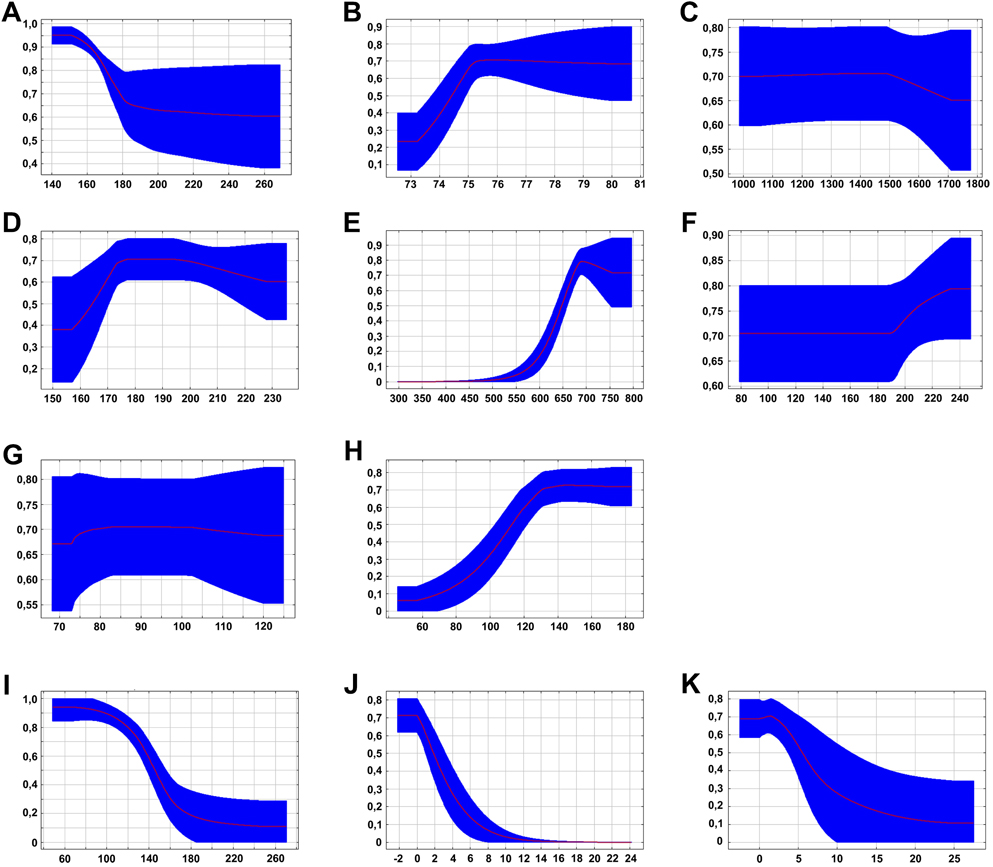
Figure 3. Parameters of the SDM model with values on Y-axes representing the respective probabilities of presence. A – annual mean temperature, B – Isothermality, C – Temperature seasonality, D – Annual temperature range, E – Annual precipitation, F – Precipitation of wettest month, G – Precipitation seasonality, H – precipitation of warmest quarter, I – Brightness, J – Greenness, K – Wetness. The highest contribution of variables had annual mean temperature (A; 45.9%), followed by precipitation of warmest quarter (H; 15.8%), annual precipitation (E; 15.6%), brightness (I; 8.2%) and greenness (J; 5.3%). The remaining variables contributed less than 5% to the final model.
Population
In 2014, we collected GPS coordinates for 21 localities with evidence for the occurrence of the Black-fronted Francolin (Figure 1): visual and acoustic evidence (n = 4), droppings (n = 7), feathers (n = 1), credible information by local villagers (n = 9). No call activity was recorded by the automatic sound recording devices in June 2014, probably due to the absence of usual seasonal rainfall in April and May and associated drought. As a result, we estimated the population in two areas of approximately 36 km2 (i.e., extension zones 2 and 4 in Figure 4), to comprise between 100 and 200 adult individuals. Extrapolating these numbers to the whole area of occupancy (385 km2) the total population size ranges between 1,100 and 2,100 mature individuals.
