





1. Introduction
Bilinguals acquire and use their second language without apparent difficulties. This belies the fact that bilingualism is a major processing demand for the cognitive system. Evidence shows that the use of multiple languages leads to parallel activation and competition between them, requiring the speaker to prioritise one and inhibit the non-target language(s) (Green, Reference Green1998). These additional processing demands have been shown to lead to neuroplastic adaptation in brain structure and function across the lifespan (Burgaleta et al., Reference Burgaleta, Sanjuán, Ventura-Campos, Sebastian-Galles and Ávila2016; Hayakawa & Marian, Reference Hayakawa and Marian2019; Kousaie & Phillips, Reference Kousaie and Phillips2012).
One particular domain where bilingualism has been shown to exert significant influence is the domain of selective attention (Abutalebi et al., Reference Abutalebi, Della Rosa, Green, Hernandez, Scifo, Keim, Cappa and Costa2012; Bialystok, Reference Bialystok2015). Yet, the exact mechanism of selective attention modulation in bilinguals remains a matter of debate. One recently proposed view is that, instead of enhanced attentional capacity (or ‘executive attention’, cf. Bialystok, Reference Bialystok2017), adaptation to the demands of bilingualism reflects the process of redistributing the available resources to support optimal behavioural performance under the increased processing load (Olguin et al., Reference Olguin, Cekic, Bekinschtein, Katsos and Bozic2019; Phelps et al., Reference Phelps, Attaheri and Bozic2022). This account is rooted in the general concept of functional plasticity and degeneracy (Mason et al., Reference Mason, Domínguez D, Winter and Grignolio2015; Navarro-Torres et al., Reference Navarro-Torres, Beatty-Martínez, Kroll and Green2021), a common feature in biological systems that enables their flexible adaptation to changing environments. Such flexible adaptation allows the biological systems to perform comparable function, or achieve equivalent performance, supported by different underlying configurations (Edelman & Gally, Reference Edelman and Gally2001; Whitacre & Bender, Reference Whitacre and Bender2010). This view of attentional modulation in bilingualism is consistent with reports that neural differences between monolinguals and bilinguals have been observed even when they display equivalent behavioural performance (Bialystok et al., Reference Bialystok, Craik, Grady, Chau, Ishii, Gunji and Pantev2005; Kousaie & Phillips, Reference Kousaie and Phillips2012; Olguin et al., Reference Olguin, Cekic, Bekinschtein, Katsos and Bozic2019).
The current set of studies investigates further the notion of flexible functional adaptation of selective attention in bilingualism. We test how the available attentional capacity is distributed to support behavioural performance when the system is presented with a processing load extending beyond the typical demands, and how this flexible adaptation changes with development and maturation.
1.1. Selective attention in bilingualism
There is substantial evidence that the demands of learning and using multiple languages modulate the neurocognitive architecture of selective attention. For example, bilinguals have been shown to have stronger subcortical representation of fundamental frequency (F0) and more consistent neural responses to attended syllables compared to monolinguals (Krizman et al., Reference Krizman, Marian, Shook, Skoe and Kraus2012, Reference Krizman, Skoe, Marian and Kraus2014), even when they have been exposed to different combinations of languages (Skoe et al., Reference Skoe, Burakiewicz, Figueiredo and Hardin2017).
There is conflicting evidence, however, on the behavioural consequences of these neural adaptations. One widely held view is that the increased processing demands of managing multiple languages lead to enhanced capacity for selective attention, resulting in better performance for bilinguals on selective attention tasks (Bialystok, Reference Bialystok2017). This hypothesis has been supported by numerous behavioural studies across age groups and tasks (Bialystok et al., Reference Bialystok, Craik and Luk2012; Costa et al., Reference Costa, Hernández and Sebastián-Gallés2008; Kovács & Mehler, Reference Kovács and Mehler2009; Poulin-Dubois et al., Reference Poulin-Dubois, Blaye, Coutya and Bialystok2011). However, it has also been challenged, with a number of studies and reviews not finding evidence for differences in behavioural performance between monolinguals and bilinguals (Antón et al., Reference Antón, Duñabeitia, Estévez, Hernández, Castillo, Fuentes, Davidson and Carreiras2014; Giovannoli et al., Reference Giovannoli, Martella, Federico, Pirchio and Casagrande2020; Kalashnikova et al., Reference Kalashnikova, Pejovic and Carreiras2021; Paap & Greenberg, Reference Paap and Greenberg2013), even when using advanced techniques or extensive sample sizes (Jones et al., Reference Jones, Davies-Thompson and Tree2021; Nichols et al., Reference Nichols, Wild, Stojanoski, Battista and Owen2020). Other studies have argued that any difference between groups can be attributed to confirmation and publication bias or to variables other than bilingual experience (Namazi & Thordardottir, Reference Namazi and Thordardottir2010; Paap, Reference Paap2016).
One way of reconciling the evidence for neurocognitive adaptation with the mixed behavioural data might be to assume that bilingualism creates a degenerate system (Green et al., Reference Green, Crinion and Price2006; Mason et al., Reference Mason, Domínguez D, Winter and Grignolio2015; Navarro-Torres et al., Reference Navarro-Torres, Beatty-Martínez, Kroll and Green2021). The concept of degeneracy is defined as structurally diverse components leading to the same output, or performing the same function. In contrast to the everyday meaning with negative connotations of decay, degeneracy in the scientific sense is a desirable characteristic, making systems robust and underpinning their capacity for adaptation (Mason et al., Reference Mason, Domínguez D, Winter and Grignolio2015; Whitacre & Bender, Reference Whitacre and Bender2010). Given that degenerate systems are functionally plastic, it is straightforward to assume that the brain has the capacity for degeneracy, and can provide reliable outputs despite inherent variation due to individual experience. In the context of the substantial variability and complexity of bilinguals' experiences (due to the well-established influences of the age of acquisition (AoA), proficiency, exposure, language similarity, usage context, etc. (Abutalebi et al., Reference Abutalebi, Cannini, Rosa, Shueng and Weekes2013; Nichols & Joanisse, Reference Nichols and Joanisse2016; Timmer et al., Reference Timmer, Wodniecka and Costa2021; Wei et al., Reference Wei, Joshi, Zhang, Mei, Manis, He, Beattie, Xue, Shattuck, Leahy, Xue, Houston, Chen, Dong and Lu2015), this will naturally lead to a non-trivial degree of variation in the patterns of bilingual adaptation, which could go some way towards explaining the mixed behavioural results seen in the literature.
