


Introduction
Endemic island birds typically exist in small populations within restricted and specialised habitat (Simberloff Reference Simberloff1995) where they are 40 times more likely to go extinct than continental species (Johnson and Stattersfield Reference Johnson and Stattersfield1990). Birds that are highly adapted to restricted, elevated forest habitats on islands are under a particularly high risk of extinction when their habitat is fragmented via land clearance and/or invaded by introduced species (see Savidge Reference Savidge1987, Johnson and Stattersfield Reference Johnson and Stattersfield1990, Steadman Reference Steadman1995). In Hawaii, the cooler high-elevation areas provide refuge for rare endemic honeycreepers (Benning et al. Reference Benning, Lapointe, Atkinson and Vitousek2002). In contrast, elevated habitats in the Galápagos Islands have high prevalence and infestations of introduced fly larvae parasites Philornis downsi (see Dudaniec et al. Reference Dudaniec, Fessl and Kleindorfer2007, Wiedenfeld et al. Reference Wiedenfeld, Jimènez, Fessl, Kleindorfer and Valerezo2007), and these highland areas have been extensively cleared for agriculture, leaving only fragmented and invaded habitats for mid-to-high elevation birds.
The P. downsi invasion is one of the most recent, yet most significant threats to all landbirds in the Galápagos Islands (Fessl and Tebbich Reference Fessl and Tebbich2002, Causton et al. Reference Causton, Peck, Sinclair, Roque-Albelo, Hodgson and Landry2006), including the iconic group of Darwin’s finches. Adult P. downsi flies lay eggs in bird’s nests that hatch into larvae and proceed to feed on the blood and body tissues of developing nestlings by external attachment or by entering through the nasal cavity to feed internally (Fessl et al. Reference Fessl, Sinclair and Kleindorfer2006b, O’Connor et al. in press). Impacts of P. downsi parasitism in Darwin’s finch nestlings include up to 55% blood loss, multiple body wounds and infections, increased mortality (Fessl et al. Reference Fessl, Sinclair and Kleindorfer2006b), reduced fledging success (shown experimentally in Fessl et al. Reference Fessl, Kleindorfer and Tebbich2006a), and beak deformation of fledglings that survive into adulthood (Galligan and Kleindorfer Reference Galligan and Kleindorfer2009). The parasite is more prevalent on the three elevated islands with human settlements (Wiedenfeld et al. Reference Wiedenfeld, Jimènez, Fessl, Kleindorfer and Valerezo2007), such as Floreana Island (which was visited in 1835 by Charles Darwin). With noticeable human impacts even in Darwin’s time, Floreana Island now has the longest history of human settlement and avian extinctions in the entire Galápagos archipelago. Native habitat on Floreana Island has been disturbed via agricultural clearance (Lack Reference Lack1947, Sulloway Reference Sulloway1982, Steadman Reference Steadman1986), and invaded by introduced plants (Mauchamp Reference Mauchamp1997, BirdLife International 2009a), predators (Curry Reference Curry1986, Baskin Reference Baskin2002, Grant et al. Reference Grant, Grant, Petren and Keller2005), and avian parasites and infectious diseases (Fessl and Tebbich Reference Fessl and Tebbich2002, Dudaniec et al. Reference Dudaniec, Kleindorfer and Fessl2006, Kleindorfer and Dudaniec Reference Kleindorfer, Chapman, Winkler and Sulloway2006, Sharon Deem pers. comm.). To date, three Darwin’s finch species have become extinct on Floreana Island, including both the Large Ground Finch Geospiza magnirostris and Sharp-beaked Finch G. nebulosa by about 1870 (reviewed in Sulloway Reference Sulloway1982, Steadman Reference Steadman1986) and the reported loss of the Warbler Finch Certhidia fusca by 2004 (see Grant et al. Reference Grant, Grant, Petren and Keller2005). Once common, the Floreana Mockingbird Nesomimus trifasciatus disappeared from Floreana by 1895 and only survives today on two small islets: Champion and Gardener-by-Floreana (Curry Reference Curry1986). Floreana Island also supports the only population of the Medium Tree Finch (Lack Reference Lack1947, Grant Reference Grant1999), which is restricted to fragmented forest patches in the humid highland area. A recent study has found that, across years, Medium Tree Finch nests have one of the highest P. downsi intensity documented in any Darwin finch species, and that P. downsi parasitism was responsible for mortality in 41% of nestlings (O’Connor et al. Reference O’Connor, Sulloway, Robertson and Kleindorfer2010).
