Introduction
Drought stress, caused by water shortage has become a main ecological problem with global climate anomalies. It limits growth and physiological mechanism in crops by reducing leaf water potential, respiration and photosynthesis (Khandal et al., Reference Khandal, Parween, Roy, Meena and Chattopadhyay2017). Many crops, including cool-season grasses, are particularly susceptible to drought stress (West, Reference West, Bacon and White2018). Drought causes yield and quality losses by reducing photosynthesis following physiological production processes such as reactive oxygen species and lipid peroxidation in cool-season grasses such as Lolium perenne L. (perennial ryegrass) (Vanani et al., Reference Vanani, Shabani, Sabzalian, Dehghanian and Winner2020).
Perennial ryegrass, a cool-season forage and turfgrass, is used in temperate regions of the world. However, its future productivity will be limited by drought due to its rather shallow rooting depth (Bothe et al., Reference Bothe, Westermeier, Wosnitza, Willner, Schum, Dehmer and Hartmann2018). Despite this, researches on drought stress have been limited compared to cereals and legumes in perennial ryegrass (Vanani et al., Reference Vanani, Shabani, Sabzalian, Dehghanian and Winner2020). Therefore, there is a need to understand perennial ryegrass responses to water shortage at both physiological and gene expression levels and develop tolerant perennial ryegrass varieties against drought stress. Perennial ryegrass plants have generally evolved severe strategies to maintain growth and development such as the synthesis of phytohormones as a result of the expression of drought-related genes (bZIP, MYB, DREB, WRKY) (Martin et al., Reference Martin, Kronmiller and Dombrowski2021; Yang et al., Reference Yang, Lu, Wang, Wang, Liu and Chen2021).
The examination of genetic diversity is a crucial aspect of successful drought stress-tolerant crop improvement programmes, as it enables breeders to select plants with the greatest potential for enhancement (Donde et al., Reference Donde, Kumar, Gouda, Gupta, Mukherjee, Baksh, Mahadani, Sahoo, Behera and Dash2019). Collecting populations from regions with high genetic diversity such as Turkey is the most natural way to find and develop plants having the potential to tolerate stress factors such as drought and salinity (Demirkol and Yılmaz, Reference Demirkol and Yılmaz2019). Studies have shown that genetic diversity is important for improving drought tolerance in perennial grass populations, and high levels of diversity can increase the success of breeding projects focused on improving drought stress tolerance (Turner et al., Reference Turner, Holloway-Phillips, Rawnsley, Donaghy and Pembleton2012; Bothe et al., Reference Bothe, Westermeier, Wosnitza, Willner, Schum, Dehmer and Hartmann2018). Molecular markers are the most effective and reliable tool for assessing genetic diversity. The utilization of Simple Sequence Repeat (SSR) markers has been widely recommended by researchers for assessing genetic diversity in various crops, including perennial ryegrass due to their high information content within and between populations (Diekmann et al., Reference Diekmann, Hodkinson and Barth2012; Guan et al., Reference Guan, Yuyama, Stewart, Ding, Xu, Kiyoshi and Cai2017).
The aim of this study was to investigate the effects of drought stress on both the physiological and molecular characteristics of genetically diverse populations of perennial ryegrass.
Materials and methods
Collected populations
The populations were collected from the natural flora of the Ordu province in Turkey (online Supplementary Table S1).
SSR primers
The molecular markers (online Supplementary Table S2) were selected from a previous study that showed high polymorphism in perennial ryegrass plants (Jones et al., Reference Jones, Dupal, Kölliker, Drayton and Forster2001). These markers have also been used in drought and submergence stress studies in perennial ryegrass (Wang, Reference Wang2009; Yu et al., Reference Yu, Bai, Luo, Chen, Liu, Liu, Warnke and Jiang2011).
Growth and stress conditions
The seeds of the populations were sterilized with 70% (v/v) ethanol solution by shaking for 1 min and then washing with ddH2O. The experiment was performed in a growth chamber. The conditions were under a 16 h light/8 h dark photoperiod at 25/18°C and with 60% relative humidity. 50 ml of deionized water was included in the pots daily. Pots contained a 3:1 mixture of perlite sand soil. The treatments were normal (non-stress) or drought stress conditions for 30-day-old seedlings. Each treatment was represented by two pots each with four plants. Hoagland's nutrient solution was used for the non-stressed group as a control, while PEG 6000 (Merck, Germany) (15%, w/v) was used for drought-stressed group in the study designed with four replications. Samples were cut in 21th day after drought stress treatment. The schema illustrating the procedural framework is presented in Fig. 1.

