


Implications
Subacute ruminal acidosis (SARA) is a common metabolic disorder in cows fed high-grain diets and has been reported to elicit perturbations of the blood metabolome. Feed additives, such as clay minerals (CM), might be a management tool to reduce the negative consequences of high-grain feeding in dairy cows. This study showed that cows experiencing SARA and receiving a CM-based product had a higher concentration of several amino acids (AA) and phosphatidylcholines (PC) in blood, while several harmful biogenic amines (BA) and liver enzymes were lowered compared to control cows. This implies that a CM-based product holds the potential to counteract SARA-induced perturbations of systemic health.
Introduction
High-grain diets have become common in the feeding of cattle worldwide. These diets are characterized by their high content of rapidly fermentable carbohydrates in the rumen, thereby stimulating the generation of short-chain fatty acids that provide metabolic fuels. However, the inclusion of high amounts of grain occurs at the expense of fiber, which decreases chewing activity, salivary buffer supply and rumen motility (Humer et al., Reference Humer, Petri, Aschenbach, Bradford, Penner, Tafaj, Südekum and Zebeli2018a). Consequently, these diets enhance the risk of disrupted acid–base regulation in the rumen, resulting in long-lasting intermittent drops of ruminal pH below 5.8, designated as SARA (Zebeli and Metzler-Zebeli, Reference Zebeli and Metzler-Zebeli2012). The SARA has evolved to be one of most common digestive disorders causing major economic losses in dairy farming worldwide (Enemark, Reference Enemark2008). Besides local alterations in the rumen metabolome (Saleem et al., Reference Saleem, Ametaj, Bouatra, Mandal, Zebeli, Dunn and Wishart2012), more recent research has also revealed that SARA is associated with major perturbations in the systemic metabolism (Zebeli and Metzler-Zebeli, Reference Zebeli and Metzler-Zebeli2012; Guo et al., Reference Guo, Chang, Zhang, Xu, Jin, Bilal and Shen2017), increasing the risk of metabolic disorders and being a serious welfare concern.
The use of high-throughput technologies such as metabolomics has recently improved both diagnosis and our understanding in the etiopathogenesis of metabolic disorders in cattle (Hailemariam et al., Reference Hailemariam, Mandal, Saleem, Dunn, Wishart and Ametaj2014; Kenéz et al., Reference Kenéz, Dänicke, Rolle-Kampczyk, von Bergen and Huber2016). A recent study conducted by our team exploring the effects of grain-induced SARA on the blood metabolomic profile revealed large shifts in the plasma metabolome of dairy cows toward a decrease in lysophosphatidylcholines, PC, sphingomyelins and most AA (Humer et al., Reference Humer, Kröger, Neubauer, Schedle, Reisinger and Zebeli2018b).
Despite the research progress made in the etiopathogenesis and diagnostic tools of SARA in dairy cattle, both prevention strategies and the consequences of SARA for systemic cow health are still largely unresolved (Humer et al., Reference Humer, Petri, Aschenbach, Bradford, Penner, Tafaj, Südekum and Zebeli2018a and Reference Humer, Aschenbach, Neubauer, Kröger, Khiaosa-ard, Baumgartner and Zebeli2018c). Supplementation of feed additives has been suggested as a tool to reduce negative effects of high-grain feeding on health and to optimize rumen fermentation in cows at high risk of SARA (Krause and Oetzel, Reference Krause and Oetzel2006; Humer et al., Reference Humer, Petri, Aschenbach, Bradford, Penner, Tafaj, Südekum and Zebeli2018a). One of these feed additives which are less explored in cattle but long used to improve human health, including gut and systemic health are CMs (Carretero, Reference Carretero2002). In cattle, Sulzberger et al. (Reference Sulzberger, Kalebich, Melnichenko and Cardoso2016) recently showed that supplementation of CMs holds potential to improve ruminal pH and energy-corrected milk yield after receiving a grain challenge. One potential mode of action of CMs in cattle could be their potential in binding pathogenic microorganisms and toxic compounds that are associated with perturbed rumen metabolism during high-grain feeding (Saleem et al., Reference Saleem, Ametaj, Bouatra, Mandal, Zebeli, Dunn and Wishart2012), likely due to their high adsorption capacity (Slamova et al., Reference Slamova, Trckova, Vondruskova, Zraly and Pavlik2011). Indeed, in a recent study, we observed modulating effects of a CM-based product on AA-, starch- and fiber-fermenting bacteria in the ruminal microbiome during high-concentrate feeding in cows. Furthermore, some methane- and sulfate-utilizers in the epimural microbiome were affected (Neubauer et al., Reference Neubauer, Petri, Humer, Kröger, Reisinger and Zebeli2017), suggesting potentials to improve metabolic health during these feeding stress conditions. As an appropriate health status is a prerequisite for sufficient feed intake and rumination, the CM might also pose positive effects on the feeding behavior of cows experiencing SARA. Therefore, the aim of this study was to evaluate the effects of the supplementation with CM on feed intake and chewing behavior, ruminal pH and the blood metabolic profile in cows. We used a diet-induced challenge model of SARA and targeted metabolomics combined with the monitoring of health and welfare of dairy cattle by using chewing behavior, ruminal pH, liver enzymes and blood metabolites. Our working hypothesis stated that the supplementation of CM to ruminant diets could exert positive effects on rumen and systemic health under SARA-conditions, as indicated by an improved chewing behavior, systemic metabolism and liver health.
