Introduction
Bambara groundnut (Vigna subterranea (L.) Verdc) is an underutilized and research neglected leguminous crop (Massawe et al., Reference Massawe, Mwale, Azam-Ali, Roberts, Ochatt and Jain2007) and in Malawi, it is grown in most areas especially by rural women. It is regarded as a complete food as it contains sufficient quantities of protein (18–20%), carbohydrate (60–65%) and fat (Goli, Reference Goli, Begemann and Mushonga1995; National Research Council, 2006). Therefore, Bambara groundnut is one of the legumes with the potential of improving nutrition and addressing malnutrition levels reportedly at 47% among the children under the age of five in Malawi.
The world holds large collections of Bambara germplasm characterized by wide genetic diversity (GD). Several studies of GD analysis in Bambara groundnut have been conducted using both agro-morphological traits (Karikari and Tabona, Reference Karikari and Tabona2004; Ntundu et al., Reference Ntundu, Shillah, Marandu and Christiansen2006; Ouedraogo et al., Reference Ouedraogo, Ouedraogo, Tignere, Bilma, Dabire and Konate2008; Abu and Buah, Reference Abu and Buah2011; Olukolu et al., Reference Olukolu, Mayes, Stadler, Ng, Fawole, Dominique, Azam-Ali, Abbott and Kole2012; Aliyu and Massawe, Reference Aliyu and Massawe2013; Molosiwa et al., Reference Molosiwa, Aliyu, Stadler, Mayes, Massawe, Kilian and Mayes2015; Olanrewaju et al., Reference Olanrewaju, Oyatomi, Babalola and Abberton2021) and molecular markers (Olukolu et al., Reference Olukolu, Mayes, Stadler, Ng, Fawole, Dominique, Azam-Ali, Abbott and Kole2012; Aliyu and Massawe, Reference Aliyu and Massawe2013; Mayes et al., Reference Mayes, Basu, Molosiwa, Redjek, Ahmad, Khan, Zehra, Noah, Mayes, Roberts, Stadler, Massawe, Kilian and Azam-Ali2013; Molosiwa et al., Reference Molosiwa, Aliyu, Stadler, Mayes, Massawe, Kilian and Mayes2015). The molecular markers that have been used in GD analysis in Bambara include; RAPD (Amadou et al., Reference Amadou, Bebeli and Kaltsikes2001; Massawe et al., Reference Massawe, Roberts and Davey2003; Rungnoi et al., Reference Rungnoi, Suwanprasert, Somta and Srinives2012), AFLP (Massawe et al., Reference Massawe, Dickinson, Roberts and Azam-Ali2002; Ntundu et al., Reference Ntundu, Bach, Christiansen and Andersen2004), SSR (Somta et al., Reference Somta, Chankaew, Rungnoi and Srinives2011; Olukolu et al., Reference Olukolu, Mayes, Stadler, Ng, Fawole, Dominique, Azam-Ali, Abbott and Kole2012; Aliyu and Massawe, Reference Aliyu and Massawe2013; Molosiwa et al., Reference Molosiwa, Aliyu, Stadler, Mayes, Massawe, Kilian and Mayes2015), DArT Array markers (Stadler, Reference Stadler2009; Olukolu et al., Reference Olukolu, Mayes, Stadler, Ng, Fawole, Dominique, Azam-Ali, Abbott and Kole2012; Molosiwa et al., Reference Molosiwa, Aliyu, Stadler, Mayes, Massawe, Kilian and Mayes2015; Ahmad et al., Reference Ahmad, Chai, Basu, Sri Redjeki, Moreton, Mayes, Ho, Massawe and Mayes2016).
GD analysis helps to align the breeding objectives with identified diversity more especially during selection of parental materials to initiate a breeding programme. No comprehensive agro-morphological or molecular marker GD analysis has been reported in Malawi on Bambara groundnuts except for one genotype that was reported by Molosiwa et al. (Reference Molosiwa, Aliyu, Stadler, Mayes, Massawe, Kilian and Mayes2015). This suggests that Bambara groundnut has been grossly neglected in Malawi in terms of research and variety selection. To date, Malawi lacks an active Bambara groundnut breeding programme. As such there is a dearth of improved varieties for farmers, resulting in the cultivation of low yielding landraces with an average yield of less than 400 kg/ha among smallholder farmers (Pungulani et al., Reference Pungulani, Kadyampakeni, Nsapato and Kachapila2012). However, this is lower than the Africa average of 0.85 t/ha. Yields well over 3 t/ha have been reported in Africa (Azam-Ali et al., Reference Azam-Ali, Sesay, Karikari, Massawe, Aguilar-Manjarrez, Bannayan and Hampson2001; Hillocks et al., Reference Hillocks, Bennett and Mponda2012; Berchie et al., Reference Berchie, Dapaah, Agyeman, Sarkodie-Addo, Addo, Addy and Blankson2016; Mubaiwa et al., Reference Mubaiwa, Fogliano, Chidewe, Jan Bakker and Linnemann2018; Tan et al., Reference Tan, Azam-Ali, Goh, Mustafa, Chai, Ho, Mayes, Mabhaudhi, Azam-Ali and Massawe2020). This suggests there is potential to develop high yielding Bambara groundnuts for improved food and nutrition security.