Another key assumption behind the view that flexible adaptation to the demands of bilingualism reflects reconfiguration of the available resources, rather than their enhancement, is that this can be achieved in the context of finite selective attention capacity. This acknowledges that human performance is supported by a limited pool of attentional resources (Kahneman, Reference Kahneman1973), which can only process a restricted amount of information at any given point (Broadbent, Reference Broadbent1965; Clark & Dukas, Reference Clark and Dukas2003). Attentional systems are typically flexible enough to accommodate increased processing demands and maintain performance as long as total capacity is not exceeded (Moray, Reference Moray1967). Thus, even assuming that the process of selecting the target language and inhibiting the non-target one will itself utilise some of the existing attentional resources in bilinguals (Dornic, Reference Dornic1980; Wickens, Reference Wickens2007), this will still leave enough bandwidth for optimal performance and explain why bilinguals can accomplish single behavioural tasks with no apparent loss of performance relative to monolinguals. Indeed, it could be argued that neurocognitive adaptations in bilingualism emerge precisely in order to enable bilinguals to overcome the consumption of attentional resources by the competing languages, and still maintain optimal behavioural performance. Although underpinned by different mechanisms (reorganisation rather than enhancement), this would converge on the same widely accepted outcome of increased flexibility in the usage of the available resources in bilinguals, enabling them to do ‘more with less’.
Any attentional system, however, despite being flexible enough to accommodate parallel processing when attentional demands are not excessive, has limitations on the processing of multiple tasks. Contrary to many examples of ‘multitasking’ in daily life being achieved without noticeable deterioration of performance (such as e.g., driving and maintaining a conversation), researchers have found that dual tasks in experimental settings usually hamper performance, often resulting in slower processing of a second stimulus (Pashler & Johnston, Reference Pashler, Johnston and Pashler1998; Schubert, Reference Schubert2008; Telford, Reference Telford1931). This has been attributed to the need to execute both tasks' demands with a finite-capacity processor that can only process a restricted amount of information at any given point (Broadbent, Reference Broadbent1965; Clark & Dukas, Reference Clark and Dukas2003; Kahneman, Reference Kahneman1973) or a bottleneck preventing parallel processing at some stage (Pashler, Reference Pashler1994).
However, despite this long-standing premise that performance typically suffers to some extent under competing dual-task demands (Treisman, Reference Treisman1969), the way this may interface with bilingualism has been less clear cut. On the one hand, there have been studies showing a protective effect of bilingualism when managing competing tasks (Janic et al., Reference Janic, Cavanagh and Rivest2020; Telner et al., Reference Telner, Wiesenthal, Bialystok and York2008). These studies used a visual search (Janic et al., Reference Janic, Cavanagh and Rivest2020) and driving simulation (Telner et al., Reference Telner, Wiesenthal, Bialystok and York2008) as the primary tasks and counting backwards or verbal tasks as the distractor task. Another combination of tasks, memory recall and card-sorting (Sörman et al., Reference Sörman, Josefsson, Marsh, Hansson and Ljungberg2017), has suggested a more nuanced effect of bilingualism. Using this paradigm, a longitudinal study investigated the dual-task costs of encoding and retrieval of a list of words while participants performed a simultaneous card-sorting task. The results showed lower costs for bilinguals in one condition (recall with card-sorting at encoding and retrieval) but this was not consistent across other conditions (recall with card-sorting at encoding or at retrieval only) and across age groups. Another set of studies (Bialystok, Reference Bialystok, Craik and Ruocco2006; Bialystok et al., Reference Bialystok2011) investigated the effect of dual tasks on different modalities for monolingual and bilingual groups, using semantic categorisation across visual and auditory domains. In the first study by Bialystok et al. (Reference Bialystok, Craik and Ruocco2006), two semantic categorisations were required (of letters/numbers or animals/musical instruments) for both visual and auditory stimuli. Both younger and older bilingual adults showed an advantage in one section of the visual task; but not for an alternative version of the visual task, nor in the auditory task. In the second study by Bialystok (Reference Bialystok2011), two groups of 8 year olds performed one of the semantic categorisation tasks in two modalities simultaneously and the only difference between the groups was the accuracy, but not response time, in the categorisation of the visual stimuli. However, a separate study of younger and older adults (Fernandes et al., Reference Fernandes, Craik, Bialystok and Kreuger2007), combining memory recall of spoken words with a visual distraction task using semantically similar or unconnected words, found that bilingual participants in both groups performed significantly worse in all conditions. The authors attributed this bilingual limitation to the task's dependency on lexical access, which might advantage monolinguals who have a larger vocabulary in their primary (only) language. However, while noting that these existing studies on dual-task performance in bilinguals offer a mixed picture, it is also worth bearing in mind that they use different combinations of tasks that often span across different cognitive functions (e.g., visual search and counting backwards in Janic et al., Reference Janic, Cavanagh and Rivest2020, or driving and verbal tasks in Telner et al., Reference Telner, Wiesenthal, Bialystok and York2008), which makes it difficult to draw direct comparisons or achieve uniform results across studies.
1.2. Current study
Given the lack of clarity from the existing literature, the aim of the current study was to establish the impact of adding a secondary task on the behavioural selective attention performance in bilinguals. Building on the findings that the processing system is flexible and adaptive, allowing bilinguals to maintain optimal performance on a single task (Olguin et al., Reference Olguin, Cekic, Bekinschtein, Katsos and Bozic2019; Phelps et al., Reference Phelps, Attaheri and Bozic2022) we asked whether a further increase in processing demands may tax the system to the extent that it would lead to differentiation in performance on the secondary task, compared to monolingual controls. If so, this would strengthen the interpretation that bilingual adaptation reflects a distribution of the available (finite) capacity, rather than its enhancement.
To avoid some of the potential issues raised with the existing studies, we chose attentional tasks that were not explicitly dependent on vocabulary size, and whose combined attentional load could be varied incrementally. The primary task was a dichotic listening task (Cherry, Reference Cherry1953), where the target story was presented alongside varying levels of interference (from purely acoustic to linguistic) that needed to be inhibited to attend to the target story. This was the same design as used in our previous work (Olguin et al., Reference Olguin, Bekinschtein and Bozic2018, Reference Olguin, Cekic, Bekinschtein, Katsos and Bozic2019; Phelps et al., Reference Phelps, Attaheri and Bozic2022), which showed equivalent behavioural comprehension in monolinguals and bilinguals across all types of interference, but supported by a different configuration of neural activity in the two groups – which was interpreted as an adaptive modification of the neural mechanisms of selective attention in bilinguals, where the available capacity is redistributed to compensate for the increased processing demands while still enabling optimal behavioural performance. In the current study, the dichotic listening task was presented simultaneously with a simple visual task assessing inhibition and vigilance (Greenberg & Waldmant, Reference Greenberg and Waldmant1993). The dual task was deliberately designed so that the listening task would take priority over the visual task in conditions of divided attention. Interest was sustained in the listening task by using stories which were unpredictable and entertaining. Furthermore, target story narratives continued across testing blocks, but were presented with different levels of interference in the non-attended ear, in order to maintain interest while adding variety and a range of difficulty across the whole task. By contrast, the visual task was repetitive and simple; with the same stimuli presented in a random order within each block. Finally, in the dichotic listening task, participants were instructed explicitly to listen carefully in order to answer comprehension questions at the end of each block; whereas in the visual task there was no feedback or performance assessment.