Material and methods
Animals
The trial was conducted at the research dairy farm of Vetmeduni Vienna (VetFarm Kremesberg, Pottenstein, Austria). The study was part of a broad research project called ‘Advancement of Dairying in Austria,‘ whereby other data of this project have been published previously (Kröger et al., Reference Kröger, Humer, Neubauer, Reisinger, Aditya and Zebeli2017; Neubauer et al., Reference Neubauer, Petri, Humer, Kröger, Mann, Reisinger, Wagner and Zebeli2018a; Humer et al., Reference Humer, Kröger, Neubauer, Schedle, Reisinger and Zebeli2018b). The experiment involved eight rumen-cannulated, dry, nonpregnant Holstein cows (average BW=863±65 kg, average lactation number=2.75±1.04) that were housed in a loose-housing stable.
Experimental design and feeding
Cows were subjected to a change-over design. To simulate an intermittent SARA-challenge, cows were switched two times from a pure forage diet to a high-concentrate diet (65% grains on DM-basis) during the experimental period lasting for 43 days. More specifically, cows were fed a hay and grass silage mix (50% each on DM basis) for 1 week (days 1 to 7, termed Baseline feeding). Thereafter, the concentrate level was gradually increased from days 8 to 14 up to 65%. From days 14 to 20 cows received the high-concentrate level to induce SARA (SARA 1). Afterward, the cows were fed the pure forage diet for 1 week (Recovery, days 21 to 27). Finally, after a short adaptation to the 65% concentrate diet (days 28 to 30) a 2-week SARA challenge was conducted (SARA 2, days 30 to 43). Afterward, cows were subjected to a 3-week washout phase with forage-only feeding to avoid carry-over effects.
The diets used in this study were similar to those reported previously (e.g. Kröger et al., Reference Kröger, Humer, Neubauer, Reisinger, Aditya and Zebeli2017; Neubauer et al., Reference Neubauer, Petri, Humer, Kröger, Mann, Reisinger, Wagner and Zebeli2018a). Details of diets are shown in Supplementary Table S1. The concentrates fed to cows to induce SARA contained either no additive as control (CON) or were mixed with a CM (Mycofix® Plus 3.E; BIOMIN Holding GmbH, Getzersdorf, Austria; 50 g per cow per day) and pelleted before being offered to the cows separately from forages.
The forage and concentrates were given in individual feeding troughs (Insentec B.V., Marknesse, The Netherlands) that enabled individual controlling of feed intake (Kröger et al., Reference Kröger, Humer, Neubauer, Reisinger, Aditya and Zebeli2017).
Reticular pH
The reticular pH was continuously measured using indwelling wireless pH-transmitting units (eCow Farmer bolus; eCow, Dekon, UK) as described recently (Neubauer et al., Reference Neubauer, Humer, Kröger, Braid, Wagner and Zebeli2018b). The position of the boli in the reticulum was confirmed throughout the experiment. The pH was measured every minute and summarized to 15-min intervals, which were used for calculating the daily mean and minimum pH as well as the pH <6.0 (min/day). Using the SARA-definition of Neubauer et al. (Reference Neubauer, Humer, Kröger, Braid, Wagner and Zebeli2018b) when using a reticular pH bolus, a reticular pH <6.0 for longer than 5 h/day was considered as indicative of SARA conditions.
Chewing activity
The chewing activity was determined using noseband-sensors (RumiWatch System; ITIN + Hoch GmbH, Liestal, Switzerland) as described recently by Kröger et al. (Reference Kröger, Humer, Neubauer, Reisinger, Aditya and Zebeli2017) during 2 consecutive days in Baseline (days 2, 3), SARA 1 (days 15, 16) and SARA 2 (days 35, 36). Chewing data included the duration of eating, ruminating and total chewing (min/day), the number of ruminating boli per day and the chews per bolus. Moreover, the feed intake data of the respective days were used for calculation of the chewing indices (i.e. eating, ruminating and total chewing per kilogram of dry matter intake (DMI) and kilogram of NDF intake).