With renewed interest in setting up a Bambara groundnut improvement programme for Malawi, there was need for a thorough understanding of the GD present in the existing germplasm using both morphological characterization and genetic markers. Therefore, this research was formulated to characterize Bambara genotypes using agro morphological traits and DartSeqLd SNP markers. Further, to identify and select high yielding Bambara groundnuts lines that can be released as new varieties to the farming communities or used as parents in a Bambara groundnut breeding programme.
Materials and methods
Study site description
The study was conducted at Bunda College of Agriculture Research farm in the 2019/2020 and 2021/2022 seasons. The site receives a mean annual rainfall of about 930 mm with a temperature range of 17.2°C to 19.1°C.
Plant materials
A diverse panel of 56 Bambara groundnut germplasm was assembled. This included materials collected locally from farmers and markets in Malawi as well as materials from different countries sourced through the IITA Gene bank Nigeria.
Field trials experimental designs and data collection
The field agronomic trials were carried out under rain-fed conditions in 2019/2020 and 2021/2022 seasons. For the 2019/2020 trials, 56 genotypes were planted using a 7 by 8 alpha lattice design in which there were 7 blocks of size 8 and replicated three times. The plot constituted of 2 ridges each measuring 1.5 m with an intra-row spacing of 15 cm and 0.75 m between ridges. One seed was planted per planting station and the trial was planted on 30 November 2019. For the 2021/2022 trial, 60 genotypes were planted on 3rd January, 2022 using a 5 by 12 alpha lattice design. The trial had 3 replications with 12 blocks of size 5. The plot had 2 ridges spaced at 0.75 m, each 2.1 m long. Data were collected following the procedure described in the Bambara groundnut descriptor (Ipgri/Iita/Bamnet, 2000).
Genotyping sample preparation, processing and analysis
For genotyping purposes, six plants for each of the 60 genotypes were tagged in preparation for leaf sampling. The six plants within the genotypes were sampled to detect the within genotype variation to facilitate single plant selection.
Leaf tissue sampling for DNA analysis
Three weeks after germination, a young leaf from each of the six tagged plants from each genotyped was sampled for DNA analysis. The sampling was done according to the required procedure by Intertek-Agritech laboratory (Intertek-Agritech, 2016). First, unique sample identifications were generated using breeding management system, which were used to track the samples throughout the genotyping process. A single hole-puncher (6.0 mm diameter) was used to punch and collect four leaf discs per sample from young and healthy newly developed leaf. The punched leaf discs from each sample were transferred with forceps directly into a single well of a 96-well plate (1.2 ml AbGene Storage Plate, number AB0564, Thermo Fisher Scientific). Wells G12 and H12 of the sampling plates were left blank as control wells according to Intertek-Agritech lab standard protocol. The forceps were cleaned with 75% alcohol before and after placing each sample in a well to avoid cross contamination. The sample plates were later oven dried for 24 h at 48 degrees Celsius. The 96-well sample plates were sealed with sealing mats (AB0674, Thermo Fisher Scientific), wrapped in plastic bags, secured firmly and shipped for DNA extraction to Intertek-Agritech laboratory in Sweden.