Figure 1. Schematic representation of the procedural framework in perennial ryegrass plants.
Growth parameters
Root and shoot samples of the harvested plants were cut. The lengths (cm) of these samples were measured. Then the samples were kept in an oven at 60°C for 72 h. Afterward, dry weights (g/plant) of the samples at constant weight were determined.
Relative water content
The relative water content of the stressed and non-stressed plants was evaluated by the method of Farrant (Reference Farrant2000), by calculating the following formulation; 100 × (fresh weight – dry weight)/(turgid weight – dry weight).
Determination of free proline content
The free proline content of the drought-stressed and non-stressed plants was evaluated by the method developed by Bates et al. (Reference Bates, Waldren and Teare1973). The samples weighing 0.3 g, were homogenized in 10 ml of 3% sulfosalicylic acid. The supernatant was obtained through centrifugation at 10 000 g for 10 min. Subsequently, 1 ml of the extract was transferred to a 5 ml glass tube, and 1 ml of glacial acetic acid, along with 1 ml of ninhydrin reagent, was added to it. The samples were covered with aluminium foil and incubated in a water bath at 95°C for 30 min. The reaction was then stopped by cooling the samples in an ice bath for 5 min. Following this, the cooled samples were vortexed with 4 ml of toluene and analysed using a spectrophotometer at 520 nm.
Determination of total chlorophyll content
The procedure for determining the total chlorophyll content in leaf samples involved the homogenization of 1.0 g of the leaves in 90% acetone, followed by filtration of the resultant extract. The absorbance values of the filtered extract were measured using a UV/Vis spectrophotometer (Shimadzu, Japan) at wavelengths of 645 nm and 663 nm, in accordance with the method proposed by Arnon (Reference Arnon1949).
Gene expression analysis
The plant total RNAs were extracted with the plant RNA kit (Qiagen), and the cDNAs were synthesized using the iScriptTM cDNA Synthesis Kit (Qiagen).
The qRT-PCR program was: 30 s at 94°C (predenaturation), 10 s at 95°C (denaturation), 15 s, 58.8°C for bZIP and MYB, 59.2°C for DREB, and 59.2°C for WRKY (annealings), 10 s, 72°C, 40 cycles (extension), 15 min, 72°C (final extension). The information about primer were shown in online Supplementary Table S3.
Genomic DNA isolations and SSR analysis
For genomic DNA isolation, samples taken from the young leaves of 10 randomly selected plants from each population were performed according to the CTAB method developed by Rogers and Bendich (Reference Rogers and Bendich1985).
PCR amplifications were conducted with 5X C Taq Master Mix (Promega Corporation, USA) in total volumes of 25 μl. The Bio-Rad Thermal Cycler was used to handle the PCR products. The program was: 5 min, 95°C (initial denaturation), of 30 s, 95°C, 35 cycles (denaturation), 30 s, 58.4–60.4°C (annealing) and 30 s at 72°C.
PCR products were analysed under UV light in 2.5% agarose gel stained with ethidium bromide and viewed using a gel imager (Bio-Rad).
Statistical analyses
Statistical analyses were performed with Minitab software (Minitab Inc., State College, PA) in the study. One-way ANOVA was used for the analyses of data and multiple comparisons were performed by using Tukey test. The genetic diversity of the populations was evaluated utilizing the NTSYS software version 2.1.
Results
Growth parameters of the populations
The drought stress significantly increased root dry weights and root lengths (average of 26 and 19% in root dry weights and root lengths, respectively) of all studied populations except for P3, P7 and P29 (Fig. 2(a) and (c)). Shoot dry weights and shoot lengths were not changed in P6, P10, P15 and P30 between control and drought treatments, while significant decreases were observed in the other populations under drought stress (average of 35 and 38% in shoot dry weights and shoot lengths, respectively) (Fig. 2(b) and (d)).

Figure 2. Growth parameters of the perennial ryegrass populations under normal and drought-stressed conditions. a. Root dry weight (g/plant), b. Shoot dry weight (g/plant), c. Root length (cm), d. Shoot length (cm). The columns represent means and the bars represent standard errors. * shows a significant differentiation among drought and control treatment in a population by P < 0.05, while ** means a similar difference by P < 0.01.