Blood sampling and analysis
Blood samples were collected shortly before the morning feeding from the jugular vein on day 6 (Baseline), day 18 (SARA 1), day 28 (Recovery) and day 38 (SARA 2). Serum was obtained using 9 ml serum vacutainer tubes (Vacuette; Greiner Bio-One, Kremsmünster, Austria) and analyzed for ß-hydroxybutyrate (BHBA), non-esterified fatty acids (NEFA), cholesterol and the liver enzymes aspartate aminotransferase (AST), glutamate dehydrogenase (GLDH) and γ-glutamyltransferase (GGT). Samples for lactate and glucose analysis were collected in 6 ml fluoride plasma tubes (Vacuette). Analyses were conducted with a fully automated autoanalyzer for clinical chemistry (Cobas 6000/c501; Roche Diagnostics GmbH, Vienna, Austria).
Metabolome profiling was carried out in plasma samples collected on day 6 (Baseline) and day 18 (SARA 1) in 9 ml heparin-containing vacutainer tubes (Vacuette). A targeted metabolomics approach using electrospray ionization-liquid chromatography-MS (ESI-LC-MS/MS) was carried out by Biocrates Life Sciences AG (Innsbruck, Austria). The fully automated assay of the Absolute-IDQ p180 platform (Kit p180; Biocrates Life Sciences AG, Innsbruck, Austria) enabled the quantification of 176 metabolites out of 10 µl blood plasma. Detailed information regarding the applied technique is given elsewhere (Ramsay et al., Reference Ramsay, Stoeggl, Weinberger, Graber and Guggenbichler2012; Humer et al., Reference Humer, Kröger, Neubauer, Schedle, Reisinger and Zebeli2018b).
Statistical analyses
Statistical analysis was performed using the MIXED procedure of SAS (SAS, version 9.2). The model included the fixed effects of the treatment (i.e. CON, CM), feeding phase and their interaction. Random effects were the experimental period, cow and sequence The measurements conducted on the same cow but at different times were considered as repeated measures in the model with a first-order autoregressive variance–covariance matrix. Comparisons among treatments were evaluated by the pdiff option and degrees of freedom were estimated with the method of Kenward–Roger. The significance level was set at P≤0.05, and trends were considered at 0.05<P≤0.10 level.
The MetaboAnalyst 4.0 software (http://www.metaboanalyst.ca) was used for multivariate analysis. Partial least-squares discriminant analysis (PLS-DA) was conducted to identify characteristic trends or grouping among cows fed the Baseline v. the SARA-diets and pertaining to group CON or CM. Data were normalized by the Baseline metabolite concentrations by averaging all samples collected during the pure forage feeding. Autoscaling was conducted for compound normalization. The variables obtained on the basis of the PLS-DA results were plotted according to their importance in separating the dietary treatments based on the variable importance in the projection (VIP) scores. Moreover, hierarchical clustering analysis (HCA) with clustered heat maps using Euclidean distance measures and Ward’s clustering algorithm was carried out to explore the presence of clustering patterns among the blood metabolites affected by CM as well as reticular pH, forage and grain intake during Baseline and SARA 1. Overall correlation pattern analyses were conducted between blood metabolites as well as reticular pH, forage and grain intake.
Results
Feed intake and reticular pH
While the overall DMI during the SARA-phases did not differ among groups (Table 1), cows receiving the CM had an on average 11% increased DMI during the Recovery phase (P=0.05). Overall, similar grain intake was found among groups, while the forage intake was higher in the Recovery phase in CM-cows (P=0.01), where no concentrates were fed.
Table 1 Effect of a clay mineral (CM)-based product on feed intake and reticular pH in dairy cows subjected to two intermittent subacute ruminal acidosis (SARA)-feeding phases with 65% concentrates for 1 week (SARA 1) or 2 weeks (SARA 2), separated by a 1-week feeding with 100% forages (Recovery)

CON=control.
a,b Different superscripts represent differences among CON- and CM-groups within a feeding phase with P⩽0.05.
During SARA 1 mean reticular pH measurements revealed that cows in both groups experienced SARA (>5 h/day below pH 6.0), while this criterion was not met during the Recovery and SARA 2. Overall, no differences in the mean pH, min pH, as well as the time the pH was below the SARA-threshold were observed among the feeding groups (P⩾0.16).
Chewing behavior
In agreement with the unaffected feed intake during SARA 1 and SARA 2, the eating time did not differ between CON and CM-cows (Table 2, P⩾0.24). Similarly, the time spent ruminating per day as well as the total chewing time was not affected by the supplementation of CM. Among the eating, ruminating, and total chewing indexes related to the intake of DM or NDF, a tendency toward a 22% higher eating time per kilogram NDF intake was observed in cows receiving CM during SARA 2 (P=0.10). Furthermore, during SARA 1 these cows had a 16% higher number of ruminating chews per regurgitated bolus (P=0.01).