DNA isolation and genotyping
After the extraction of High Molecular Weight DNA from the tissue samples and after quality check, DNA was sent for high-throughput DArTseqLD SNP genotyping at the Diversity Arrays Technology Pty Ltd. Canberra, Australia. The DNA samples were subjected to complexity reduction by digestion/ligation reactions as described by Kilian et al. (Reference Kilian, Wenzl, Huttner, Carling, Xia, Blois, Caig, Heller-Uszynska, Jaccoud, Hopper, Aschenbrenner-Kilian, Evers, Peng, Cayla, Hok and Uszynski2012) but replacing the single PstI-compatible adaptor with two adaptors corresponding to PstI and MseI restriction enzymes’ overhangs. The PstI-compatible adaptors were designed to include the Illumina flowcell attachment sequence, barcode region, while the reverse adaptor contained a flowcell attachment region and MseI-compatible overhang sequence. Only mixed fragments (PstI-MseI) were effectively amplified in PCR. Subsequently, equimolar amounts of PCR product from each sample were bulked and sequenced by the Hiseq2500/ Novaseq6000 (Illumina® Inc., San Diego, CA, USA). Thereafter, the generated sequences were processed using proprietary DArT analytical pipelines. Approximately 250,000 sequences per barcode/sample were used in marker call. Identical sequences were collapsed into FASTQCOL files followed by SNPs calling using the software package DArTsoft14.
Data analysis
Agro-morphological data
An exploratory analysis using density plots was done on quantitative data to check for ANOVA assumptions which included Shapiro–Wilk test for normality and Levine's test for equal variance. Thereafter, all analyses were done in the R software environment (R Core Team, 2021). ANOVA was done by fitting the model with the lmer function of the lme4 package (Bates et al., Reference Bates, Mächler, Bolker and Walker2015) in R and P values were adjusted using Tukey method to compare means at 0.05 significance level. Principal component analysis (PCA) was done using the FactoMineR package (Lê et al., Reference Lê, Josse and Husson2008) and Pearson correlation was performed using the corr. function of the stats package in R. A hierarchical cluster analysis using Euclidean distance metric and the associations between groups were done using the ward D2 method in a cluster R package (Maechler et al., Reference Maechler, Rousseeuw, Struyf, Hubert and Hornik2019).
Analysis of Bambara groundnut genotyping data
Genotyping data were filtered using a minor allele frequency (MAF) threshold of 0.05 which eliminated 660 SNPs. Diversity metrics which included MAF, Neis GD, polymorphic information content and inbreeding coefficient (Fi) were estimated using the snpReady package (Granato et al., Reference Granato, Galli, de Oliveira Couto, e Souza, Mendonça and Fritsche-Neto2018). Principal coordinate analysis was also conducted using dartR package in R (Gruber et al., Reference Gruber, Unmack, Berry and Georges2018) to determine the contributions of each component to the variation that existed in the germplasm. AMOVA was performed to quantify genetic variation among individuals, among populations and within individuals using Genalex ver. 6.502 (Peakall and Smouse, Reference Peakall and Smouse2012), with statistical significance based on 999 permutations. Bayesian information criterion was used to estimate the optimum number of clusters (K), followed by examination of population structure by discriminant analysis of principal components (DAPC) using the adegenet package in R (Jombart et al., Reference Jombart, Kamvar, Collins, Lustrik, Beugin, Knaus and Jombart2020).
Results
Genotypic variation in Bambara groundnuts
Tables 1 and 2 show the mean square values for the different traits scored in 2019/2020 and 2021/2022 seasons, respectively. For the 2019/2020 season, there were significant genotype effects for grain yield (P < 0.001), pod length (P < 0.001), pod width (P < 0.001), number of pods per plant (P < 0.001) and number of seeds per pod (P < 0.001). For the 2021/2022 season, a similar trend was observed where significant differences were observed for grain yield (P < 0.01), 100 seed weight (P < 0.05), seed length (P < 0.001), number of pods per plant (P < 0.001), pod width (P < 0.001), pod length (P < 0.001), plant height (P < 0.001), canopy spread (P < 0.001), number of stems per plant (P < 0.001), number of branches per stem (P < 0.001), number of nodes per stem (P < 0.001), internode length (P < 0.001), petiole length (P < 0.001), terminal leaflet width (P < 0.001), terminal leaflet length (P < 0.001), number of leaves per plant and peduncle length (P < 0.001). However, significant genotypic effects, were not observed for number of seeds per pod (P > 0.05), number of days to 50% flowering (P > 0.05) and number of days from sowing to first flowering (P > 0.05).
Table 1. Mean square values for traits scored in 2019/2020 season

SOV, Source of variation; DF, Degrees of freedom; GYLD, Grain yield (kg/ha); PODL, Pod length; PODW, Pod width. ***highly significant at P < 0.001, **highly significant at P < 0.01, *Significant at P < 0.05. P values adjusted using Tukey method at 0.95 Confidence level.