The preferred nesting tree of the Medium Tree Finch, Scalesia pedunculata, is endangered (Boada Reference Boada2005) and only remains in small patches within the Floreana highlands. The size of the Medium Tree Finch population is unknown, yet evidence suggests it may be declining due to lack of suitable habitat and high nestling mortality. Adult age structure within the population is also unknown, yet is an essential component for predicting adult survival. Another closely related species, the Large Tree Finch C. psittacula, is also suspected to be declining on Floreana Island (Grant et al. Reference Grant, Grant, Petren and Keller2005).
In this study we conducted surveys of all birds in the highlands across four sites, three of which contain the largest remnant patches of Scalesia forest, to estimate the maximum population size of several highland bird species. Our objectives were to (1) survey densities of bird species in the highlands of Floreana Island across years, with special reference to Darwin’s finches, flycatchers, and yellow warblers; and (2) assess male age structure of nesting Medium Tree Finches across years.
Methods
Study site: Cerro Pajas Volcano (2004 and 2008)
We surveyed bird abundance at the base of the Cerro Pajas Volcano (c. 300–400m), which is the highest volcano (maximum elevation 550 m) on Floreana Island (173 km2, 1°28′S, 90°48′W; Figure 1). The survey was conducted across seven days in mid-February of 2004 and 2008. The Cerro Pajas study site covers an area of approximately 2.4 km2 and is characterised by humid forest habitat, dominated by the tallest endemic Scalesia pedunculata (up to 15 m high) found on the island (Table 1).
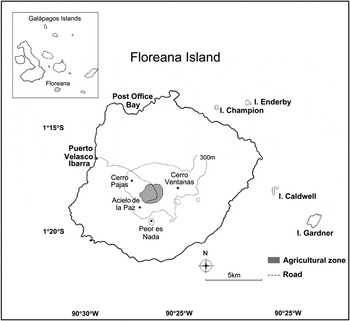
Figure 1. Map of Floreana Island, Galápagos Archipelago, Ecuador. The highlands zone includes all areas within the 300 m contour line. Site locations and contour lines were established using GPS coordinates and Google Earth ProTM.
Table 1. Description of vegetation found at each of the four survey sites. The total area with native forest is regarded as any forested area dominated by native tree species. We have noted the occurrence of seven of the most common dominant highland plant species across sites in the following descending order of abundance: (1) Dominant, (2) Common, (3) Patch (common, but only within a specified area in km2), (4) Present, (5) Absent. Site sizes were calculated using Google Earth ProTM.
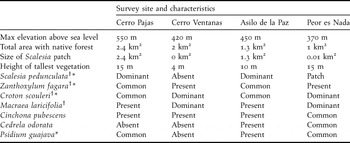
† Native species
* Used as nesting substrate by tree finches (see results)
Study site: Other highland forest areas (2008 only)
To gain a better understanding of the entire highland forest bird community, we surveyed three additional highland sites in 2008 (Figure 1). These sites were situated around the base of mountains and volcanoes and are considered to be the last patches of native highland forest containing Scalesia (Walter Cruz, Walter Simbaña pers. comm.). The characteristics of each site, including total patch size (km2), Scalesia patch size (km2), and dominant plant species are shown in Table 1. Notably, Peor es Nada supports only a small 100 m × 100 m patch of Scalesia on its north-western slope. Cerro Ventanas does not contain Scalesia, but it does contain native highland shrubs (not trees). Our study has essentially surveyed finch populations in prime habitat in the peak breeding season when song and foraging activity is very high. Birds – especially tree finches – do not generally nest in the agricultural area due to lack of suitable nesting substrate, and would only be temporary visitors there to forage on crops.