Table 2 Effect of a clay mineral (CM)-based product on chewing behavior in cows subjected to two intermittent subacute ruminal acidosis (SARA) challenges with 65% concentrates for 1 week (SARA 1) or 2 weeks (SARA 2)

CON=control.
a,b Different superscripts represent differences among CON- and CM-groups within a feeding phase with P⩽0.05.
y,z Different superscripts represent differences among CON- and CM-groups within a feeding phase with 0.05<P⩽0.10.
Blood parameters
The concentration of glucose, cholesterol and BHBA measured during SARA 1, Recovery and SARA 2 did not differ among groups (Table 3). In SARA 1, the cows of the CM group showed higher NEFA (P=0.03), while the lactate concentration in blood was 29% lower in the respective cows (P=0.03). Among the measured liver enzymes, AST and GLDH were reduced in the cows receiving CM in SARA 2 (−25% for AST, P=0.01 and −47% for GLDH, P=0.02).
Table 3 Effect of a clay mineral (CM)-based product on the concentration of blood metabolites in dairy cows subjected to two intermittent subacute ruminal acidosis (SARA)-feeding phases with 65% concentrates for 1 week (SARA 1) or 2 weeks (SARA 2), separated by a 1-week SARA recovery with 100% forages

CON=control; BHBA=β-hydroxybutyrate; NEFA=non-esterified fatty acids; AST=aspartate aminotransferase; GLDH=γ-glutamyltransferase; GGT=glutamate dehydrogenase.
a,b Different superscripts represent differences among CON- and CM-groups within a feeding phase with P≤0.05.
Table 4 provides a list of the blood metabolites analyzed with the targeted metabolomics approach during SARA 1 and being affected by the addition of CM, while the remaining metabolites are summarized in Supplementary Table S2. Supplementation of CM resulted in an increase in several AA up to 41% compared to CON. More specifically, glycine and the essential AA isoleucine, lysine, methionine and threonine were increased by on average 32% (P≤0.05) and arginine and histidine tended to be on average 27% higher in CM-cows compared to CON (P≤0.09). Among the BA, asymmetric dimethylarginine and dopamine were increased in CM cows (P≤0.04), while the opposite effect was found on dihydroxyphenylalanine, histamine and spermine (P≤0.01). Moreover, methionine-sulfoxide (Met-SO) tended to be reduced in CM-cows (P=0.09). Several acylcarnitines (i.e. C2, C3, C3OH, C4, C5, C5 : 1, C10 : 2, C16 : 1OH) were higher in CM-cows (P≤0.05), while the opposite was noticed for C10 (P=0.02) and C12 (P=0.08). Among PC, an effect of the CM supplementation was only found for those with diacyl-residues (i.e. PC aa C34 : 2, PC aa C36 : 4, PC aa C38 : 5, PC aa C38 : 6), showing an increase by on average 28% (P≤0.09).
Table 4 Effect of a clay mineral (CM)- based product on the concentration of blood metabolites in dairy cows subjected to a subacute ruminal acidosis (SARA)-feeding phase with 65% concentrates for 1 week

CON=control; C=carnitine; PC=phosphatidylcholine; aa=diacyl.
Only a subset of metabolites (with P≤0.10) are presented.
Multivariate analyses
The PLS-DA in Figure 1a visualizes the differences in the data among feeding phases and treatment groups. The major part of the variation in the sample set can be explained by the first principal component. Overall, the SARA samples clustered separately from the Baseline samples, whereby the cows receiving CM clustered closer to the Baseline than CON-cows. As demonstrated in Figure 1b, the most influential variables in the PLS-DA model were PCaaC364, glycine and cholesterol. The P-value for 1000 permutations was P<0.001.

Figure 1 A partial least-squares discriminant analysis (PLS-DA) of the blood metabolites that were affected by the feed additive (a). The two-dimensional score plot distinguishes the metabolic profiles of cows fed either a pure forage diet (Baseline; red) or a 65% concentrate diet (subacute ruminal acidosis, SARA) without feed additive (control, CON; blue), or a clay mineral-based product (CM; green). Variable importance in the projection (VIP) scores of 15 most influential variables for PLS-DA discriminating along principal components (b). aa=diacyl; C=carnitine; DOPA=dihydroxyphenylalanine; GGT=γ-glutamyltransferase; GLDH=glutamate dehydrogenase; Met-SO=methionine-sulfoxide; PC=phosphatidylcholine.
In addition, HCA with a clustered heat map visualization was conducted on all metabolomics variables that were affected by the CM, all classical blood metabolites as well as the feed intake and pH data to depict the relationship and differences in the concentrations among the respective variables. The HCA in Figure 2 reveals the presence of a subcluster consisting of a group of PC, Met-SO, cholesterol and forage intake. Further clusters were formed by AA and the liver enzymes, glucose and lactate. Overall, the decrease in the variables belonging to the first cluster in the SARA-diets was less pronounced in several cows receiving CM. Moreover, the increase in BA was less pronounced in CM-cows. Altogether, during the SARA-phase CON and CM-cows showed a clear separate clustering. Interestingly, two Baseline-cows even clustered within the CM-cows that received the SARA-diets.
