


Introduction
In the domestic rabbit, the first days after birth and the weeks around weaning are periods of major challenges to the viability of offspring. Under various breeding conditions, it was noted that the first wave of losses generally occurred between birth and day 10 (e.g. Lebas, Reference Lebas1974; Pizzi and Crimella, Reference Pizzi and Crimella1984; Gualterio et al., 1988; Seitz Reference Seitz1997; Coureaud et al., Reference Coureaud, Schaal, Coudert, Hudson, Rideaud and Orgeur2000a). The survival of individual newborns depends then on their ability to orient to the mother’s abdomen, to locate nipples and to suck efficiently in a highly constrained time span. Indeed, rabbit females nurse their litter once a day for less than 5 min (Zarrow et al., Reference Zarrow, Denenberg and Anderson1965; Gonzalez-Mariscal, Reference Gonzalez-Mariscal2007). To locate the nipples rapidly, pups rely on odour cues emitted on the surface of the females’ abdomen and/or in milk (e.g. Schley, Reference Schley1976; Hudson and Distel, Reference Hudson and Distel1983; Keil et al., Reference Keil, von Stralendorff and Hudso1990; Coureaud et al., Reference Coureaud, Schaal, Langlois and Perrier2001). These odour cues may be learned before birth (e.g. Bilkó et al., Reference Bilkó, Altbäcker and Hudson1994; Coureaud et al., Reference Coureaud, Schaal, Hudson, Orgeur and Coudert2002) or in the first postnatal hours or days (e.g. Hudson, Reference Hudson1985; Kindermann et al., Reference Kindermann, Gervais and Hudson1991; Coureaud et al., Reference Coureaud, Moncomble, Montigny, Dewas, Perrier and Schaal2006b). Some of these odour signals are however active immediately after birth, seemingly in total independence from any postnatal or prenatal experience. The mammary pheromone belongs to this latter category. It is carried in rabbit milk and is an extremely potent activator of typical orocephalic movements that pups generally display to find the nipples under the nursing female (Coureaud, Reference Coureaud2001; Coureaud et al., Reference Coureaud, Langlois, Perrier and Schaal2003 and Reference Coureaud, Langlois, Perrier and Schaal2006a; Schaal et al., Reference Schaal, Coureaud, Langlois, Giniès, Sémon and Perrier2003).
Although the bulk of rabbit pups (>90%) are highly responsive to the mammary pheromone within seconds of its presentation, a small proportion of them remains unresponsive. This observation of pheromonal unresponsiveness is even maintained under circumstances when the pups are experimentally aroused and highly motivated to suck (i.e. in the minutes preceding the daily nursing). The perceptual or motor correlates of such unresponsiveness are unknown. Yet, it may be interesting to assess whether the pups that do not react in the typical way to the mammary pheromone undergo a particular developmental trajectory, in terms of adaptation to the nursing interaction. Thus, the present study aimed to evaluate whether the responsiveness displayed by rabbit newborns to the mammary pheromone on postnatal days 1 or 3 was related with milk intake, growth and viability along most of the suckling period (i.e. between days 1 and 21).
Material and methods
Animals and housing conditions
Thirty multiparous does (strain INRA A1077; inseminated with PS19 semen, Grimaud Frères Sélection, France) and their litters were used in the breeding unit of INRA (Castanet-Tolosan, France). The does were housed in individual cages (0.28 × 0.46 × 0.69 m) to which a nest box (0.35 × 0.25 × 0.15 m) containing shavings (Copeaux Gascons, Cobois Diffusion, Lafitte sur lot, France) was appended 2 days before parturition. The animals were kept under constant light/dark cycle (16 h/8 h; lights on at 0600 h) and ambient temperature (20 to 24°C). Water and pelleted food (Rablo-Maternité; Groupe Coopératif Occitan, Castelnaudary, France) were provided ad libitum. The access to the nest was free for the mother, except on days 1 and 3 when the responsiveness to the mammary pheromone was tested (see section Behavioural assay). In total, 293 newborns from 30 litters were involved in the study. They were individually identified by ear-rings (Boucles simplex baby, Etablissement Chevillot, Albi, France) on the day of birth (day 0). On day 1, immediately after the behavioural assay and before weighing, the litters were randomly culled or completed to 10 pups (except for two litters culled to nine) to balance between litters pup competition under the mother (five pups of medium weight born to three females excluded from the protocol were fostered to two experimental females; these pups were excluded from the analyses).