Survey methods
We followed protocols of the variable circular plot method to obtain our survey data (see Martin et al. Reference Martin, Paine, Conway, Hochachka, Allen and Jenkins1997). At Cerro Pajas we sampled from 15 point counts along the trail leading to the inner crater. Fourteen point counts were made at Cerro Ventanas, 11 point counts at Asilo de la Paz, and 12 at Peor es Nada. All point counts were conducted a minimum of 150 m apart. At each survey point we recorded: (1) GPS co-ordinates, (2) species identity, (3) estimated radial distance of each bird from the observer (in 10-m intervals), and (4) detection method (sight or sound). Birds were recorded up to a distance of 200 m away, but we only analysed data from within a 20–70 m radius due to a decline in rates of detection, which varied by species. Surveys were conducted for five minutes at each point, which was sufficient time to identify all birds actively using the area (this was trialled in a pilot study on Santa Cruz Island in 2000 and repeated in 2008). At each point, the observer surveyed the area facing 0°, 90°, 180° and 270° to eliminate visual or audio bias while facing only one direction. Counts were taken during peak bird activity which was between 06h00 and 12h00.
Analysis of population density
Population density estimates (individual birds km−2) and species detectability estimates were initially tested using the software program DISTANCE version 5.0 (Thomas et al. Reference Thomas, Laake, Strindberg, Marques, Buckland, Borchers, Anderson, Burnham, Hedley, Pollard, Bishop and Marques2006), but our data did not meet the model assumptions/criteria. We also did not obtain the minimum 60–100 detections per species recommended for calculating accurate density estimates with DISTANCE (Somershoe et al. Reference Somershoe, Twedt and Reid2006) because of restrictions imposed by habitat patch sizes/accessibility.
We instead calculated population density using the inflection-point-per-species method (described in Reynolds et al. Reference Reynolds, Scott and Nussbaum1980). We plotted the number of detections of each species within 10-m concentric bands and determined the distance from the observer at which its rate of detection begins to decline (the inflection point). The detectability of each species is affected by: (1) the ability of its song to be heard through thick vegetation, and (2) its visibility to the observer (due to differences in foraging behaviour). Habitat density analyses only include birds observed at a distance within the inflection point. For the tree finches and ground finches this point was 70 m, but it differed for the remaining species (see Table 2). The number of birds km−2 was determined by calculating the number of birds from each species within each 10-m concentric circle, dividing the number by the total area within each circle, and dividing the density estimate by the number of survey points.
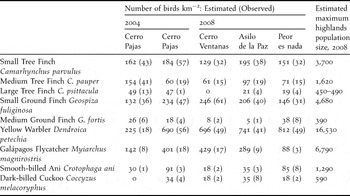
Table 2. Population density estimates for bird species surveyed at Floreana Island highland forest sites in 2004 and 2008. Methods for calculating density are detailed in the methods section. The maximum population size for 2008 was calculated using the mean density (birds km−2) across the four sites divided by the total area of native forest (22.5 km2). For the Large Tree Finch, a range in population size was calculated (see results, Cerro Ventanas was excluded from the lower estimate). The inflection point (distance from the observer at which the density of each species declined) was 70 m for each of the five species of Darwin’s finches, 50 m for the Smooth-billed Ani and Dark-billed Cuckoo, 40 m for the Yellow Warbler, and 30 m for the Galápagos Flycatcher.
Male age structure in Medium Tree Finches
We are particularly interested in estimating changes in population density in the Medium Tree Finch, because (1) this species is locally restricted, (2) has high P. downsi parasite intensity (O’Connor et al. Reference O’Connor, Sulloway, Robertson and Kleindorfer2010), and (3) only six of the 63 monitored nests produced fledglings (O’Connor et al. Reference O’Connor, Sulloway, Robertson and Kleindorfer2010). We examined the age structure of Medium Tree Finches nesting in Scalesia forest at the base of Cerro Pajas during the peak finch-breeding period (February-April) of 2006 and 2008. Male tree finches increase the proportion of black on their chins and crowns with each year of annual moult until attaining a fully black head by five years of age (Lack Reference Lack1947, Grant Reference Grant1999, Kleindorfer Reference Kleindorfer2007). Therefore, male age is considered to co-vary with male plumage colouration. Each male found singing at a display nest was assigned a unique colour category based on the length of black on the chin (cm) and the extent of black on the crown (see Kleindorfer Reference Kleindorfer2007). Black 0 (zero) males are yearling males and Black 5 males are five years and older. Females of all species of Darwin’s finches remain brown throughout their lives; therefore female plumage colouration gives no indication of age.