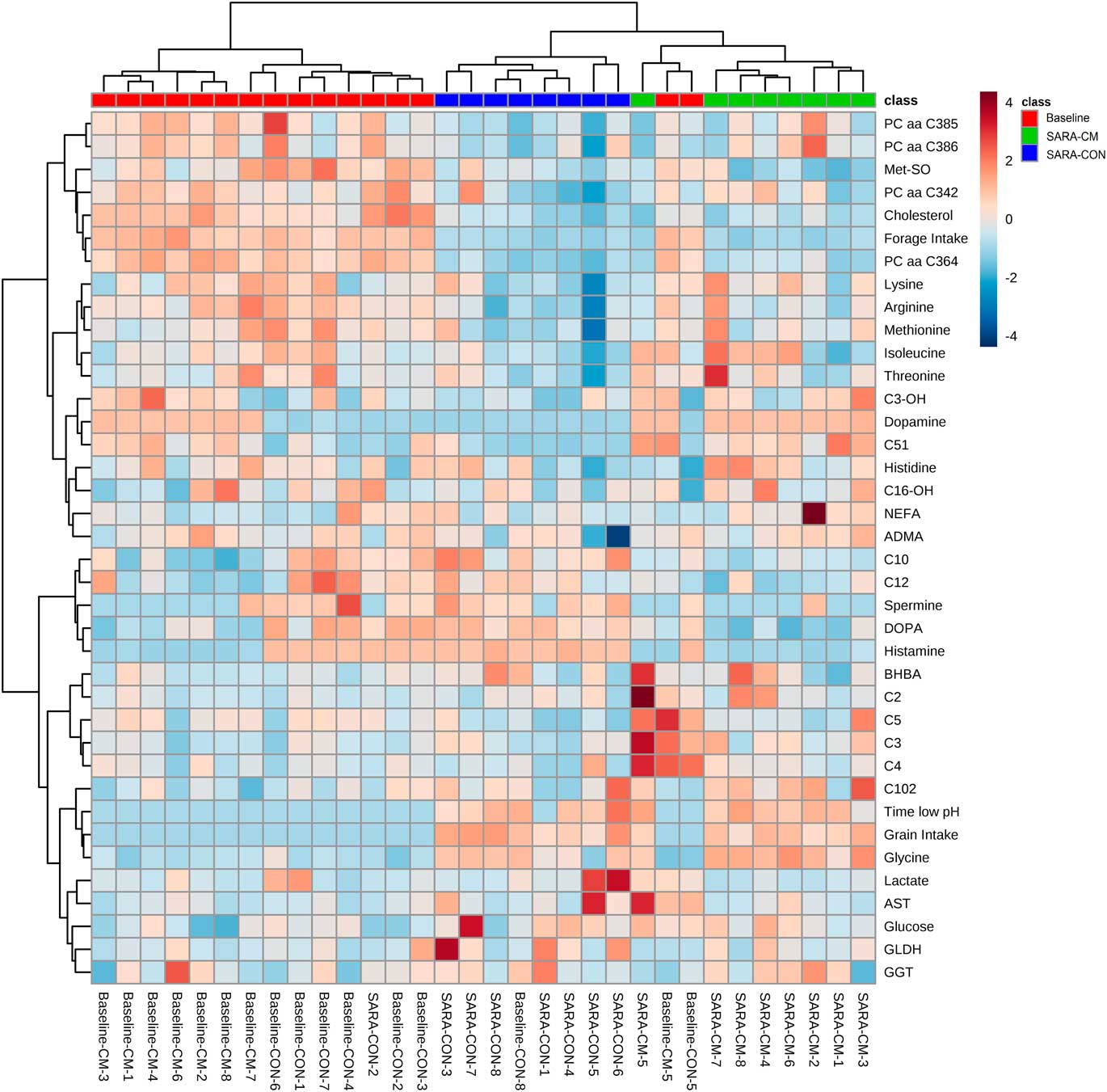
Figure 2 Hierarchical clustering analysis for blood metabolites that were affected by the feed additive, grain and forage intake, as well as reticular pH below the subacute ruminal acidosis (SARA)-threshold, measured in dairy cows receiving a pure forage diet (Baseline; red) or 65% concentrates (SARA) and receiving either no additive (control, CON; blue) or a clay mineral-based product (CM; green) during the SARA-feeding regimen. ADMA=asymmetric dimethylarginine; AST=aspartate aminotransferase; BHBA=ß-hydroxybutyrate; C=carnitine; DOPA=dihydroxyphenylalanine; GLDH=glutamate dehydrogenase; GGT=γ-glutamyltransferase; Met-SO=methionine-sulfoxide; NEFA=non-esterified fatty acids; PC=phosphatidylcholine; aa=diacyl.