Stimulus
The odour stimulus was the female rabbit’s mammary pheromone, i.e. 2-methylbut-2-enal (Sigma-Aldrich, Saint-Quentin Fallavier, France). It was diluted in distilled water at 10−6 g/ml, a concentration optimally efficient to release typical responses in 2- to 3-day-old pups (Coureaud et al., Reference Coureaud, Langlois, Sicard and Schaal2004). The stimulus was presented to the pups’ nose on a glass-stick (length × diameter: 15 × 0.3 cm), dipped in a standardised manner into the pheromonal solution.
Behavioural assay and variables
The responsiveness to the mammary pheromone was assayed on individual pups on day 1 (n = 293), then on day 3 (n = 288 pups; five pups died between days 1 and 3). The behavioural assay (e.g. Coureaud et al., Reference Coureaud, Langlois, Perrier and Schaal2003 and Reference Coureaud, Langlois, Perrier and Schaal2006a; Schaal et al., Reference Schaal, Coureaud, Langlois, Giniès, Sémon and Perrier2003) consisted in holding a pup in one gloved hand with its head left free to move. When the pup came to rest, the tip of the glass-stick carrying the mammary pheromone was positioned 0.5 cm in front of its muzzle, avoiding any contact. The duration of the odour presentation was set at 15 s.
Three levels of response were considered: (1) no response (0), meaning that the pup expressed merely sniffing or at most random head movements; (2) searching (S), that is extension of the head towards the stimulus, with low amplitude horizontal and vertical scanning movements leading to the eventual contact with the stick; and (3) grasping (G), that is searching movements terminated by one or several oral seizing of the stick. The pup responsiveness to the mammary pheromone was tested 10 min before the daily nursing (occurring, for a given litter, each day at regular time, between 0830 and 1130 h). Therefore, the nest box was closed in the evening of the day preceding the test.
Milk intake, growth and mortality
Immediately after behavioural testing, the pups of a litter were individually weighed (balance Balea P113 445; accuracy: 0.1 g; Saint Mathieu de Tréviers, France) before being put back to the nest.
Ten minutes later, the nest was opened to allow the female’s entry for suckling her litter. Almost all females readily jumped into the nest and nursed (two females on day 1 and four females on day 3 were reluctant to nurse during the time scheduled; their litters were accordingly excluded from the milk intake analyses). Five minutes after the end of the nursing episode, the pups were weighed individually to determine their milk intake. The weight was also measured after nursing on day 21. Mortality was checked daily from birth through day 21.
Regarding the does, their weight was controlled immediately after parturition, then on days 1 and 3, and their milk production was estimated on days 1 and 3 by the milk intake of their litter (measured by weighing the entire litter before/after the daily nursing; Maertens et al., Reference Maertens, Lebas and Szendrö2006). This allowed comparing the females’ lactation in the present conditions with that of females from the same strain bred in other rabbitries.
Statistical analyses
Analyses were made with the Statistical Analysis Systems Institute (1985). The rates of responsiveness to the mammary pheromone and of pup mortality were analysed using the McNemar test (for related samples) and Fisher’s exact test (for independent samples). Milk intake was examined by analysis of variance, using the general linear model. To run the analyses, pup weight was transformed in three-level variables (light, medium, heavy; referred to three subgroups of pups in Tables 1–4). The model was as follows:
where L is the litter effect (30 levels corresponding to the 30 litters); R the responsiveness to the mammary pheromone (three levels: 0, S and G); W the pup weight class (three levels: light, medium and heavy); RW the interaction between the responsiveness to the pheromone and the weight at birth; and e the residual error. When the interaction was significant, the effect of the responsiveness to the mammary pheromone was analysed for each weight class.