Our primary interest in this study was in the performance of bilingual participants, as compared to their monolingual counterparts, across the two tasks and across different levels of interference. Following our earlier studies, participants were presented with different types of dichotic listening interference (no interference, acoustic interference, linguistic interference), thus manipulating the processing load required to separate the attended from the unattended streams (cf. multimode accounts of selective attention, Bronkhorst, Reference Bronkhorst2015; Johnston & Heinz, Reference Johnston and Heinz1978). We expected that this variation in the type of interference would not necessarily impact the dichotic listening performance in either group, replicating our existing findings (Olguin et al., Reference Olguin, Bekinschtein and Bozic2018, Reference Olguin, Cekic, Bekinschtein, Katsos and Bozic2019; Phelps et al., Reference Phelps, Attaheri and Bozic2022). However, the additional load of the dual task was expected to impact performance on the secondary task more strongly in bilinguals, as the system was potentially stretched beyond what can typically be accommodated by reconfiguration of the finite attentional resources. We also hypothesised that this impact on secondary task performance might interact with the level of interference, though it was unclear whether this would simply reflect the amount of competing information (with performance in all dual-task interference conditions slower than in conditions with no interference), or whether performance might further deteriorate with the increase of interference from purely acoustic to specifically linguistic.
The other main goal of the study was to test how bilingual adaptation might change with maturation. It has been well-established in the literature that age is one of the crucial factors determining behavioural performance of bilinguals relative to monolinguals, with their performance on single tasks difficult to differentiate in young adults (Bialystok et al., Reference Bialystok, Craik, Grady, Chau, Ishii, Gunji and Pantev2005; Valian, Reference Valian2015), but often diverging in childhood (Bialystok, Reference Bialystok2017; Carlson & Meltzoff, Reference Carlson and Meltzoff2008; Filippi et al., Reference Filippi, Ceccolini and Bright2022; Poulin-Dubois et al., Reference Poulin-Dubois, Blaye, Coutya and Bialystok2011) or late adulthood (Bak et al., Reference Bak, Nissan, Allerhand and Deary2014; Blumenfeld et al., Reference Blumenfeld, Schroeder, Bobb, Freeman and Marian2016; Gold et al., Reference Gold, Kim, Johnson, Kryscio and Smith2013). On the assumption that bilingual performance on selective attention tasks reflects adaptation in the usage of a finite-capacity resource, this trajectory could be explained as a process of learning to calibrate the usage of the available resources as the attentional system matures, initially operating in the context of a developing system with attenuated ability to allocate attention relative to adults (Ridderinkhof & van der Stelt, Reference Ridderinkhof and van der Stelt2000); followed by the optimally functioning system whose flexible usage of the available capacity has protective qualities in older age (Abutalebi et al., Reference Abutalebi, Guidi, Borsa, Canini, Della Rosa, Parris and Weekes2015; Bak & Robertson, Reference Bak and Robertson2017; Gold, Reference Gold2015; Wood, Reference Wood2016). Critically however, this view predicts that behavioural performance in the earlier stages of this learning-induced adaptation might be different to that of monolingual controls, particularly in the contexts of high processing load. In the current study we therefore used the same dual-task paradigm to test a sample of school-aged children, and compare their performance to that of young monolingual and bilingual adults. The children were in the 7–12 age bracket, reflecting the findings that tests of selective attention in younger children lead to inconsistent results and high variance in response speed and accuracy (Gomes et al., Reference Gomes, Duff, Barnhardt, Barrett and Ritter2007; Takio et al., Reference Takio, Koivisto, Jokiranta, Rashid, Kallio, Tuominen, Laukka and Hämäläinen2009); whereas the selected age range represents a developmental plateau for selective attention in childhood and can generate relatively stable effects, while ensuring that children can reliably perform a selective attention task (Phelps et al., Reference Phelps, Attaheri and Bozic2022).
2. Materials and methods
The experiment was conducted on two groups of participants: monolingual and bilingual children aged 7–12 (Study 1) and adults aged 18–45 (Study 2). Both studies were approved by the Cambridge Psychology Research Ethics Committee. The authors assert that all procedures contributing to this work comply with the ethical standards of the relevant national and institutional committees on human experimentation and with the Helsinki Declaration of 1975, as revised in 2008.
Study 1 participants: Eighty typically developing children aged 7–12 were tested. They were grouped into two categories: monolingual (n = 40, 25 males, mean age 10.4 years, standard deviation [SD] = 1.64) and bilingual (n = 40, 22 males, mean age 9.7 years, SD = 1.39). All participants were healthy with no history of hearing problems or neurological disorder. All participants' parents completed a language history questionnaire, which provided an overview of children's exposure to languages. As confirmed by the questionnaire, all monolingual participants were native speakers of English, with no significant exposure to other languages. The participants in the bilingual group all had a similar profile: they all had one or both parents whose first language was not English, and they used this language at home on a daily basis. The average age of acquisition for the language other than English was .1 years (age 0 for 38 participants, age 2 for 2 participants). All children were also fluent in English, being resident in the UK, and following English-speaking curriculum at school. The average age of acquisition for English was .3 years (from birth for 32 participants, age 1 for 5 participants, age 2 for 2 participants, age 3 for 1 participant). The second languages spoken were Creole, French, German, Greek, Gujarati, Hungarian, Japanese, Italian, Portuguese, Russian, Spanish and Swedish. Additionally, three children spoke a third language proficiently (French, Japanese and Urdu). Children were recruited via their parents, who responded to posters on social media, or word of mouth. The monolingual children were marginally older on average than the bilingual group [10.4 vs 9.7 years respectively, t(76) = 2.06, p = .043], and age was included as a predictor in all analyses. Parental educational information was collected as an indication of socio-economic status (SES), a well-documented influence on selective attention in children (Stevens et al., Reference Stevens, Lauinger and Neville2009). The majority of participants' parents (79.2%) were educated to degree level or higher, and the groups were not significantly different on this approximation of SES (bilinguals: M = 2.34, SD = .66; monolinguals: M = 2.09, SD = .69; Mann–Whitney Wilcoxon test: W = 608.5, p = .085). We also included SES as a predictor in the analyses throughout. In summary, our monolingual participants had no significant exposure to other languages, while all bilingual children were fully proficient in English and another language; had daily exposure to both and were using both daily (English at school, English and/or the other language at home and with family).