Biochemical status of the populations
P6, P10, P15 and P30 significantly maintained their relative water content under drought stress, compared to control, while dramatic decreases were observed in the other populations under drought stress (Fig. 3(a)). Free proline contents of the thirteen populations increased under drought stress, compared to control (Fig. 3(b)). The populations of P6, P10, P15 and P30 showed higher free proline contents under drought stress, compared to non-stress condition (7.8, 9.6, 8.3 and 11 folds, respectively) (Fig. 3(b)). Total chlorophyll contents were significantly decreased under drought stress in the thirty-three populations (average of 39%), while no changes were observed in P10, P15 and P30 (Fig. 3(c)).

Figure 3. Biochemical status of the perennial ryegrass populations under normal and drought-stressed conditions. a. Relative water content (%), b. Free proline content (μmol/g), c. Total chlorophyll content (mg/g FW). The columns represent means and the bars represent standard errors. * shows a significant differentiation among drought and control treatment in a population by P < 0.05, while ** means a similar difference by P < 0.01.
The drought-related gene expression levels
The relative expression level of bZIP11 exhibited an average of 5.2-fold up-regulation under drought stress in P6, P10, P15, P21, P22 and P30, while an average of 2.5-fold up-regulation was determined in bZIP37 in P6, P10, P15, P21, P25 and P30. (Fig. 4(a) and (b)). No change was detected in the expression level of bZIP11 and bZIP37 in the other populations (Fig. 4(a) and (b)).

Figure 4. The relative expression levels of the transcription factors of the drought-related genes of the perennial ryegrass populations under normal and drought-stressed conditions. a. bZIP11 (folds), b. bZIP37 (folds), c. DREB3 (folds), d. DREB5 (folds), e. MYB48 (folds), f. MYB112 (folds), g. WRKY56 (folds), h. WRKY108715 (folds). The columns represent means and the bars represent standard errors. * shows a significant differentiation among drought and control treatment in a population by P < 0.05, while ** means a similar difference by P < 0.01.
The expression levels of DREB3 and DREB5 were found to be significantly up-regulated under drought stress in six specific populations (P6, P10, P15, P18, P25 and P30) (Fig. 4(c) and (d)). However, no changes in expression were observed in the remaining populations when compared to the control group (Fig. 4(c) and (d)).
The relative expression level of MYB48 was nearly 10-fold up-regulated in all three populations in response to drought stress in P6, P10 and P30, while MYB112 was up-regulated in P6, P10, P15 and P30 (Fig. 4(e) and (f)). No change was observed in the expression levels of MYB48 and MYB112 in the other populations (Fig. 4(e) and (f)).
The drought stress up-regulated the expression levels of WRKY56 in P6, P10, P15, P21 and P30, while similar up-regulations were observed in WRKY108715 in P6, P10, P15, P21, P22 and P30 (Fig. 4(g) and (h)H). Especially, P30 showed higher increases under drought stress (7.1 and 6.2 folds in WRKY56 and WRKY108715, respectively). No change was detected in the transcription factors of WRKY56 and WRKY108715 in the other populations (Fig. 4(g) and (h)).
SSR analyses
Totally, 38 alleles were observed using ten SSR primers (online Supplementary Table S4). The average polymorphic information content (PIC) value was determined as 0.74 (online Supplementary Table S4). Two SSR primers (P-07 and P-08) had four unique alleles in P6, P10 and P15 (data not shown).