Table 1 Milk intake (g) on day 1 as a function of the responsiveness to the mammary pheromone (MP) and weight on day 1

a,b,A,BMeans with different superscripts differ at P < 0.05. The interaction between weight and responsiveness to the MP was significant (P < 0.05).
†(0) no response, (S) searching, (G) grasping.
For the growth performance (i.e. days 1 to 21 and days 3 to 21 weight gains), the milk intake on days 1 or 3 was added to the previous model. To that goal, the milk intake was also transformed in a three-level variable (low, medium, high; referred to three subgroups in Tables 3c and 4c).
Results
General performances of females and pups
The mean weight of the females at parturition was 3508 ± 60 g, and the mean litter size and weight at birth were 12.5 ± 0.4 and 635 ± 36 g, respectively. The proportion of pups dead at birth was 4 ± 1.6% (following the trends observed in rabbit breeding; Xiccato et al., Reference Xiccato, Trocino, Sartori and Queaque2004).
Regarding the sucking success, 14% of the pups did not get milk on day 1. This proportion decreased significantly on day 3 to reach 2% (P < 0.001). The milk intake per pup was, on average, 8.7 ± 0.3 g on day 1 and 15.7 ± 0.4 g on day 3. Regarding the females, the total milk production per doe was 85 ± 4.4 and 153 ± 5.5 g on days 1 and 3, with a high inter-individual variability (range: 36–130 g on day 1 and 68–203 g on day 3).
Along the day 0–21 period, the pup mortality was 6.8% (n ± 20/293). Most of the pups (60%) died during the first postnatal week, a common observation in rabbitries (e.g. Venge, Reference Venge1963; Lebas, Reference Lebas1974; Coureaud et al., Reference Coureaud, Schaal, Coudert, Hudson, Rideaud and Orgeur2000a; Drummond et al., Reference Drummond, Vázquez, Sanchez-Colón, Martinez-Gómez and Hudson2000). The mean weight gain of pups across the day 1–21 period, calculated by subtracting each pup individual weight on day 1 from the weight on day 21, was 275 ± 4 g (this was also in line with data from other colonies raising the same breed; Xiccato et al., Reference Xiccato, Trocino, Sartori and Queaque2004).
Immediate impact of pup responsiveness to the mammary pheromone
Among the 288 pups tested on day 1 and then on day 3, 8.7% and 7.2%, respectively, did not respond to the mammary pheromone. This rate of non-response was stable between days 1 and 3 (χ 2 < 0.5, P > 0.05). The remaining pups responded strongly to the mammary pheromone by searching (S), and more often by grasping (G) (more than 70% on days 1 and 3; comparisons between S and G pups: χ2 > 137, P< 0.001 on day 1 as on day 3). The rate of pups that only searched decreased from days 1 to 3 (21.2% to 3.1%; χ 2 = 43.3, P< 0.001) while the pups that grasped increased correlatively (70.1% to 89.6%; χ 2 = 49.1, P< 0.001; Figure 1). Taken together, the pups responding to the pheromone (by S or G) were as frequent on day 1 than on day 3 (91.3% v. 92.8%; χ 2 = 0.5, P > 0.05).