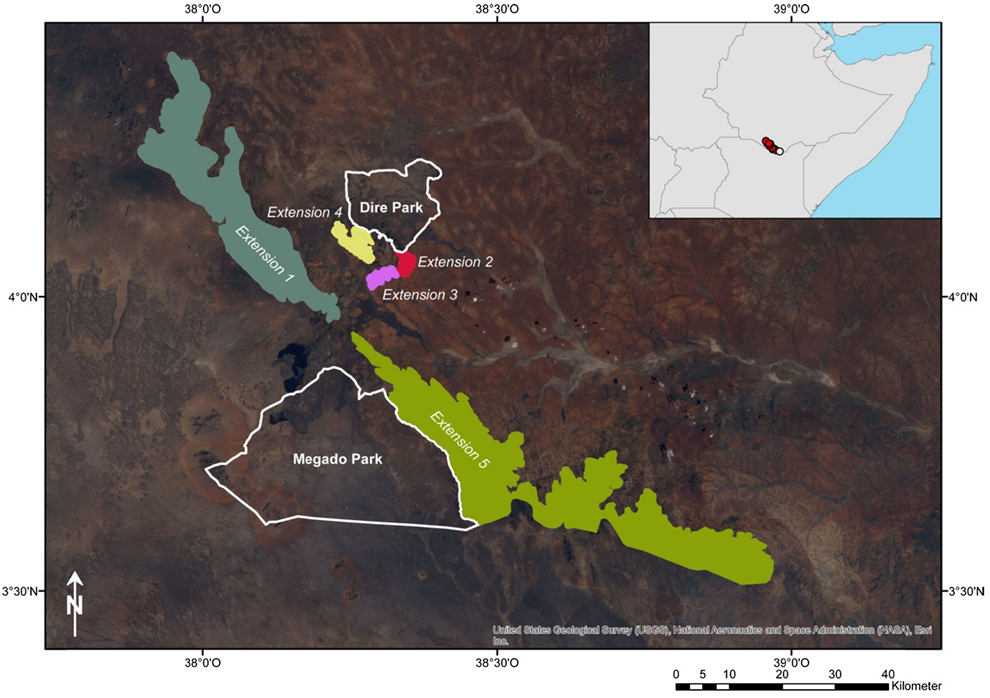
Figure 4. Proposal for extensions of the currently projected Borana National Park. White lines indicate the projected Dire and Megado Park which clearly lie outside of the occurrence of the Black-fronted Francolin. Therefore, five extensions of protected areas are suggested.
Habitat and ecology
Based on our field data, the presence of the Black-fronted Francolin is limited to a mountainous, richly structured terrain. Its altitudinal distribution ranges from 1,096 m asl in the south to 2,223 m in the north. Important habitat features are rocks, shrubby vegetation and a herbaceous, semi-open ground cover. Whenever we observed birds directly or where we found droppings, these were on or in the immediate vicinity of smaller or medium-sized rock structures, surrounded by shrubs with a semi-open herbal undergrowth. Moreover, males often called from exposed sites on the top of the rocks. In contrast, no birds were found on rocky, dry slopes with rich grassy vegetation and in agricultural areas.
Discussion
Assessment of IUCN Red List Category & Criteria
Our data indicate that the assessment of the Black-fronted Francolin as ‘Least Concern’ (EOO < 20,000 km2, stable population, > 10,000 mature individuals) by BirdLife International (2016) does not reflect the actual situation. Instead, following the IUCN Red List criteria version 3.1 (IUCN 2001), we conclude that the species should be classified as ‘Endangered’ with a declining population trend on criteria A2c; B1ab(i,iii,v); B2a,b(i,iii,v). This evaluation is justified below.
Geographic range
The EOO (criterion B1 in IUCN 2001) is around 1,300 km2 and thus well below the maximum range size of 5,000 km2 of the ‘Endangered’ category. The following subcriteria are relevant:
a, because the population is severely fragmented. We have found the Black-fronted Francolin only at a few places within the modelled range (Figure 2). Thus, we conclude on four largely separated sub-populations: two in the high mountains in the far north, one around Mega town and one along the escarpment and its craggy hinterland of its central and southern part. In between, there is either farmland or dry scrubland, both of which are unsuitable habitats.
b(ii), because the AOO is in decline. Considerable parts of the potential range are used agriculturally today and therefore uninhabited by the Black-fronted Francolin. This applies in particular to the area around Mega town, but also to the south-eastern part of the range (pers. obs.). By contrast, about 50 years ago, farming was virtually unknown to the local Borana people.