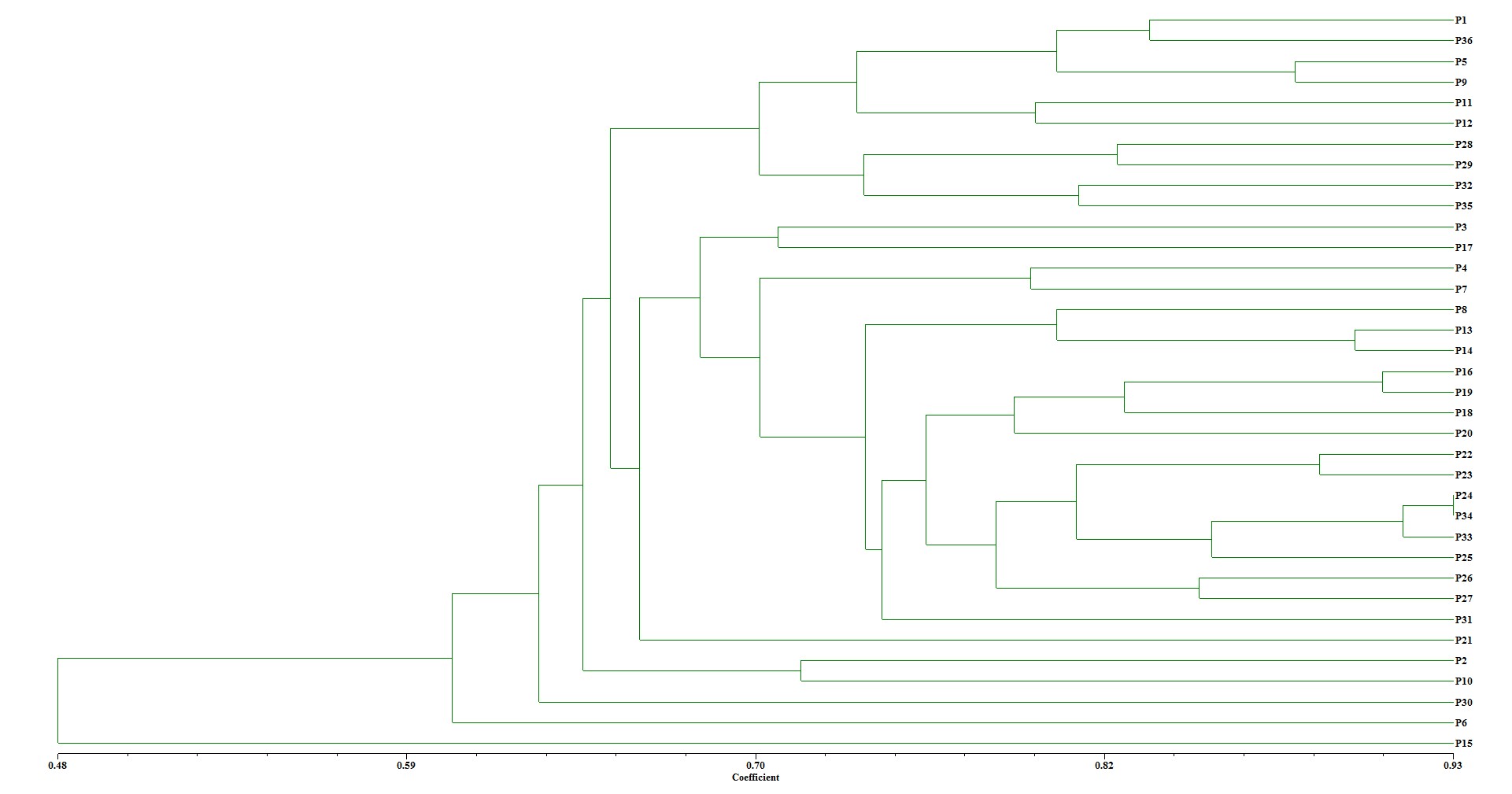
A dendrogram was formed between the populations using the UPGMA method (online Supplementary Fig. S1). The perennial ryegrass populations were divided into four groups based on the dendrogram, which revealed high genetic differences between the populations (online Supplementary Fig. S1). P6, P15 and P30 were classified into unique groups (online Supplementary Fig. S1). In addition to UPGMA clustering, similar outcomes were observed in the principal coordinate analyses (PCA). The P6, P10, P15 and P30 were located further apart from other genotypes, by PCA (Fig. 5).

Figure 5. The PCA analysis of the perennial ryegrass populations.
Discussion
Increased root development is an essential defence response in plants to cope with drought stress (Bothe et al., Reference Bothe, Westermeier, Wosnitza, Willner, Schum, Dehmer and Hartmann2018; Demirkol, Reference Demirkol2021a). In a study, a similar increase was reported in root development in perennial ryegrass under drought stress (Foito et al., Reference Foito, Byrne, Shepherd, Stewart and Barth2009). In the study, P6, P10, P15 and P30 maintained their biomass and relative water content under drought stress condition (Figs 2(b), (d) and 3(a)), suggesting that these populations were able to preserve more water during the drought condition. The accumulation of free proline in P6, P10, P15 and P30 served to protect against osmotic stress and maintained cellular turgor pressure, thereby preserved the structural integrity of the cell membrane. Similar free proline content increases were reported in perennial ryegrass and red fescue plants in response to drought (Bandurska and Jozwiak, Reference Bandurska and Jozwiak2010). Total chlorophyll contents of P10, P15 and P30 were not changed under drought stress (Fig. 3(c)), suggesting that these populations prevented the breakdown of chlorophylls by minimizing available water loss in the face of drought stress. These results collectively demonstrate that P6, P10, P15 and P30 showed better growth characteristics and physiological status compared to other populations under drought stress. The physiological characteristics observed in P6, P10, P15 and P30, including increased root development (root dry weight and root length), maintained biomass (shoot dry weight and shoot length), relative water content, total chlorophyll content and increased free proline content under drought-induced stress have crucial potential for the enhancing drought tolerance breeding strategies in perennial ryegrass plants.