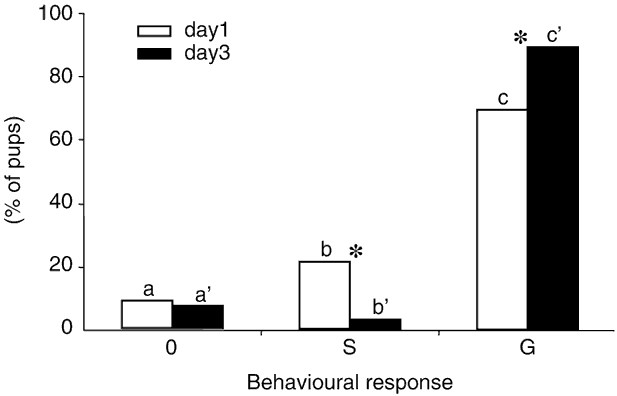
Figure 1 Frequency (%) of rabbit newborns displaying only sniffing (0), sniffing and searching (S), or sniffing, searching and grasping (G) to the mammary pheromone on days 1 and 3 (n = 288 pups, from 30 litters, tested on both days). a, b, c and * indicate significant differences (P < 0.05) among types of responses on a given day, or between days for a given response, respectively.
The milk intake on day 1 was affected by an interactive effect of the responsiveness to the mammary pheromone and the weight of the pups (P < 0.05). In light pups (<48 g), the sucking success was low for those who were unresponsive, as compared with the responding pups displaying either searching or grasping (comparisons between 0, S and G pups: P < 0.01). In contrast, the milk intake of the medium and heavy pups (≥48 g) was similar whatever the responsiveness to the mammary pheromone (Table 1). The weight by itself was also influential, at least in unresponsive pups or in pups responding by grasping. In these subgroups, the lightest pups were less successful in gaining milk than the medium and heaviest pups (difference in milk gain between light v. medium, and light v. heavy pups: −97.2% and −98.1% for unresponsive pups, and −22% and −33% for G pups; P < 0.05 in all cases; Table 1).
The results were clearly different on day 3, however. The interaction effect of the reactivity to the mammary pheromone and the pre-nursing weight on milk intake did no more reach significance. However, milk intake was still influenced by pup weight, at least in G pups (i.e. 89.2% of the pups). Thus, lighter pups gained less milk than medium pups, which themselves were less efficient than heavier pups (r = 0.49; P < 0.001; Table 2).
Table 2 Milk intake (g) on day 3 as a function of the responsiveness to the mammary pheromone (MP) and weight on day 3

A,B,CMeans with different superscripts differ at P < 0.05. The interaction between weight and responsiveness to the MP was not significant (P = 0.53).
†(0) no response, (S) searching, (G) grasping.
Table 3 Growth and mortality of pups between days 1 and 21 according to the responsiveness to the mammary pheromone (MP), weight and milk intake on day 1

a,b,cMeans with different superscripts differ at P < 0.05. Regarding the weight gain, the interactions between the pup weight, the responsiveness to the MP and the milk intake on day 1 and on day 3 were not significant.
†(0) no response, (S) searching, (G) grasping.
Table 4 Growth and mortality of pups between days 3 and 21 according to the responsiveness to the mammary pheromone (MP), weight and milk intake on day 3

a,b,cMeans with different superscripts differ at P < 0.05. Regarding the weight gain, the interactions between the pup weight, the responsiveness to the MP and the milk intake on day 1 and on day 3 were not significant.
†(0) no response, (S) searching, (G) grasping.
To sum up, pup responsiveness to the mammary pheromone was linked to milk intake on day 1, but no more on day 3. This pheromone-milk intake link on day 1 is effective in the lightest pups, leading to significantly lower milk intake in unresponsive pups.
Delayed impact of the early responsiveness to the mammary pheromone
(1) Pup survival during the day 1–21 period. Over the first postnatal week, pups who were unresponsive on day 1 had a higher rate of mortality (14.3%) than those who searched or grasped (0% and 3.9% for S and G pups, respectively; Table 3a). Moreover, but regardless of the response to the mammary pheromone, pup nursing weight and milk intake on day 1 were related to pup mortality: more losses were recorded for light, as opposed to medium/heavy pups (Table 3b), and for those pups who took less than 7 g of milk on day 1 (Table 3c). It may be noted that the four pups that died among the unresponsive group were lighter than the surviving pups in this group (36.0 ± 9 v. 59.2 ± 2 g, P < 0.001). The same trend was seen in the pups who responded by oral grasping to the mammary pheromone (dying v. surviving pups: 34.2 ± 5 v. 51.9 ± 1 g, respectively; P < 0.001).