Table 2. Mean square values for traits scored in 2021/2022 season

GYLD, Grain yield (kg/ha); S100W, Weight of 100 seeds; SDL, Seed length; NSP, Number of seeds per pod; NPP, Number of pods per plant; PDW, Pod width; PDL, Pod length; D50F, Number of days from first flowering to 50% flowering; PHT, Plant height; CNS, Canopy spread; DFF, Number of days from sowing to first flowering; NSTP, Number of stems per plant; NBS, Number of branches per stem; NNS, Number of nodes per stem; INL, Internode length; PTL, Petiole length; TLW, Terminal leaflet width; TLL, Terminal leaflet length; NLP, Number of leaves per plant; PEDL, Peduncle length. ***highly significant at P < 0.001, **highly significant at P < 0.01, *Significant at P < 0.05. P values adjusted using Tukey method at 0.95 Confidence level.
Performance of Bambara groundnuts for yield and related traits
The results on the mean performance of Bambara genotypes for the 2019/2020 growing season are presented in Table S1. Grain yield ranged from 252 kg/ha (TVSu-1831) to 1410 kg/ha (TVSu-1321). For pod length, values ranged from 15.4 mm to 21.3 mm for TVSu-1831 and TVSu-368, respectively. In terms of pod width, genotypes TVSu-1448 and TVSu-1733 had the highest pod width of 11.3 mm and 11.0 mm, respectively and genotypes TVSu-1258 (7.6 mm) and TVSu-1831 (6.7 mm) recorded lowest pod width. On average, the Bambara genotypes produced 18.4 pods per plant. For the 2021/2022 growing season, (Tables S2 and S3), grain yield ranged from 104 kg/ha (TVSu-250) to 1828 kg/ha (Mazira-a-mpheta). In terms of weight of 100 seeds, the highest weights were obtained from genotypes TVSu-1557 (90.1 g) followed by BC13 (85.1 g). The lowest 100 seed weight was recorded on genotype TVSu-1888 (46.5 g).
Principal component and correlation analysis for agro-morphological traits
PCAs grouped all the agronomic traits scored in 2021/2022 season under five components (Eigen values ≥1) which accounted for 71.8% of the variation (Table S4). All the traits exhibited a negative association with PC1 which accounted for 32.2% of the total variation. Number of branches per stem, number of stems per plant and number of leaves per plant were correlated with PC2 and PC3, while grain yield, pod length, pod width and peduncle length were correlated with PC4.
The correlation analysis exhibited several significant correlations among the traits scored (Fig. 1). The distribution of each trait is shown on the diagonal while on the bottom of the diagonal, the bivariate scatter plots with a fitted line are displayed. On the top of the diagonal, the correlation coefficients and the significance level (P-values) are indicated. Grain yield (kg/ha) was significantly correlated with 100 seed weight (r = 0.32, P < 0.05), number of pods per plant (r = 0.53, P < 0.001), plant height (r = 0.41, P < 0.01), canopy spread (r = 0.58, P< 0.001), petiole length (r = 0.41, P< 0.01), number of leaves per plant (r = 0.44, P< 0.001), terminal leaflet width (r = 0.35, P< 0.01) and peduncle length (r = 0.29, P< 0.05). Apart from grain yield, several other traits were also highly correlated showing a positive association between the traits.

Figure 1. A scatter plot matrix showing the correlation coefficients among traits and the significance levels scored in 2021/2022 season.
Hierarchical clustering of genotypes based on agro-morphological traits
Cluster analysis grouped the genotypes into five clusters based on the agro-morphological traits (Fig. 2). Cluster V had the highest number of genotypes (17) and lowest number was observed in cluster I (8) (Table S5). In terms of origin of the genotypes, clusters II and IV were dominated with genotypes from Malawi while cluster I was dominated by genotypes from Cameroon and cluster III was dominated by genotypes from Burkina Faso and Malawi. Cluster V had a unique distribution in which genotypes origin varied remarkably but it was dominated by genotypes from West Africa. Clusters I and II were dominated by high yielding genotypes, high number of pods per plant, and most of these originated from Malawi. Clustering analysis showed genotypes from the same geographical areas were generally grouped together. A similar trend was observed with clustering based on SNP markers.

Figure 2. Hierarchical clustering dendrogram analysis; Euclidean distance was used and the associations between groups were done by the Ward method for the morphological traits.