b(iii), because the area and/or quality of habitat is in decline. Many of the remaining francolin habitats are under heavy exploitation pressure because of overgrazing, woodcutting and slash-and-burn practices. This pressure is increasing constantly due to the growing population in the surrounding settlements, especially Mega, but also Dilo and Megado (cf. Bassi Reference Bassi2010, Worldometer 2017). The process is further accelerated by the recent expansion of asphalt roads (between Yabello and Mega) and the construction of new roads (e.g. between Dubluk and Dilo).
b(v), because the number of mature individuals is in decline. As a consequence of the described loss of range and habitats, a significant decrease in population size is to be assumed. Hunting birds with snares is an additional factor affecting the population (Töpfer et al. Reference Töpfer, Podsiadlowski and Gedeon2014).
Likewise, the AOO (criterion B2 in IUCN 2001) is estimated at 385 km2 and thus below the maximum range size of 500 km2 of the ‘Endangered’ category. The following subcriteria are relevant:
a, because the population is severely fragmented,
b(i), because the EOO is in decline,
b(iii), because the area and/or quality of habitat is in decline, and
b(v), because the number of mature individuals is in decline.
Population
We suspect a past reduction of population of ≥ 50%, based on the decline in AOO and habitat quality with irreversible causes of reduction (details in threats paragraph). This corresponds to the requirements of criterion A2c in IUCN (2001). We estimate the total population size at about 1,100–2,100 mature individuals. This is below the maximum threshold of 2,500 mature individuals of the ‘Endangered’ category under criterion C (IUCN (2001). However, we cannot assess the specific conditions of the mandatory criteria C1 and C2 precisely since there are no detailed data on (declining) population trends available to date.
Threats and conservation actions
Major threats have already been identified by Töpfer et al. (Reference Töpfer, Podsiadlowski and Gedeon2014), among them range and habitat losses due to agricultural expansion, grazing pressure, commercial firewood and timber exploitation, and hunting. As stated above, these factors will persist and, in the context of human population growth, even increase in extent.
Possible climatic changes have to be considered as well. Projections indicate that an increase of maximum annual temperatures is to be expected in south-eastern Ethiopia (period 2046-2065 compared to historical data; Ethiopian Panel on Climate Change 2015). Projected changes in precipitation may be even more severe: wetter scenarios for the same period indicate earlier spring rains and longer autumn rains while drier scenarios indicate a drastic reduction of spring rains.
The latter would seriously impact the reproductive success of the Black-fronted Francolin in particular (Töpfer et al. Reference Töpfer, Podsiadlowski and Gedeon2014). As the birds prefer a relatively cool and moist environment, the aforementioned climatic forecasts, along with anthropogenic habitat exploitation pressure, render the survival of the species doubtful. Thus, we suggest that the following most important conservation actions are implemented as soon as possible to save the remaining population of the Black-fronted Francolin:
Extend the currently projected Borana National Park to include the main range of the species. A corresponding proposal is shown in Figure 4. The extensions include the core areas of the range according to the modelling results, with human settlements and agricultural areas being omitted. This procedure results in three small extensions around the town of Mega and two major extensions along the mountain ranges in the west and south, comprising a total area of 1,212 km2.
Enforce the sustainable use of natural resources, especially firewood, timber and grazing, in areas of importance to the Black-fronted Francolin.
Inform villagers about threats to and the need for protection of the Black-fronted Francolin.
Restrict the spread of agriculture into mountain forests.
Prevent forest fires and prohibit slash-and-burn practices.
Ban bird hunting with snares.
Undertake further work on the Black-fronted Francolin to assess range and population changes in order to better understand its ecology and threats in response to habitat and climate change.
Acknowledgements
We thank the World Pheasant Association (WPA) and the Zoologische Gesellschaft für Arten- und Populationsschutz (ZGAP) for their generosity in supporting our research in southern Ethiopia in 2014. Our studies in 2013 were supported by a grant from the Gesellschaft für Tropenornithologie (GTO). We are grateful to many people for their help in the field, including Tesfaye Mekonnen, Okotu Dida and Ali Guche. We thank Stefanie Rick (ZFMK) for her helpful comments on the text. Heiner Nagel (Landesamt für Umweltschutz Sachsen-Anhalt) supported GIS data manipulation. We also thank the Ethiopian Wildlife Conservation Authority and the Oromia Forest and Wildlife Enterprise for their institutional support.