Our analysis of male age structure was restricted to males that sang at nests to attract females. We located nests by systematically searching four 100 m × 200 m study plots at elevations of 300–400 m within the Cerro Pajas site and locating either (1) singing males with display nests, (2) males building new nests or (3) active nests with a female present. We found 27 Medium Tree Finch males with display nests in 2006, and 36 in 2008. We noted the nesting tree and height for each nest, and GPS co-ordinates were recorded with a hand held Garmin GCX12. Nesting activity was monitored for every unpaired singing male using 20 minute continuous focal sampling at least every second day to determine the status of the nest. At this time, we recorded male colour category.
Results
Avian population trends in 2004 and 2008 at Cerro Pajas
A total of 344 individuals of nine bird species was counted between years at the Cerro Pajas site within 70 m point counts and was included in the population density analysis (Table 2). The population density (number of birds km−2) of some species differed across years, while it remained stable in others (Table 2). Notably, we found that the Medium Tree Finch population declined from 154 birds km−2 in 2004 to 60 birds km−2 in 2008 at the Cerro Pajas site, a reduction of 61%. In 2008, we observed substantially higher densities of six of the other eight species, and collectively these eight species increased their numbers by an average of 280%.
Avian population size across four highland sites in 2008
In 2008, the density of most species was similar across sites (Table 2) with the exception of Large Tree Finches, which were not seen or heard at Cerro Ventanas. Avian species composition was most different at Peor es Nada where we observed (1) fewer Small Ground Finches and Galápagos Flycatchers Myiarchus magnirostris, and (2) more Medium Ground Finches and Yellow Warblers Dendroica petechia compared with other sites.
Calculating population estimates for the Medium and Large Tree Finch according to habitat suitability
The highland area of Floreana covers an area of approximately 25 km2. It contains an inner region of approximately 2.5 km2 that has been cleared for agriculture. The Floreana highlands thus contain a 22.5 km2 uncleared area that may contain suitable forest habitat for Medium Tree Finches. Scalesia dominated forest is estimated at 3.71 km2 (see Table 1).
A mean of 72 Medium Tree Finches km−2 was observed in surveyed forest habitat (calculated as an average over the four survey sites in 2008). Using 22.5 km2 as the maximum habitat size, we estimate that the entire Medium Tree Finch population on Floreana Island consists of up to 1,620 individuals (Table 2). Using the same reasoning, the Large Tree Finch population has a mean of 22 finches km−2 with up to 490 individuals on Floreana Island in 2008. However, Large Tree Finches were not detected at Cerro Ventanas. If we exclude this area from our calculation, then the maximum number of Large Tree Finches on Floreana Island in 2008 was 450 (Table 2).
Male age structure in Medium Tree Finches
We recorded data from 63 males that were observed singing at 77 display nests to attract a female. Some males built multiple display nests. Most nests (83%) were built in S. pedunculata (n = 63), 14% in Z. fagara (n = 11), 2% in C. scouleri (n = 2), and 1% in guava (n = 1). Mean nest height was 6.15 m (± SE 0.22), though nest height ranged from 3 to 12 m high. Most nesting males were young (88% of nesting males were Black category 0-3). Only 12% of males were in category Black 4, and no Black 5 males were seen or mist-netted. There was a significant difference in the age structure of nesting males between 2006 and 2008: no Black 0 males were found in 2006 and more Black 1 males were found in 2008 (Likelihood ratio = 21.79, df = 4, P < 0.001) (Figure 2). This finding suggests that breeding did not occur in 2005, which was a drought year in the archipelago.
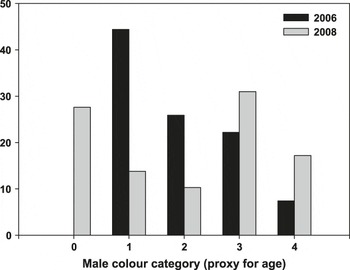
Figure 2. Change in the percentage of male colour categories at active Medium Tree Finch nests in the Floreana highlands between 2006 and 2008. Note that there were no Black 0 (young) males in 2006, which suggests unsuccessful breeding in the previous year(s). No Black 5 (old) males were observed in either study year.