Molecular diversity and population structure
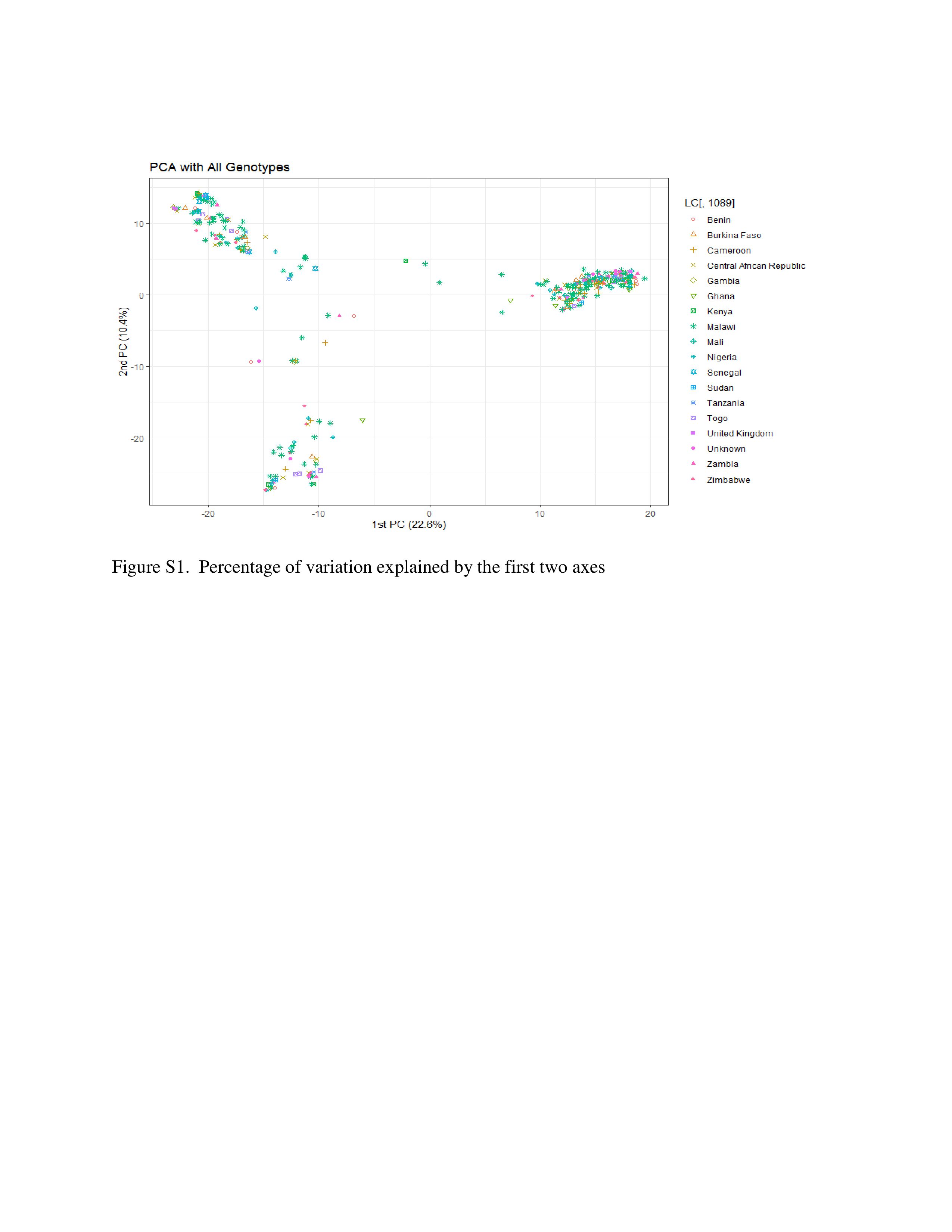
The average Nei genetic distances (Table 3) among the genotypes within each population revealed that the genotypes in population 1 (mostly West and some East African genotypes) were closely related to population 3 (Southern African genotypes) which exhibited the smallest genetic distance value (0.030). Though both population 1 and 2 were dominated by West African genotypes, they exhibited the greatest genetic distance (0.137). Similarly, population 3 (Southern African genotypes) were distantly (0.115) related to population 2 (West African genotypes). The populations 3 and 1 were closely related probably due to the presence of East African genotypes (Sudan and Kenya) which geographically are in close proximity to the Southern African region, or they might have shared many common alleles and are closely related to each other.
Table 3. Pairwise Nei genetic distances among three subpopulations

Pop 1, Cluster 1 = > West African genotypes (including some East African Genotypes from Kenya & Sudan); Pop 2, Cluster 2 = > West African genotypes (Tanzania is the outlier, and UK); Pop 3, Cluster 3 = > Mostly Southern African genotypes.