After day 7, the responsiveness to the mammary pheromone initially displayed on day 1 did no more appear to impact on pup survival. However, the mortality during the day 8–21 period, and more generally during the day 1–21 period, was influenced by the weight and milk intake of the pups on day 1: the pups that died were the lightest and the most unsuccessful in gaining milk (Table 3b and c).
On day 3, the link between pup pheromonal response and survival seemed to fade (for days 3 to 7, 8 to 21 and 3 to 21; Table 4a). However, the pup weight and milk intake on day 3 remained associated with survival on the following days, in the sense that the lightest and least successful suckers survived less than the other pups (Table 4b and c).
(2) Pup growth during the day 1–21 period. The mean weight (before nursing) of the pups was 52 ± 0.7 g on day 1, 70 ± 1 g on day 3 and 328 ± 4.1 g on day 21. Under our conditions, no link appeared between the day-1 response to the mammary pheromone and the weight gain of the pups until day 21 (Table 3a). However, the weight and milk intake had a strong impact, a low weight or a weak sucking success on day 1 affecting significantly the growth across the day 1–21 period (r = 0.60, P < 0.001 and r = 0.25, P < 0.001, respectively; Table 3b and c).
Similarly, pup response to the mammary pheromone on day 3 did not appear to be correlated with later growth (Table 4a). In contrast, the weight gain during the day 3–21 period was influenced both by the weight and by the milk intake of the pups on day 3 (r = 0.52, P < 0.001 and r = 0.45, P < 0.001, respectively; Table 4b and c).
Discussion
The present study aimed to evaluate whether a link could be evidenced between the response of rabbit pups to the pure mammary pheromone and the rate of survival and growth in breeding conditions. It was based on the hypothesis that the unresponsiveness to the pheromone on the first postnatal day should predict non-response to this signal in the nursing context.
In the present conditions, about 90% of the tested pups responded to the mammary pheromone on days 1 or 3. This high level of response corroborates previous observations on the strong releasing potency of this compound (Coureaud et al., Reference Coureaud, Langlois, Perrier and Schaal2003, Reference Coureaud, Langlois, Sicard and Schaal2004 and Reference Coureaud, Langlois, Perrier and Schaal2006a; Schaal et al., Reference Schaal, Coureaud, Langlois, Giniès, Sémon and Perrier2003; Montigny et al., Reference Montigny, Coureaud and Schaal2006). In the same time, the behavioural assay using the mammary pheromone permitted to target pups that are unresponsive to the pheromone on days 1 and 3. The follow-up of these non-responsive pups allowed gauging their differential developmental trajectory in terms of milk intake, growth and viability.
A superficial look on the longitudinal results would suggest that unresponsive pups are those who will die between birth and day 21 (6.8%). However, it comes out that the birth weight and milk intake have buffering effects in pups who do not respond to the mammary pheromone. The pups that died were not only those who failed to respond to the mammary pheromone on day 1 (80% responded). Unresponsive pups on day 1 were indeed not all ineffective in gaining milk (only 18.5% did not get milk on day 1), nor were they all dying out (85.7% survived). In fact, the impact of a non-response to the mammary pheromone rests on the pup weight (only the lightest unresponsive pups display difficulties to suck and survive) and on the period when pheromonal reactivity is assessed (the impact of non-response on day 1 is limited to the day 1 milk intake and the days 1 to 7 survival). Factors other than pheromonal perception certainly influence early survival, among which pup maturity at birth, females’ parity and behavioural treatment of the nest and litter are the most noteworthy (Coudert, Reference Coudert1982; Verga et al., Reference Verga, Canali, Pizzi and Crimella1986; Fortun-Lamothe, Reference Fortun-Lamothe1998; Canali et al., Reference Canali, Ferrante, Todeschini, Verga and Carenzi1991; Hamilton et al., Reference Hamilton, Lukefahr and McNitt1997; Coureaud et al., Reference Coureaud, Schaal, Coudert, Hudson, Rideaud and Orgeur2000a). Lastly, it must be stated that the present behavioural assay is rather punctual, being run once for 15 s, and that it may therefore not be ubiquitously sensitive for the varied risk factors affecting viability.