Correlation pattern analysis revealed a cluster comprising of glucose, NEFA, glycine, GLDH, GGT, C10 : 2, grain intake and the time the pH was below the SARA-threshold, which showed a strong negative association with Met-SO, cholesterol, the forage intake and PC aa C36 : 4 (Figure 3). A further cluster comprising of AA (isoleucine, threonine, lysine, arginine and methionine) as well as PC aa C34 : 2 showed high positive associations with the forage intake, cholesterol and PC (i.e. PC aa C36 : 4, PC aa C38 : 5, PC aa C38 : 6) and MetSO. The latter cluster was negatively associated with a cluster of acylcarnitines (C2, C3, C4, C5), BHBA and AST.

Figure 3 Correlation of the different blood metabolites that were affected by the feed additive, as well as forage and grain intake and reticular pH below the subacute ruminal acidosis (SARA)-threshold, measured during a pure forage feeding regimen or a 65% concentrate-challenge. A heat map of Spearman correlation coefficients was generated for different concentration levels. The red color indicates a positive correlation coefficient, and the blue color represents a negative coefficient. ADMA=asymmetric dimethylarginine; AST=aspartate aminotransferase; BHBA=ß-hydroxybutyrate; C=carnitine; DOPA=dihydroxyphenylalanine; GLDH=glutamate dehydrogenase; GGT=γ-glutamyltransferase; Met-SO=methionine-sulfoxide; NEFA=non-esterified fatty acids; PC=phosphatidylcholine; aa=diacyl.
Discussion
In this research, we used a diet-induced intermittent SARA model and targeted metabolomics as well as traditional methods of monitoring health and welfare to establish the role of CM supplementation in cattle experiencing SARA. Our working hypothesis stated that the supplementation of CM to ruminant diets could exert positive effects on rumen and systemic health under such SARA-conditions. Accordingly, the study revealed that the supplementation with CM to a high-concentrate diet increased the feed intake and chewing behavior as well as altered the blood metabolomic profile in dairy cows subjected to an intermittent SARA-challenge. More specifically, the DMI was increased by 11% in cows receiving the CM during the Recovery phase, which might be indicative of an improved rumen health status due to a faster recovery from SARA, as cows experiencing SARA typically reduce their feed intake until pH returns to physiological values (Enemark, Reference Enemark2008). Moreover, during the phases in which the cows underwent the severest pH drops (SARA 1), the number of chews per bolus was enhanced from 49.9 to 57.8 in groups supplemented with CM. Overall, the number of chews per bolus is often used as a health and SARA indicator in the practice, with a common threshold of 50 chews per bolus to differentiate between healthy and diseased cows (Humer et al., Reference Humer, Aschenbach, Neubauer, Kröger, Khiaosa-ard, Baumgartner and Zebeli2018c). Therefore, the higher number of chews per bolus in cows receiving CM might be a regulatory mechanism of the cows to counteract the low ruminal pH by increased saliva secretion (Krause et al., Reference Krause, Combs and Beauchemin2002). Moreover, changes in the chewing behavior might become evident in cows experiencing SARA. In this regard, CM-cows enhanced the chewing index in terms of time spent eating per kilogram of NDF intake during SARA 2. Thus it seems that the effectiveness of forage in promoting eating chews was higher in cows receiving CM. The enhanced eating chews might have been an adaptive response to attenuate the decline in rumen pH, as previous studies have reported that chewing time per unit of NDF intake was higher for low-NDF diets compared to diets richer in NDF (Maulfair and Heinrichs, Reference Maulfair and Heinrichs2013).
Another important finding of this study was that cows receiving CM had lower concentrations of liver enzymes AST and GLDH in SARA 2. Aspartate aminotransferase and GLDH represent markers for hepatocyte integrity of dairy cows (Bobe et al., Reference Bobe, Young and Beitz2004), and the lower concentration in CM-cows likely suggests a less affected liver function in the respective cows during SARA 2. The exact mechanisms behind the beneficial effects of CM on liver health and the increase of liver enzymes following SARA are not clear. We speculate that the release of toxic compounds in the rumen and their translocation into systemic circulation (Wang et al., Reference Wang, Zhang, Zhu and Mao2013) might have played a role in the increase of liver enzymes, either directly by an increased detoxification in the liver as well as indirectly by an increased oxidative stress to the liver cells in response to toxic load (Guo et al., Reference Guo, Chang, Zhang, Xu, Jin, Bilal and Shen2017). Moreover, beneficial effects of CM supplementation on improved liver health can be explained by a lower toxic load in the rumen and systemic circulation because CM are known to adsorb toxins (Slamova et al., Reference Slamova, Trckova, Vondruskova, Zraly and Pavlik2011). Our finding of increased liver enzymes during SARA 2 indicate that effects SARA on liver enzymes are retarded compared with effects of ruminal pH which was more pronounced during SARA 1.