Study 2 participants: Eighty-nine adults aged 18–45 were recruited, five of whom were rejected after initial quality checks, leaving 84 participants in the analyses (see details of rejection criteria in Section 2.2). They formed a monolingual (n = 42, 15 males, mean age 28.9 years, SD = 7.29) and a bilingual group (n = 42, 6 males, mean age 27.4 years, SD = 5.95) and matched on age (t(78.9) = 1.01, p = .32). All participants were neurotypical with no history of hearing problems or neurological disorder. They completed a language questionnaire on age of acquisition, proficiency and daily usage of their languages. All monolingual participants were native speakers of English with no fluency in other languages. All bilingual participants were resident in the UK; fluent and highly proficient English speakers (average age of English acquisition = 6.1 years; average English proficiency = 9.3/10, SD = .96); and using English on a daily basis (average English use = 73%, SD = 24.9), but with a language other than English as their native language. The native languages spoken by the bilingual participants were Afrikaans, Bengali, Danish, Farsi, Filipino, Finnish, French, Galician, Greek, Gujarati, Hindi, Igbo, Indonesian, Italian, Konkani, Latvian, Lithuanian, Malayalam, Persian, Polish, Portuguese, Romanian, Russian, Scots, Spanish, Turkish and Welsh. In addition to English as a second language, many participants spoke a third language proficiently (Albanian, Catalan, Dutch, French, German, Hindi, Icelandic, Italian, Japanese, Russian, Russian Sign Language, Spanish, Tamil, Telugu, Urdu and Xhosa), several a fourth (Annang, French, Italian, Polish, Portuguese, Spanish and Swedish) and one a fifth (Japanese). Highest level of education was collected as an indication of SES, and the groups were comparable on this approximation (monolinguals: M = 1.5, SD = .59; bilinguals: M = 1.81, SD = .74; W = 685, p = .053). The key characteristics of our adult participants were therefore comparable to those of children: monolinguals were English speakers with no fluency in other languages; whereas bilinguals were UK residents fully proficient in English as well as in their native language; exposed to both on a daily basis, and using both daily. Adult participants were recruited via the online platform Prolific (https://www.prolific.co/).
2.1. Design
In this dual-task paradigm, participants were required to perform tasks simultaneously in two modalities: auditory and visual (Figure 1a). The visual task demanded pressing the space bar in response to a pre-specified target image on the screen. The auditory task employed a dichotic listening paradigm, where participants were asked to concentrate on a story in English played in one ear, while ignoring a distractor steam presented in the other ear. We manipulated the distractor stream to create different levels of interference. This created a total of five conditions, which are summarised in Table 1.

Figure 1. (a) Dual-task procedure. Participants were instructed to attend to a story in one ear and ignore a distractor stream in the other ear, while also responding to a visual task. The distractor streams were manipulated to create different levels of interference. In the visual task, a picture of a dog at the top of the screen was the target, while a picture of a dog at the bottom of the screen was the distractor. After each block of the audio–visual task participants answered comprehension questions about the target story. (b) Dichotic listening task results from Study 1 (children). Data show that both groups performed the comprehension task equally well. (c) Dichotic listening task results from Study 2 (adults). There was again no difference in performance between the two groups.
Table 1. Experimental conditions

In the first condition (‘Control’), the participants only performed the visual task. From condition 2 onwards, the participants were also presented with the dichotic listening task, and asked to attend to a target story in English in one ear. Condition 2 had no interference in the other ear (‘Single Talker’). In the remaining three conditions, participants also heard interference in the other ear, which they were instructed to ignore. In condition 3 the interference was a different story in English (‘English–English’). The interference in condition 4 was a story in Latin, a language they could not process for meaning (‘English–Latin’). The interference in condition 5 was non-linguistic acoustic signal (‘English–Musical Rain’ [MuR]).
The target stories were two children's stories in English, taken from online resource storynory.com. Both stories were transcribed into 120 sentences each, with each sentence lasting approximately 3 seconds in length. Each target story was split into two blocks and participants attended to the first half in one ear, with interference (as determined by the condition) in the other ear, and swapped accordingly for the second half. Each block (half of a story) consisted of 60 sentences, which were concatenated with a 300 ms gap between them to create a single block per condition lasting 3.3 min. Latin was chosen as the interference in condition 4 as a non-artificial language which would most likely be unknown to the participants. Gender of the speaker was kept the same for all stories to reduce segregation strategies based on talker's gender (Brungart & Simpson, Reference Brungart and Simpson2007); but different female speakers were used for attended and unattended stories in conditions 3 and 4, allowing listeners to maintain separation with high accuracy (Zekveld et al., Reference Zekveld, Rudner, Kramer, Lyzenga and Rönnberg2014). All stories' volumes were normalised to ensure equivalent average amplitude. The non-linguistic interference of Musical Rain in condition 5 was identical in length, root mean squared level and long-term spectrotemporal distribution of energy to the corresponding target story, but did not trigger a speech percept (Uppenkamp et al., Reference Uppenkamp, Johnsrude, Norris, Marslen-Wilson and Patterson2006). It was generated in MATLAB by extracting temporal envelopes of the target sentences and filling them with 10 ms fragments of synthesised vowels jittered in frequency and periodicity. Instructions were recorded by the same female speaker of the target stories. These were played before each block in the target ear, telling the participant: ‘This is your right/left ear. Please listen carefully to the story in this ear, on your right/left side, and ignore the story or sound in the other ear’.
After each block had finished, a screen appeared with 10 comprehension questions in multiple-choice format. Comprehension questions consisted of simple written sentences to check understanding of each story. Participants did not receive feedback on their responses.
Visual task: The visual task required pressing the space bar as quickly as possible when the cartoon picture of a dog appeared at the top of the screen. If the dog appeared at the bottom of the screen, the participant was instructed to ignore and wait for the next trial. Each dog image was on the screen for a maximum of 2000 ms, with a blank screen lasting 500 ms presented between the trials. If the participant detected the target correctly, the reaction time was recorded and the next trial began immediately. If the participant failed to detect the target, or pressed the space bar when the dog was at the bottom of the screen, the trial was recorded as error.