The AMOVA results (Table 4) showed that the total genetic variance found among populations was 10% while among the individuals contributed to 90% of the genetic variance and no variation was detected within the individuals. The high percentage value of GD within-population (among individuals) obtained from AMOVA could be due to natural adaptation.
Table 4. Analysis of molecular variance (AMOVA) for the three geographical regions of the Bambara groundnut Genotypes

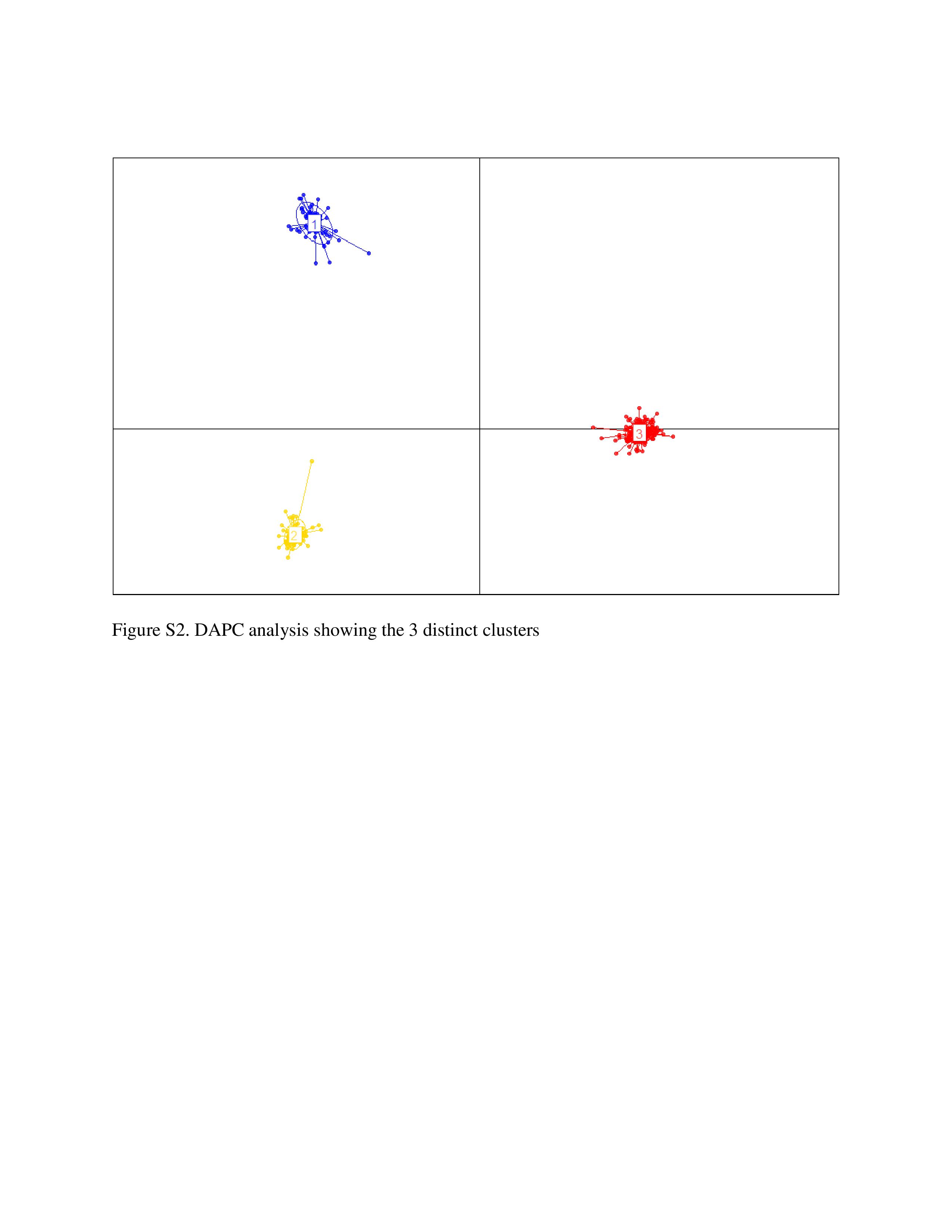
The cumulative percentage contributions of the first two principal components (PCs) to the total variations in the populations were 33%, with 22.6% and 10.4% respective contributions to PC1 and PC2 (Figure S1). The membership clustering of DAPC revealed three major groups of genotypes (Figure S2). DArTseqLD showed that 358 samples derived from 60 genotypes of Bambara groundnut were grouped into three clusters (Figure S3). Cluster I had 60 samples, cluster II had 110 samples and cluster III had 188 samples.