However, it appears that for the lightest pups, responding to the mammary pheromone on day 1 bears a clear advantage to gain milk. Those pups are indeed as successful in sucking as their heavier, pheromone-unresponsive or responsive littermates. Thus, for the lightest pups, the day 1 pheromonal responsiveness, weight and milk intake are additive in improving viability. The impact of the two latter factors was already known (McNitt and Moody, Reference McNitt and Moody1987; Coureaud et al., Reference Coureaud, Schaal, Coudert, Rideaud, Fortun-Lamothe, Hudson and Orgeur2000b; Poigner et al., Reference Poigner, Szendro, Lévai, Radnai and Biro-Németh2000; Perrier et al., Reference Perrier, Jouanno and Drouet2003). But here, we add the perceptual ability of day-1 pups as an additional predictor, which is especially valid for the thinnest pups during the first postnatal week.
The causes of such individual differences in the early behavioural impact of the mammary pheromone are unclear. Pheromonal unresponsiveness of pups may first relate to uneven maturation among littermates in terms of perceptual, integrative or motor performances. It may also be due to birth-related factors, such as delayed recuperation from the stress of delivery or from obstructed respiration. Anyway, defective response to the pure mammary pheromone on day 1 may reflect general difficulty to respond to odour cues of the nursing female, increasing the risk to miss the first nursing sessions and to rapidly die (failing to suck on two to three consecutive nursing bouts is generally lethal; e.g. Venge, Reference Venge1963; McNitt and Moody, Reference McNitt and Moody1987; Coureaud et al., Reference Coureaud, Schaal, Coudert, Rideaud, Fortun-Lamothe, Hudson and Orgeur2000b). Here, 100% of the pups who were pheromone-unresponsive on day 1 and who died during the experimental period, died effectively before day 7. However, the majority of pups who were unresponsive to the pure pheromone administered once on day 1 became responsive on day 3 and survived (n = 22/27), suggesting that a functional catch up is possible. Such recuperative outcome has then most chance to turn into an adaptive end in the nursing competition (e.g. Drummond et al., Reference Drummond, Vázquez, Sanchez-Colón, Martinez-Gómez and Hudson2000; Bautista et al., Reference Bautista, Mendoza-Degante, Coureaud, Martina-Gomez and Hudson2005). Moreover, after the day-1 test and during the next sucking bouts (days 2 to 3), some individuals could find the nipples and gain milk in tracing circumstantial maternal odorants that they have learned as directional cues (e.g. Hudson, Reference Hudson1985; Coureaud et al., Reference Coureaud, Moncomble, Montigny, Dewas, Perrier and Schaal2006b).
To sum up, the responsiveness of newborn rabbits to the mammary pheromone appears to be an indicator for individual viability, at least for the lightest pups and when their responsiveness is checked on postnatal day 1. The use of a pheromone to probe viability in rabbitries remains now to be ascertained. Pheromone testing on day 1 may permit selection among the lightest pups those who can support the competition for milk with heavier newborns. Conversely, pheromone-response defective pups on day 1 being at higher mortality risk, they might be reunited with light pups to be artificially fed or fostered under one female, allowing alleviation of competition and thus improved welfare and survival.
Acknowledgements
The authors acknowledge the important technical assistance and generous help of B. Lamboley-Gaüzère, C. Bannelier, J. Dedapper, A. Lapanouse and P. Aymard.