SARA might be associated with an increase in lactate in the rumen, that can cause absorption of lactate into the bloodstream and thus metabolic acidosis (Enemark, Reference Enemark2008). In agreement, we found increasing ruminal lactate during the SARA compared to non-SARA (Neubauer et al., Reference Neubauer, Petri, Humer, Kröger, Mann, Reisinger, Wagner and Zebeli2018a) and increased concentrations of lactate in the blood have been previously measured in cows experiencing SARA (Li et al., Reference Li, Gozho, Gakhar, Khafipour, Krause and Plaizier2012). Interestingly, lactate in the plasma was lower in CM-cows in SARA 1, which might be indicative of a lower absorption of lactate into the bloodstream and therefore might suggest improved health status of the respective cows.
In our companion study, we reported that SARA had a profound effect on the concentration of several AA, showing lowering effects in the plasma of affected cows (Humer et al., Reference Humer, Kröger, Neubauer, Schedle, Reisinger and Zebeli2018b). In addition, the feature selection based on VIP scores revealed that some AA (i.e. glycine and arginine) were instrumental for the separate clustering of the groups (Baseline, SARA-CON, SARA-CM). Interestingly, cows receiving CM showed higher concentrations of several AA during SARA than CON-cows. Besides the importance of AA for protein synthesis and carbohydrate metabolism, they are also involved in immunological processes (Suliman et al., Reference Suliman, Qureshi, Stenvinkel, Pecoits-Filho, Bárány, Heimbürger, Anderstam, Ayala, Filho, Alverstrand and Lindholm2005). As inflammatory responses trigger protein catabolism, their depletion during the SARA-phase likely derives from the release of AA from muscle proteins to serve as a substrate for acute phase proteins (Grimble, Reference Grimble2001). In this regard, it has been reported that the concentration of glycine, among others, changes differently between healthy cows and cows experiencing diseases (Hailemariam et al., Reference Hailemariam, Mandal, Saleem, Dunn, Wishart and Ametaj2014). Moreover, arginine has been reported to improve the functional activities of immune cells (Newsholme, Reference Newsholme2001). An increased synthesis of inflammatory markers was previously associated with decreasing arginine in the systemic circulation, assuming that decreased AA might be attributable to an increased synthesis of acute phase proteins (Humer et al., Reference Humer, Kröger, Neubauer, Schedle, Reisinger and Zebeli2018b). Moreover, the higher concentration of threonine might derive from an improved immune status in CM-cows, as this essential AA is the most abundant AA in immunoglobulin proteins and is also involved in maintaining the integrity of nonspecific defenses of the gut wall (Le Floc’h et al., Reference Le Floc’h, Melchior and Obled2004). It can also be speculated that a lower concentration of reactive oxygen species in the CM-cows might have contributed to the less pronounced decrease in several AA, as enhanced oxidation of AA due to the activation of pro-inflammatory cytokines might affect their concentration in the blood (Suliman et al., Reference Suliman, Qureshi, Stenvinkel, Pecoits-Filho, Bárány, Heimbürger, Anderstam, Ayala, Filho, Alverstrand and Lindholm2005).
Besides AA, also their decarboxylation products differed between CON- and CM-cows. Biogenic amines have been discussed for their deleterious effects on cow’s health when entering the systemic circulation (Wang et al., Reference Wang, Zhang, Zhu and Mao2013). In general, increasing amounts of BA are produced during high-grain feeding (Saleem et al., Reference Saleem, Ametaj, Bouatra, Mandal, Zebeli, Dunn and Wishart2012; Wang et al., Reference Wang, Zhang, Zhu and Mao2013), which can be attributed to a shift in the ruminal microbiome as well as to effects of low ruminal pH on AA-decarboxylase (Mao et al., Reference Mao, Huo and Zhu2016). More specifically, high-grain diets result in an increased accumulation of fermentation acids and promote the growth of starch- and sugar degrading bacteria such as Lactobacillus spp. and Streptococcus bovis, which produce BA (Bailey et al., Reference Bailey, Rycroft and Elliott2002). Indeed, in our companion study, the abundance of Lactobacillus increased by 100% during SARA (Neubauer et al., Reference Neubauer, Petri, Humer, Kröger, Mann, Reisinger, Wagner and Zebeli2018a). Interestingly, cows receiving CM showed changes in the ruminal microbiome compared to CON-cows (Neubauer et al., Reference Neubauer, Petri, Humer, Kröger, Reisinger and Zebeli2017), such as a lower abundance of the genus Lactobacillus in SARA 1. Moreover, the present study revealed lower concentrations of histamine, Met-SO, and spermine in the blood in the respective cows during SARA 1. This lowering effect might be either attributable to a lower production of BA in the rumen of the cows receiving CM, or a lower translocation of them into the systemic circulation. Overall, the 28% decreased concentration of plasma histamine is a positive outcome, as this BA is considered as a major toxic factor, not only in terms of acidosis but also regarding its speculated role in the pathogenesis of laminitis (Zebeli and Metzler-Zebeli, Reference Zebeli and Metzler-Zebeli2012). As Met-SO, the oxidized form of methionine has been considered to represent a biomarker for oxidative stress in vivo (Tarrago et al., Reference Tarrago, Peterfi, Lee, Michel and Gladyshev2015), the lowered concentration of Met-SO in CM-cows might derive from suspected lower oxidative protein damage (Sadri et al., Reference Sadri, von Soosten, Meyer, Kluess, Dänicke, Saremi and Sauerwein2017). Moreover, the lower concentration of spermine can be interpreted as improved health status, as spermine in the blood can be oxidized into aldehyde and hydrogen in ruminants, thereby causing oxidative stress as well as toxicological properties on eukaryotic cells (Ronchi et al., Reference Ronchi, Bernabucci, Lacetera and Nardone2000).