Discussion
The significant variations observed in the genotypes for a number of traits suggest that genetic variability exists in Bambara groundnuts which could be exploited in breeding programmes aimed at developing high performing varieties. Significant genotypic variability has extensively been reported (Ntundu et al., Reference Ntundu, Shillah, Marandu and Christiansen2006; Vurayai et al., Reference Vurayai, Emongor and Moseki2011; Shegro et al., Reference Shegro, Jansen Van Rensburg and Adebola2013; Mohammed, Reference Mohammed2014; Aliyu et al., Reference Aliyu, Massawe and Mayes2016; Gbaguidi et al., Reference Gbaguidi, Dansi, Dossou-Aminon, Gbemavo, Orobiyi, Sanoussi and Yedomonhan2018; Valombola et al., Reference Valombola, Akundabweni, Awala and Hove2019; Olanrewaju et al., Reference Olanrewaju, Oyatomi, Babalola and Abberton2021; Onwubiko, Reference Onwubiko2021) on a number of traits in Bambara groundnut including grain yield, 100 seed weight, pod length, pod width and many others. The existence of this genetic variability signifies that Bambara groundnut could be responsive to selection and that superior genotypes could be developed despite reported difficulties in making crosses (Basu et al., Reference Basu, Roberts, Azam-Ali, Mayes and Kole2007) due to strict self-pollination in the crop.
The average grain yield reported here is 714 kg/ha which is above the national average that smallholder farmers realize in their farms in Malawi (Pungulani et al., Reference Pungulani, Kadyampakeni, Nsapato and Kachapila2012). The differential performance could be attributed to different management conditions under farmers’ fields and heterogeneity of the farm lands. This can also be attributed to better genetic potential of the genotypes used in this study than landraces used by farmers which tend to be inherently low yielding compounded by poor management practices. Other studies have also reported grain yield values within this margin, for example, Akpalu et al. (Reference Akpalu, Sarkodie-Addo and Akpalu2012) reported Bambara groundnut grain yield in the average range of 650 kg/ha–850 kg/ha. The results on grain yield from this study are consistent with what others have reported as yield ranges in Bambara groundnuts (Akpalu et al., Reference Akpalu, Sarkodie-Addo and Akpalu2012; Pungulani et al., Reference Pungulani, Kadyampakeni, Nsapato and Kachapila2012; Mohammed, Reference Mohammed2014; Gbaguidi et al., Reference Gbaguidi, Dansi, Dossou-Aminon, Gbemavo, Orobiyi, Sanoussi and Yedomonhan2018; Valombola et al., Reference Valombola, Akundabweni, Awala and Hove2019; Kunene, Reference Kunene2021; Olanrewaju et al., Reference Olanrewaju, Oyatomi, Babalola and Abberton2021).
The 100 seed weight is a useful indicator for yield and this study has also shown some significant positive correlation between the two traits. Others have used 100 seed weight in evaluating morphological features such as seed size (Massawe et al., Reference Massawe, Mwale and Robert2005; Mohammed, Reference Mohammed2014; Unigwe et al., Reference Unigwe, Gerrano, Adebola, Pillay and Monrovia2016) signifying its role in determining grain yield. The positive association between grain yield and other traits suggested that indirect selection for yield trait can be achieved to an extent through selection based on other traits especially in the early stages of selection where multi location trials may not be possible. This also suggests that breeding for any one of these correlated traits will not reduce the desired level of the other and therefore, selection of high yielding genotypes could be based on the other traits such as 100 seed weight and number of pods per plant. These findings auger well with other studies (Ofori, Reference Ofori1996; Aliyu et al., Reference Aliyu, Massawe and Mayes2016; Gbaguidi et al., Reference Gbaguidi, Dansi, Dossou-Aminon, Gbemavo, Orobiyi, Sanoussi and Yedomonhan2018; Valombola et al., Reference Valombola, Akundabweni, Awala and Hove2019; Olanrewaju et al., Reference Olanrewaju, Oyatomi, Babalola and Abberton2021) which had also reported significant correlations in Bambara groundnuts for different traits.
GD is an important enabler for crop improvement. The current study observed higher GD within population (90%) than among populations, suggesting substancial gene flow among the landraces from the different origins, across large geographical distances. Though some literature have reported higher genetic variation among populations than within population, this study is consistent with what Minnaar-Ontong et al. (Reference Minnaar-Ontong, Gerrano and Labuschagne2021) and Uba et al. (Reference Uba, Oselebe, Tesfaye and Abtew2021) observed in their respective studies.