The ratio of target:non-target stimuli was 1:3 with the 24 targets and 72 non-targets appearing in a random order in each block. These 96 visual stimuli aligned with the length of a block of 60 sentences; however, if a participant was responding very fast and completed the visual task before the end of the auditory stream, extra visual stimuli were presented in the same 1:3 ratio until the story ended.
Procedure: The experiment was run online, requiring the use of a PC or a laptop with keyboard and headphones. Prior to testing, all participants (and their parents in Study 1) were given detailed information on the aims of the project and what to expect from the session. They then completed a consent form and a background questionnaire on their demographic information and language history (this was completed by children's parents in Study 1). Study 1 then also presented participants with a short video explaining the rules of both tasks in child-appropriate language. Here, the visual task was called ‘Spot the Dog’ and the auditory task was referred to as ‘the listening task’. Before starting the experiment, participants were also reminded that they could withdraw from the study at any time. We also ran a short headphone check to ensure that participants could hear the spoken narratives clearly and at a comfortable level. The experiment started with a practice session (which could be repeated if the participant needed) and was followed by condition 1 (control) first and then the dual-task conditions in fixed order (single talker, English–English, English–Latin, English–MuR). This ensured that the first half of attended stories was always followed by the second half, and that participants remained engaged throughout the study by following the attended narratives across conditions. A review of participants' comments and data distribution on competition confirmed that this was a successful strategy that did not interfere with the response patterns across conditions. The task was prepared and presented in PsyToolkit (Stoet, Reference Stoet2010, Reference Stoet2017). The study lasted approximately 30 min, and participants were compensated for their time.
2.2. Analyses
Dichotic listening task: At the end of each block participants answered 10 comprehension questions about the attended narrative. Correct answers were scored as 1 and errors as 0, and their probability across groups and conditions was modelled using a glmer function from the binomial family in R (RStudio Team, 2020).
Visual task: Reaction times and error rates were recorded. As an initial quality check, all outlier reaction times under 200 ms and over 1500 ms were eliminated. The reaction times were then log transformed to eliminate skew and datapoints more than 1.5 interquartile ranges above the upper quartile or below the lower quartile were removed as outliers. This resulted in exclusion of 129/8860 datapoints (1.5% of the total) in Study 1 (children) and 302/9724 datapoints (3.1% of the total) in Study 2 (adults). Error rates were calculated per participant, including both missing the target (not pressing the space bar when the target was presented) and incorrect detection of targets (pressing the space bar when the target was not presented). The initial quality check of total error rates in Study 1 did not identify any children whose error score exceeded the normal range for attention task in that age group (maximum error rate was 10%; normal range in neurotypical children aged 6–13 performing the task is .8–14%, Greenberg & Waldmant, Reference Greenberg and Waldmant1993), hence the data of all 80 children were included in the analyses. In Study 2 (adults), a check of total error rate identified two monolingual and two bilingual participants whose total error rates significantly exceeded 3 SD of the mean; likely indicating that they did not attend to the task as instructed. One further monolingual participant's language questionnaire revealed proficiency in a second language, acquired in adulthood. The data of these five participants were excluded, leaving 84 participants' data for further analysis.
The reaction times of correct target responses were compared across groups and conditions using linear mixed-effects models (Baayen et al., Reference Baayen, Davidson and Bates2008) as implemented in the lme4 R package (Bates et al., Reference Bates, Mächler, Bolker and Walker2015), with the emmeans function used for post-hoc comparisons. Checks of models' assumptions (linearity, homoscedasticity, normality) did not indicate significant violations. The Satterthwaite approximation (Satterthwaite, Reference Satterthwaite1946) was used for degrees of freedom. Significant p-values are reported at p < .05.
3. Results
3.1. Dichotic listening task
3.1.1. Study 1: children
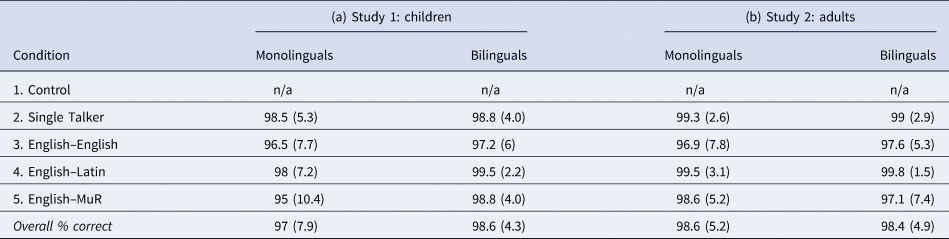
Children from both groups performed the task equally well, with overall comprehension scores of 97% in the monolingual group, and 98.6% in the bilingual group (Table 2a). To test whether the probability of making an error differed across groups and conditions we ran a model with categorical response accuracy as the dependent variable, fixed effects of group (two levels: monolingual, bilingual), condition (four levels: single talker, English–English, English–Latin and English–MuR) and their interaction, participant age and parental SES; and participants as random intercept. The results showed a slight overall drop in performance in condition 5 (odds ratio [OR] = .27, confidence interval [CI] = .10–.71, p = .008), but no effects of group or group × condition interaction (all p > .1), confirming that both groups performed the auditory task equally well. Age was a significant predictor (OR = 1.59, CI = 1.24–2.05, p < .001). A summary of comprehension scores and standard deviations for auditory task performance in children is shown in Table 2a and Figure 1b; and the full model for the auditory comprehension data in children is presented in Table A1a in the Appendix.
Table 2. Comprehension scores (% correct) and standard deviation across groups and conditions in the dichotic listening task for (a) Study 1: children and (b) Study 2: adults
3.1.2. Study 2: adults
Comparable to the results from Study 1, adults from both groups performed the task equally well, with overall comprehension scores of 98.4% in the monolingual group, and 98.5% in the bilingual group (Table 2b). A model with categorical response accuracy as the dependent variable; group (monolingual, bilingual), condition (four levels) and their interaction, age and SES as fixed effects, and participants as random intercept, indicated a slight drop in performance in condition 3 (OR = .21, CI = .06–.77, p = .018); but again showed no effects of group or group × condition interaction. A summary of comprehension scores and standard deviations for adults is shown in Table 2b and Figure 1c, and full model is given in Table A1b in the Appendix.