Overall, cows experiencing SARA showed a strong decrease in PC (Humer et al., Reference Humer, Kröger, Neubauer, Schedle, Reisinger and Zebeli2018b), which is in agreement with Saleem et al. (Reference Saleem, Ametaj, Bouatra, Mandal, Zebeli, Dunn and Wishart2012), who demonstrated reduced concentrations of ruminal PC with increasing amounts of grain in the diet. As it is assumed that most of the ruminal PC derive from protozoa, the decreasing concentrations of plasma PC likely reflect the common decrease in protozoa counts in the rumen when grain-rich diets are fed (Saleem et al., Reference Saleem, Ametaj, Bouatra, Mandal, Zebeli, Dunn and Wishart2012). Moreover, PC are used for the assembly or export of very low-density lipoprotein (VLDL) from the liver to transport the triacylglycerols (TAG). Therefore, the higher concentration of some PC in CM-cows in SARA 1 could be also related to their higher NEFA level, as NEFA taken up by the liver can be esterified into TAG and exported into to bloodstream as VLDL, as supported by the positive association between several PC and NEFA.
Furthermore, we found a counteracting effect on the SARA-associated decrease in several acylcarnitines in cows supplemented with CM. In general, acylcarnitines are involved in the transport of fatty acids from the cytoplasm across the inner mitochondrial membrane into the mitochondrial matrix (Stanley et al., Reference Stanley, Hale, Berry, Deleeuw, Boxer and Bonnefont1992). As the removal of excess of acylcarnitines from the mitochondria was hypothesized to represent a protective mechanism, it has been recently suggested that higher levels of blood acylcarnitines reflect a greater ability of hepatocytes to release any surplus of acylcarnitines from the mitochondria, thereby avoiding mitochondria damage (Huber et al., Reference Huber, Dänicke, Rehage, Sauerwein, Otto, Rolle-Kampzyk and von Bergen2016). Since properly functioning hepatic mitochondria are of vital importance for metabolic health (Kenéz et al., Reference Kenéz, Dänicke, Rolle-Kampczyk, von Bergen and Huber2016), the higher concentrations of acylcarnitines in the plasma of CM-cows may reflect their ability to adapt mitochondrial functions properly to changing metabolic situations, which concurs with the observed lowering effect on liver enzymes.
In conclusion, CM showed potential to alleviate the negative consequences of high-grain feeding in dairy cows by improving chewing behavior and liver health and counteracting multiple SARA-induced perturbations in the systemic metabolism.
Acknowledgements
The authors acknowledge the staff of the VetFarm Kremesberg (Pottenstein, Austria), S. Aditya, T. Braid, N. Kraft, P. Pourazad and A. Stauder (Institute of Animal Nutrition and Functional Plant Compounds, Vetmeduni Vienna, Austria) and M. Valera Rojas (Institute of Animal Science, Carretera Central, San José de las Lajas, Mayabeque, Cuba) for their valuable help in the experiment. Further thanks go to Königshofer Futtermittel, Königshofer GmbH (Ebergassing, Austria) for providing the concentrate. This research was funded by the project ‘ADDA – Advancement of Dairying in Austria’ of the Austrian Research Promotion Agency (FFG, Vienna, Austria) and BIOMIN Holding GmbH under the scope of the Competence Centers for Excellent Technologies (COMET) program (Grant No. 843543).
Declaration of interest
N. R. is employed by Biomin Holding GmbH, a company that manufactures and commercializes feed additives.
Ethics statement
All procedures involving animal handling and treatment were in accordance with national regulations for animal use in research and the national authority approved the study according to §26 of the Law for Animal Experiments (GZ: 68.205/0023-WF/V/3b/2015).
Software and data repository resources
Data are available from the corresponding author on reasonable request.
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.1017/S1751731118002665









 Open access
Open access