Genotypes in the same cluster reflect genetic relationships and most diverse accessions were out grouped. The understanding of GD within Bambara genotypes will enhance future planning in breeding programmes and provide helpful information in maintaining and monitoring GD required for a robust breeding programme (Eltaher et al., Reference Eltaher, Sallam, Belamkar, Emara, Nower, Salem K, Poland and Baenziger2018).
Clustering analysis showed that some of the Bambara groundnut genotypes from the same region did not cluster accurately based on their origin. According to Uba et al. (Reference Uba, Oselebe, Tesfaye and Abtew2021) this could be due to low genetic differentiation among the populations, which suggests that the genetic background of Bambara groundnut populations does not always correlate with their geographical area of origin. However, it was worth noting that majority of the genotypes from the same origin clustered together which provides evidence in the origin and relationship of Bambara groundnut genotypes. Similar patterns of grouping of landraces according to geographical origin have been reported using morphological markers (Ntundu et al., Reference Ntundu, Shillah, Marandu and Christiansen2006; Aliyu and Massawe, Reference Aliyu and Massawe2013) and molecular markers (Jaccoud et al., Reference Jaccoud, Peng, Feinstein and Kilian2001; Rungnoi et al., Reference Rungnoi, Suwanprasert, Somta and Srinives2012; Odongo et al., Reference Odongo, Oyoo, Wasike, Owuche, Karanja and Korir2015; Uba et al., Reference Uba, Oselebe, Tesfaye and Abtew2021) for collections of Bambara groundnut landraces within countries and among regions. The clustering of some genotypes from the different regions into the same cluster might suggest some relatedness between accessions from different regions which could partly be attributed to seed exchange as previously alluded to by Uba et al. (Reference Uba, Oselebe, Tesfaye and Abtew2021). Population structure analysis of this study suggested that the genotypes with unknown origin and United Kingdom genotypes might have originated from Western Africa region, because they clustered consistently with the subpopulation consisting of genotypes from West Africa.
Other studies (Somta et al., Reference Somta, Chankaew, Rungnoi and Srinives2011; Rungnoi et al., Reference Rungnoi, Suwanprasert, Somta and Srinives2012; Uba et al., Reference Uba, Oselebe, Tesfaye and Abtew2021) had reported similar trend where they observed that genotypes from locations close to each other were more related and had small genetic distances between them. High distant genetic relatedness reported here has important implications in parental selection for the genetic improvement of Bambara. If hybridization is made between distant related genotypes, it may result in high genetic recombination in the progenies that might be useful in enhancing the genetic gain from selection in the crop.
Conclusion
The significant differences observed on various morphological traits among Bambara groundnut genotypes such as grain yield and weight of 100 seeds confirm that there is genetic variation within and between Bambara groundnut genotypes. While some Bambara groundnut genotypes can be identified and selected and grown directly for their high yields, some may be identified, selected and exploited for utilization in Bambara groundnut breeding programmes. DArT SNP marker molecular characterization of Bambara groundnut genotypes from different regions revealed that variation exists among the genotypes. This level of GD could be the basis for developing new Bambara cultivars with desirable characteristics such as high yield while being adapted to diverse environments. The study identified three subpopulations which could be explained by their geographical differentiation, natural selection and regional adaptation history. Based on grain yield, the study identified the following genotypes as top performers across the evaluation seasons TVSu-1321, TVSu-1557, TVSu-1256, TVSu-1733, Mazira-a-mpheta, BC7, Mazira-a-mpheta-1, TVSu1794, TVSu-1448, TVSu-793, TVSu-388, TVSu-1312, TVSu-1797, BC13, TVSu-542, Kayera, BC3 and TVSu-1045. These will need to be further tested under farmers’ field conditions to confirm the high yielding ability and most preferred ones and later on released for production by farmers.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S1479262123000503.
Acknowledgements
Research assistants at Bunda College of Agriculture provided technical support for the trials. International Foundation for Science provided the financial support for the study.
Author contributions
Michael Chipeta conceptualized, designed and implemented the research, collected data, analysed phenotyping data and wrote the first draft of the manuscript. Davis Gimode analysed molecular data and revised the draft. Both authors read and approved the final manuscript.
Financial support
This research was supported by the International Foundation for Science, Stockholm, Sweden, through a grant number 1-1-c-6190-1 to Michael Chipeta.
Competing interest
The authors have no competing interests to declare that are relevant to the content of this article.