Discussion
Population trends in tree finches
This is the first study to estimate the population size/density and range restrictions of any highland bird community in the Galápagos Islands. Of the three Camarhynchus tree finch species observed in the Floreana survey, Small Tree Finches were the most common (Table 2). Medium and Large Tree Finches were observed at lower densities relative to Small Tree Finches using both point count sampling and non-targeted mist netting (Christensen and Kleindorfer unpubl. data). Large Tree Finches were always the least common tree finch species in both the point count and mist netting surveys. This species warrants close monitoring on Floreana Island because it exhibits three hallmark features of a species that could be on a trajectory to extinction: (1) small population size: < 450 individuals, (2) large body size (which is associated with higher P. downsi parasitism levels), and (3) more specialist foraging behaviour (Bennett and Owens Reference Bennett and Owens1997, Reference Bennett and Owens2002; discussed in Christensen and Kleindorfer Reference Christensen and Kleindorfer2008). In addition, we know that only one Large Tree Finch nest that we monitored over two years produced fledglings, and this nest had high parasite intensity (50 P. downsi larvae in the nest; O’Connor et al. Reference O’Connor, Sulloway, Robertson and Kleindorfer2010).
Although the Small and Large Tree Finch populations appear to be stable, the intermediate-sized form of these two species – the Medium Tree Finch – has declined in numbers. By 2008, the Medium Tree Finch population at Cerro Pajas had dropped to 39% of its size in 2004, which is equivalent to a decline of ~ 15% per year. Natural populations of birds are known to fluctuate in numbers across years (Holmes and Sherry Reference Holmes and Sherry2001), and declining populations can recover from major losses (Roth and Johnson Reference Roth and Johnson1993, Holmes and Sherry Reference Holmes and Sherry2001, Hale and Briskie Reference Hale and Briskie2009). However, in the case of the Medium Tree Finch, the probable causes for its decline (introduced parasites and predators, and habitat loss) are not likely to subside without concentrated effort. Fledging success was lower in 2008 (a wet year) compared with 2006 (a dry year) despite abundant resources, high bird nesting activity, and large clutch size. Lower fledging success in 2008 compared with 2006 was explained by higher levels of nestling depredation and parasitism (O’Connor et al. Reference O’Connor, Sulloway, Robertson and Kleindorfer2010). The entire Medium Tree Finch population currently consists of a maximum 1,620 individuals, which is significantly smaller than it was 50–100 years ago (Kleindorfer and Sulloway in prep.). This finding may be explained by a combination of low nesting success and low adult survival. In a recent comparison of P. downsi parasite intensity in three Floreana Island tree finch species, O’Connor et al. (Reference O’Connor, Sulloway, Robertson and Kleindorfer2010) found that the larger bodied Medium and Large Tree Finches had higher parasite intensity than the Small Tree Finch. The study also found that Medium Tree Finches had higher P. downsi parasite intensity than expected based on body mass and that parasitism was responsible for 41% of nestling mortality. The Medium Tree Finch has since been uplisted from ‘Vulnerable’ to ‘Critically Endangered’ on the 2009 IUCN Red List (Birdlife International 2009b).
Tree finch habitat on Floreana Island
We estimated maximum population sizes of each species according to their densities km−2 across sites in 2008, multiplied by the area of native forest area. However not all of the maximum uncleared highland forest area used to calculate population range/size may be suitable tree finch habitat, as non-Scalesia forest is of considerably lower height (eg., Cerro Ventanas, maximum 4 m) (Table 1) and lower vegetation density. For example, Large Tree Finches were only observed at sites with tall trees, and where Scalesia is present (Table 2). Medium Tree Finches nested at a mean height of 6.15 m, and 83% of their nests were in Scalesia trees (at Cerro Pajas). Displacement of native Scalesia forest by exotic fruit trees was identified as a conservation problem on Floreana as early as 1957 (Eibl-Eibesfeldt Reference Eibl-Eibesfeldt1959), and the species now remains only in small patches because of agricultural clearance, competition from invasive plants, and destruction by introduced mammals.
Male age structure in Medium Tree Finches
Of the 63 Medium Tree Finch males observed singing at nests to attract females, most were less than four years old (see Figure 2), as indicated by the extent of black colouration in crown and chin plumage. Recent studies have found that older male Small Tree Finches have higher pairing success compared with young males of the same species (Kleindorfer Reference Kleindorfer2007, Kleindorfer et al. Reference Kleindorfer, Sulloway and O’Connor2009). Kleindorfer (Reference Kleindorfer2007) also found that older males built more concealed nests that were less likely to be depredated and experienced higher fledging success. Thus, the scarcity of older Medium Tree Finch males in the population may negatively influence nesting outcome if male age predicts nesting success.