3.2. Visual task
3.2.1. Study 1: children
Accuracy: The total error rate was 1.9% for monolingual children and 2.07% for bilingual children, which did not significantly differ between the groups [t(77.7) = −.36, p = .72]. There were too few errors across all participants and conditions in Study 1 (762/>40,000 datapoints) to allow for any finer-grained analyses.
Reaction times: Response times (RTs) across conditions for monolingual and bilingual children are shown in Table 3a. To test whether RTs for correctly detected targets differed between groups and conditions, we ran a model with log-transformed reaction times (log RTs) as the dependent variable, the fixed factors of group (two levels: monolingual, bilingual), condition (five levels: 1. control; 2. single talker; 3. English–English; 4. English–Latin; 5. English–MuR), their interaction, participant age and parental SES; and participants as random intercept. Results (Table 4a) showed that condition was a significant predictor, with RTs in all conditions significantly slower than in the control condition (all p < .001). Although the main effect of group was not significant (b = −.01, CI = [−.08 to .05], p > .1), there was a significant condition × group interaction in conditions 3–5, driven by slower RTs in the bilingual group. Post-hoc comparisons showed no significant differences between the effects in these three conditions in bilinguals (all p > .1; Table A2 in the Appendix). Data also showed that age was a significant predictor of RTs across both groups of children (b = −.05, CI = [−.07 to −.03], p < .001). A full summary of the results is presented in Table 4a, and displayed in Figure 2a.
Table 3. Mean reaction time (ms) and standard deviation across groups and conditions in the visual task for (a) Study 1: children and (b) Study 2: adults

Table 4. Model summary for RTs in (a) Study 1: children, and (b) Study 2: adults

Bold denotes a statistically significant result.

Figure 2. Visual task results from (a) Study 1: children; (b) Study 2: adults and (c) across the two studies. The results reveal that bilingual children are disproportionately affected by interference conditions 3–5 compared to all other groups. Asterisks indicate the presence of significant interactions; ***p < .001; **p < .01.
3.2.2. Study 2: adults
Accuracy: The total error rate was .66% for monolinguals and .59% for bilinguals, which was not significantly different between the groups [t(81.5) = −.51, p = .61]. Because of too few errors across all participants and conditions in Study 2 (250/>40,000) we did not run any further error analyses.
Reaction times: RTs across conditions for monolingual and bilingual adults are shown in Table 3b. Following the same procedure as in Study 1, we tested whether group and condition were significant predictors of RTs to correctly detected targets in the two groups. The dependent variable was log-transformed RTs, the fixed factors were group (monolingual, bilingual), condition (five levels) and their interactions, participant age and SES; with participants as random intercept. As in Study 1, results showed that condition was a significant predictor, with all conditions significantly slower than the control condition (all p < .001, Table 4b). This time however neither the effect of group (b = .03, CI = [−.02 to −.09], p > .1) nor condition × group interaction were significant (all p > .1), suggesting no modulation of this pattern across the two groups (Table 4b and Figure 2b).
3.2.3. Supplementary Bayesian analyses of visual task RTs
The analyses reported so far used standard Null Hypothesis Significance Testing (NHST), revealing no differences in response patterns between monolingual and bilingual adults, but slower visual task RTs in dual-task interference conditions for bilingual children. However, NHST statistics does not distinguish whether findings of no difference reflect a genuine absence of difference or insensitivity of the data (Dienes, Reference Dienes2014), and, given the significance of this point for our interpretation, we supplemented our NHST analyses with Bayesian analyses of variance (JASP version 0.17 with default priors). For both children and adults, fixed factors were group and condition, and participant was a random factor. We derived the inclusion Bayes factors for the effects of group and condition and their interaction, quantifying the evidence for including them as predictors. The output of these analyses is presented as Table A3 in the Appendix. As expected, there was very strong evidence for the inclusion of condition in both groups. The evidence for the inclusion of group and group × condition interaction however was much weaker, and also different between children and adult datasets. In children, we saw anecdotal evidence for the inclusion of group (BFincl = 1.33) but stronger evidence for group × condition interaction (BFincl = 3.47), replicating our original results that condition significantly interacted with group to determine patterns of responses in this cohort. In adults, there was no evidence for the inclusion of either group (BFincl = .19) or the group × condition interaction (BFincl = .00013), suggesting that monolingual and bilingual adults were indeed not different from each other.
3.3. Comparison of visual data across studies 1 and 2
The final set of analyses compared the visual RT data across the two studies for monolingual and for bilingual participants separately, to contrast directly their performance across the two age groups and assess whether visual task results change with maturation. As the RTs in children were slower than in adults, we expected that study would be a significant predictor in the analyses of both monolingual and bilingual groups. We also expected to see condition as a significant predictor in both groups, reflecting the finding that the control condition was consistently faster than other conditions in all previous analyses. The key question was therefore whether study and condition would interact in either monolingual or bilingual group, suggesting a differential pattern of responses in children and adult participants as a function of their language status.
In monolinguals, a model included fixed factors of study (adults, children), condition (five levels) and their interaction, participant age and SES; with participant as random intercept. As expected, results showed significant effects of both study (children slower than adults, b = .17, CI = [.03–.31], p < .01) and condition (control faster than other conditions, all p < .01), however there was no interaction between them (all p > .1), indicating that both groups responded comparably to all five conditions (Table 5a). In bilinguals, the equivalent model also revealed that study and condition were significant predictors (children slower than adults, b = .18, CI = [.05–.31], p < .01; control faster than other conditions, all p < .001), but there was also an additional robust interaction between the two in all dual-task conditions (Table 5b). Figure 2c displays a summary of mean RTs for participants from all groups, clearly showing that the responses of monolinguals followed the same pattern across conditions in both children and adults; whereas bilingual children were disproportionately affected by dual-task conditions compared to all other groups.
Table 5. Model summary for reaction times in monolinguals and bilinguals across studies 1 and 2

Bold denotes a statistically significant result.
Taken together, these two sets of results show a difference in language groups' performance on a secondary task, which is discernible in childhood, but not present in adulthood. The implications of these findings are discussed below.
4. Discussion
The current study investigated the performance of monolingual and bilingual listeners in a dual selective attention task. Starting from the assumption of functional plasticity and degeneracy – where the increased processing demands of competing languages in the bilingual brain are expected to induce flexible adaptation in the usage of a finite attentional capacity – our primary aim was to examine how the available resources are employed to support performance under high processing loads. To assess this, we measured participants' performance on two simultaneous tasks: a dichotic listening task with comprehension scores as indicator of attentional performance; and a visual attention task, in which we tracked both the accuracy and the speed of responses. The second aim of the study was to test how this adaptation to high processing loads might change with maturation and experience. To this end, we conducted the experiment on two groups: school-age children (age range 7–12) and adults (age range 18–45). Our results showed equivalent performance on the dichotic listening task for monolinguals and bilinguals in both age groups, with all participants achieving near perfect comprehension scores across all interference conditions. For the visual attention task, accuracy was again equivalent for monolinguals and bilinguals in both age groups, with very low error rates across the board. Reaction times, however, revealed a trend for slightly slower responses in bilinguals, with this being particularly prominent in the dual-task interference conditions in bilingual children. We discuss these results in more detail below.