Researchers have indicated that bZIP, DREB and WRKY family member genes have crucial roles to cope with abiotic stress factors by regulating ABA signalling pathway (Liu et al., Reference Liu, Wu and Wang2012; Wang et al., Reference Wang, Wang, Yao, Zhou, Li and Jiang2014; Yan et al., Reference Yan, Jia, Chen, Hao, An and Guo2014; Dossa et al., Reference Dossa, Wei, Li, Fonceka, Zhang, Wang, Yu, Boshou, Diouf, Cisse and Zhang2016; Dong et al., Reference Dong, Ma, Wisniewski and Cheng2017; Gao et al., Reference Gao, Wang, Xu and Zhang2018; Wang et al., Reference Wang, Zhu, Li, Wang and Wu2018; Demirkol, Reference Demirkol2021b). These genes have been found to be induced by dehydration and triggered in ABA-dependent ways (Kudo et al., Reference Kudo, Kidokoro, Yoshida, Mizoi, Todaka, Fernie, Shinozaki and Yamaguchi-Shinozaki2017; Zhang et al., Reference Zhang, Li, Hassan, Li and Peng2020). The results of this study suggest the possible regulatory role of up-regulated bZIP11, bZIP37, DREB3, DREB5, WRKY56 and WRKY108715 in the response to drought stress in the drought-tolerant perennial ryegrass plants P6, P10, P15 and P30 (Fig. 4(a), (b), (c), (d), (g) and (h)). Researchers have showed that MYB genes enable the plant to cope with drought stress by stimulating the biosynthesis of proline, ABA and gibberellic acid in the plant (Zhang et al., Reference Zhang, Zhao, Xia, Jia, Liu and Kong2012; Baldoni et al., Reference Baldoni, Genga and Cominelli2015; Chen et al., Reference Chen, Li, Hu, Guo, Liu and Zhang2015). The results of this study showed that the up-regulated MYB48 and MYB112 may be effective genes for improving the drought tolerance of the drought-tolerant perennial ryegrass plants (Fig. 4(e) and (f)). ABA is known to up-regulate free proline accumulation by promoting the expression of genes involved in proline synthesis (Kishor and Sreenivasulu, Reference Kishor and Sreenivasulu2014). In this study, the enhanced free proline accumulation and maintained phenotypic characteristics under drought conditions in drought-tolerant populations, confirm the function of ABA under drought stress.
In the molecular characterization study, two SSR primers (P-07 and P-08) identified a total of four unique alleles out of 38 alleles in P6, P10 and P15. Unique alleles have great potential to be used for desired diagnostic studies that allow the targeted plants to be easily distinguished from other genotypes (Kumbhar et al., Reference Kumbhar, Kulwal, Patil, Sarawate, Gaikwad and Jadhav2015; Donde et al., Reference Donde, Kumar, Gouda, Gupta, Mukherjee, Baksh, Mahadani, Sahoo, Behera and Dash2019). Surprisingly, these unique alleles were detected in drought-tolerant populations in this study. These primers have been reported to be related with physiological drought tolerance of perennial ryegrass plants (Wang, Reference Wang2009; Yu et al., Reference Yu, Bai, Luo, Chen, Liu, Liu, Warnke and Jiang2011). In the dendrogram, P6, P15 and P30 were classified into unique groups (online Supplementary Fig. S1). Similarly, P6, P10, P15 and P30 (drought-tolerant populations) were located further apart from other genotypes, by PCA (Fig. 5). This means that the drought-tolerant perennial ryegrass populations are genetically different from other populations. Similar separation as drought tolerant and others was reported in perennial ryegrass plants (Jonavičienė et al., Reference Jonavičienė, Statkevičiūtė, Kemešytė and Brazauskas2014). Taken together, supporting the physiological data with molecular analysis makes these drought-tolerant perennial ryegrass populations promising genetic resources.
Conclusion
The results indicate that P6, P10, P15 and P30 populations seem to have great potential for their resistance to drought stress possibly by activating drought-related genes. These populations could be utilized to develop perennial ryegrass cultivars that can mitigate drought stress conditions.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S1479262123000795
Acknowledgement
The present study was funded Ordu University Scientific Research Projects Coordination Unit, as A-1834.