Our finding of no young (Black 0) males in 2006 suggests unsuccessful breeding in the dry year of 2005 (see Dudaniec et al. Reference Dudaniec, Fessl and Kleindorfer2007 for rainfall data). Many two-year-old (Black 1) males were found in 2008 and may represent a cohort that fledged in 2006. Few Black 4 and no Black 5 males were observed, which suggests that adults are not surviving past 4–5 years of age. The overall young age structure of the Medium Tree Finch population signifies that adult males are not generally surviving to full maturity (five years or more).
Population trends in other bird species
Significant changes in Galápagos finch population sizes have been reported from small and low-elevation islands such as Daphne Major where selection pressures are high and extremely variable across years (Grant Reference Grant1999). The six elevated Galápagos islands with forest highland regions receive higher and more consistent annual rainfall (Wiedenfeld et al. Reference Wiedenfeld, Jimènez, Fessl, Kleindorfer and Valerezo2007), which should support more stable bird population sizes. Here, we show that some bird species experienced rapid and dramatic population fluctuations in as little as four years on the elevated island of Floreana. Between 2004 and 2008 at the Cerro Pajas site, the Medium Tree Finch declined in numbers and the density of all other species either increased or remained relatively stable (Table 2). The ratio of change in densities of all other species in Table 2, compared to the Medium Tree Finch, is 7.2 to 1. Thus the Medium Tree Finch population, as of 2008, represented about 14% of what would be expected based on the more favourable ecological conditions that year, as reflected in the population sizes of other species. Warblers and flycatchers are insectivorous and may have been more commonly observed in 2008 because of an increase in insect abundance due to heavier rainfall. Small Ground Finches have the widest foraging breadth among Darwin’s finches, which enables them to exploit modified habitats from which more specialised finches could be excluded (see also Kleindorfer et al. Reference Kleindorfer, Chapman, Winkler and Sulloway2006, Kleindorfer and Mitchell Reference Kleindorfer, Mitchell, Kleindorfer and Wind2009, Sulloway and Kleindorfer in prep. for a discussion of habitat use and range expansion on Santa Cruz Island). The increase in Small Ground Finch abundance may be due to their expansion into the highlands during a period of drought, where they can use the available food resources given their generalist foraging behaviour and diet.
The Vegetarian Finch Platyspiza crassirostris was not detected in surveys, though one female was observed at the Cerro Pajas site by J.O’C in 2008. The Vermilion Flycatcher Pyrocephalus rubinus, once considered relatively common in the highlands of Floreana Island (Edwin Egas and Walter Cruz pers. comm.), was not detected in our bird surveys, although two Vermilion Flycatchers were observed by S.K. in 2004 at the Cerro Pajas site while conducting other fieldwork. Finally, although the Warbler Finch Certhidea fusca was considered locally extinct by 2004 (Grant et al. Reference Grant, Grant, Petren and Keller2005), we heard a male singing in 2008 (approximately 20 m high in a Cedrela odorata tree at Asilo de la Paz).
Conclusion
Here we show that the sole population of Darwin’s Medium Tree Finch is small, declining, and at risk of extinction. Three other species (Vermilion Flycatcher, Vegetarian Finch, and Warbler Finch), once common in the Floreana highlands, have also become extremely rare. The Galápagos National Park has recently implemented programmes to control and eradicate invasive plants and feral goats within the Floreana highlands. To help prevent another local avian extinction on the island, there is a need for effective P. downsi parasite and rodent predator control, as well as regeneration and expansion of the endemic Scalesia forest.
Acknowledgements
This paper is contribution number 2005 of the Charles Darwin Foundation for the Galápagos Islands. We are grateful to the Charles Darwin Research Station and Galápagos National Park Service for the opportunity to work on the Galápagos, and for logistical support. This work was generously supported by Flinders University (Research Establishment Grant), Conservation International, the American Bird Conservancy, the Winifred Violet Scott Trust with awards to SK, and also Flinders University (travel grant), the Royal Zoological Society of South Australia, Birdfair/RSPB Research Fund for Endangered Birds and the Australian Federation of University Women (SA) with awards to JO’C. TAME airlines provided reduced airfares. We thank Santos Humberto for field assistance, Walter Cruz and Michael Dvorak for advising JO’C on fieldwork methodology, and Chris Holden for assistance with DISTANCE software.