4.1. Bilingual adaptation to increased processing demands
It is widely accepted that the demands of managing competing languages trigger adjustments of the underlying neurocognitive architecture in bilinguals (Abutalebi & Green, Reference Abutalebi and Green2016; Burgaleta et al., Reference Burgaleta, Sanjuán, Ventura-Campos, Sebastian-Galles and Ávila2016; Hayakawa & Marian, Reference Hayakawa and Marian2019). We argue that this represents just one example of adaptation of a biological system to changes in the environment, in line with the concepts of functional plasticity and degeneracy (Green et al., Reference Green, Crinion and Price2006; Mason et al., Reference Mason, Domínguez D, Winter and Grignolio2015; Navarro-Torres et al., Reference Navarro-Torres, Beatty-Martínez, Kroll and Green2021). With adaptation underpinning most human behaviours to enable adjustment to different environments and their demands (e.g., Barkow et al., Reference Barkow, Cosmides and Tooby1992), it follows that the processing patterns in monolinguals – rather than being a normative benchmark – are also just another example of adaptation, this time to the unique set of communicative processing demands that monolingual speakers encounter in their environments.
In the domain of bilingual selective attention – where the demands of selection and inhibition of the non-target language can themselves utilise some of the existing capacity – this adaptation will arguably lead to engagement of the remaining resources in a way that best supports current task requirements. Even in the context of a single selective attention task in which the processing demands are unlikely to exceed the capacity of the system, and where bilinguals and monolinguals perform equivalently, there is evidence that this behavioural performance is underpinned by a differentiable configuration of the underlying resources in the two groups (Filippi & Bright, Reference Filippi and Bright2023; Olguin et al., Reference Olguin, Cekic, Bekinschtein, Katsos and Bozic2019; Phelps et al., Reference Phelps, Attaheri and Bozic2022). In the current dual-task context we again saw that optimal behavioural performance in the primary task was maintained such that both monolingual and bilingual participants were able to perform dichotic listening without any issues. This result aligns with numerous findings from the literature that reported no evidence for enhanced performance in bilinguals, even when using advanced techniques or extensive sample sizes (Nichols et al., Reference Nichols, Wild, Stojanoski, Battista and Owen2020; Samuel et al., Reference Samuel, Roehr-Brackin, Pak and Kim2018). It is also consistent with the interpretation about a flexible system capable of adapting to changes in the environment, equivalent to learning-induced changes seen across domains as diverse as learning to juggle or read, memorising a sequence or acquiring detailed spatial knowledge (Bassett et al., Reference Bassett, Yang, Wymbs and Grafton2015; Dehaene et al., Reference Dehaene, Cohen, Morais and Kolinsky2015; Draganski et al., Reference Draganski, Gaser, Busch, Schuierer, Bogdahn and May2004; Maguire et al., Reference Maguire, Gadian, Johnsrude, Good, Ashburner, Frackowiak and Frith2000). The analysis of the results from the secondary visual attention task, however, revealed a more nuanced picture, with some aspects of performance on this task comparable between the groups, and others indicative of bilinguals using the available resources differently relative to monolingual participants.
One aspect of visual task performance that was comparable across groups was that reaction times in condition 1 (control) were consistently and significantly faster than in any other condition. Given that condition 1 was the only one where participants performed a single (visual) task, while all other conditions included both visual and auditory components, this result replicates the well-established finding that increasing competition for the limited capacity resource leads to deterioration in performance (Osman & Moore, Reference Osman and Moore1993; Pashler & Johnston, Reference Pashler, Johnston and Pashler1998; Schubert, Reference Schubert2008). Another aspect of visual task performance that was consistent across groups was that there was no deterioration in accuracy across the board: all groups displayed very low error rates, which could not be distinguished between monolinguals and bilinguals in either age group. However, the element of visual task performance where differences started to emerge was in the overall speed of responses, which indicated a trend towards slower responses for bilinguals (mean of 452 ms across all conditions and both age groups for monolinguals, and 476 ms for equivalent in bilinguals), and was significantly slower for bilingual than for monolingual children in dual-task conditions with interference. Thus, a hierarchy was revealed in which comprehension in the auditory task and accuracy in the visual task were arguably prioritised and optimal performance maintained. This produced a difference between monolingual and bilingual participants in the speed of responses in the visual task. We interpret this result as directly reflecting the economising of the available attentional resources under high processing load in bilinguals, where some of this capacity has already been utilised by the demands of selection and inhibition of the non-target language. Although this interpretation of the current behavioural data is necessarily functional, it is fully consistent with the existing neuroimaging evidence for flexible functional reorganisation of selective attention in bilinguals (Olguin et al., Reference Olguin, Cekic, Bekinschtein, Katsos and Bozic2019; Phelps et al., Reference Phelps, Attaheri and Bozic2022). The finding that RTs were particularly affected in conditions of dual-task interference is also consistent with this interpretation, as the impact of any reduction in capacity can be expected to be particularly visible when the processing demands are very high. In addition, this effect was broadly comparable across different interference conditions, suggesting that it is the presence of interference in itself (i.e., the increase in the amount of competing information) – rather than its type – that taxes the system beyond what it can accommodate. Alternatively, this latter finding might also reflect the choice of task in the current study, where the content of the interfering stream was not directly relevant for visual target detection.
One explanation behind this apparent hierarchy of responses across tasks (auditory/visual) and their components (speed/accuracy) is that the auditory task was deliberately designed as the primary task. It was more varied and entertaining than the visual task, which was simple and repetitive, thus capturing more attention through content and presentation even without explicit directive. For the visual task, the results are indicative of a speed-accuracy tradeoff (SAT), a mechanism that allows response accuracy to be maintained at the expense of taking longer to respond (Heitz, Reference Heitz2014). The decision criteria that underpin SAT have been previously linked to attentional capacity and processing load (Heitz & Engle, Reference Heitz and Engle2007; Lavie, Reference Lavie1995); complementing the interpretation that the high processing load of the current task combined with a more heavily taxed attentional capacity in bilinguals leads to a different pattern of responses in this group compared to the monolingual controls, such that RTs are fractionally longer – but not any less accurate. This is also reflected in different patterns of results across conditions in monolingual and bilingual children (Table A2 in the Appendix), where bilinguals showed a more graded pattern (single task < dual task without interference < dual task with interference) compared to monolinguals (single task < dual task); arguably reflecting their heightened sensitivity to changes in task demands, that is necessary for optimised attentional allocation.
This proposed explanation of the processes behind the current results contrasts with the theoretical position where bilingualism is argued to lead to an enhanced attentional capacity and therefore superior behavioural performance on tasks requiring attentional control (Bialystok, Reference Bialystok2015, Reference Bialystok2017; Comishen et al., Reference Comishen, Bialystok and Adler2019; Timmer et al., Reference Timmer, Wodniecka and Costa2021). Even assuming that our task was simply not challenging enough, and thus not capable of separating attentional performance in participants at the peak of their capacities (Bialystok, Reference Bialystok2017), the observed trend for slower responses in bilinguals across the board (Table 3) would pose a problem for this account. Instead, the results indicate a more stretched attentional capacity that is adapted to meet additional language demands and reach optimal performance by adulthood, consistent with the account of increased flexibility in the usage of the available resources in bilinguals (Abutalebi et al., Reference Abutalebi, Della Rosa, Green, Hernandez, Scifo, Keim, Cappa and Costa2012; Kousaie & Phillips, Reference Kousaie and Phillips2012).
4.2. Changes to bilingual adaptation with maturation and experience
The neural mechanisms of selective attention have been linked to a network of mostly prefrontal brain areas (Salo et al., Reference Salo, Salmela, Salmi, Numminen and Alho2017), with tests of selective attention such as the Flanker and Stroop – where the participant must suppress distracting information in order to focus on a target – identifying frontal lobes and the anterior cingulate cortex as areas supporting monitoring of attentional conflict and conflict resolution, respectively (Botvinick et al., Reference Botvinick, Cohen and Carter2004; Walsh et al., Reference Walsh, Buonocore, Carter and Mangun2011). Most importantly for the research presented here however, these neural systems have a notably protracted period of structural and functional development from infancy (Giedd et al., Reference Giedd, Blumenthal, Jeffries, Castellanos, Liu, Zijdenbos, Paus, Evans and Rapoport1999; Gogtay et al., Reference Gogtay, Giedd, Lusk, Hayashi, Greenstein, Vaituzis, Nugent, Herman, Clasen, Toga, Rapoport and Thompson2004; Tsujimoto, Reference Tsujimoto2008). The consequences of this on behavioural performance are such that children can display attenuated ability to allocate attention relative to adults, assigning proportionately more attention to irrelevant stimuli at the expense of relevant stimuli in selective attention tasks (for a review see Ridderinkhof & van der Stelt, Reference Ridderinkhof and van der Stelt2000).
Our results across the two studies align with this evidence, while also adding further detail on how a developing bilingual system accommodates heavy processing loads of a dual task. This task, tapping two competing domains, deliberately increased processing load beyond the boundaries of typical demands on selective attention. As argued above, the patterns of RT responses we saw suggest that the available attentional resources in bilingual children were distributed in response to the tasks' demands, maintaining optimal performance in the task of higher priority, at the expense of performance on aspects of the lower priority task. Importantly however, the performance limitations in the lower priority task largely appear to recede by adulthood. This suggests a developmental trajectory in which the resource allocation strategies fine-tune with development and maturation of the prefrontal cortex associated with selective attention – allowing bilinguals to optimise the process of learning to distribute the available resources to interface efficiently with the environment. In the context of our task, this suggests that maturation of the selective attention system enables adaptation to the combined load of second-language processing and competing task demands; resulting in optimal selective attention processing and performance in ‘extreme’ tasks too. It is necessary to note however that our study was cross-sectional and that, in the absence of longitudinal data, any conclusions about developmental trajectories of these processes are necessarily tentative. In addition, the effects of age and maturation on learning how to use resources efficiently in the bilingual context are most likely non-linear; further underpinning the need to characterise these processes longitudinally in future research.
Finally, although the current study focused on attentional capacity as a driver of performance on the dual task, it is clear that cognitive factors are only one of the several variables that influence behavioural outcomes in bilinguals (and monolinguals alike). Factors such as AoA, language similarity, usage context and trajectory of language development have all been shown to affect bilinguals' performance (Abutalebi et al., Reference Abutalebi, Cannini, Rosa, Shueng and Weekes2013; De Houwer, Reference De Houwer2021; Timmer et al., Reference Timmer, Wodniecka and Costa2021; Wei et al., Reference Wei, Joshi, Zhang, Mei, Manis, He, Beattie, Xue, Shattuck, Leahy, Xue, Houston, Chen, Dong and Lu2015), and these variables are likely to be interacting with the cognitive factors to shape the patterns of adaptation and processing in bilingualism. Although disentangling such interactions is complex (Kersten & Greve, Reference Kersten, Greve, Kersten and Winsler2022) and beyond the scope of this paper, it is arguably only the adaptation to this complexity of bilingual experience that can fully capture and explain performance outcomes, and account for the variability of findings in the bilingualism literature.
5. Conclusions
This set of studies investigated the effects of increased processing demands on bilingual adaptation across two age groups. The results revealed a pattern indicative of distribution of the available capacity and task prioritisation in bilingual children, relative to their monolingual counterparts, with these differences between monolingual and bilingual performance essentially receding by adulthood. This suggests that adaptation to the combined load of second-language processing and competing task demands fine-tunes as the selective attention system matures; resulting in optimal selective attention processing in bilinguals even under very high processing loads.
Data availability statement
The datasets generated and analysed in the current study are available on request from the first author.
Acknowledgements
We thank anonymous reviewers for feedback on previous versions of this manuscript, and participants for taking part in our study. We are also grateful to Brandon Lino and Qamil Pajaziti for their help in implementing the visual task. This research was supported by funds from the Department of Psychology, University of Cambridge.
Author contributions
J. P. and M. B. designed the research; J. P. performed the research and analysed the data; J. P. and M. B. wrote the manuscript.
Competing interests
The authors declare no competing interests.
Appendix
Table A1. Model summary for auditory comprehension data in (a) Study 1: children, and (b) Study 2: adults.

Table A2. Summary of post-hoc comparisons for visual RTs across conditions for monolingual and bilingual children separately (corrected for multiple comparisons)

Table A3. Supplementary Bayesian analyses of visual RTs for children and adults












 Open access
Open access