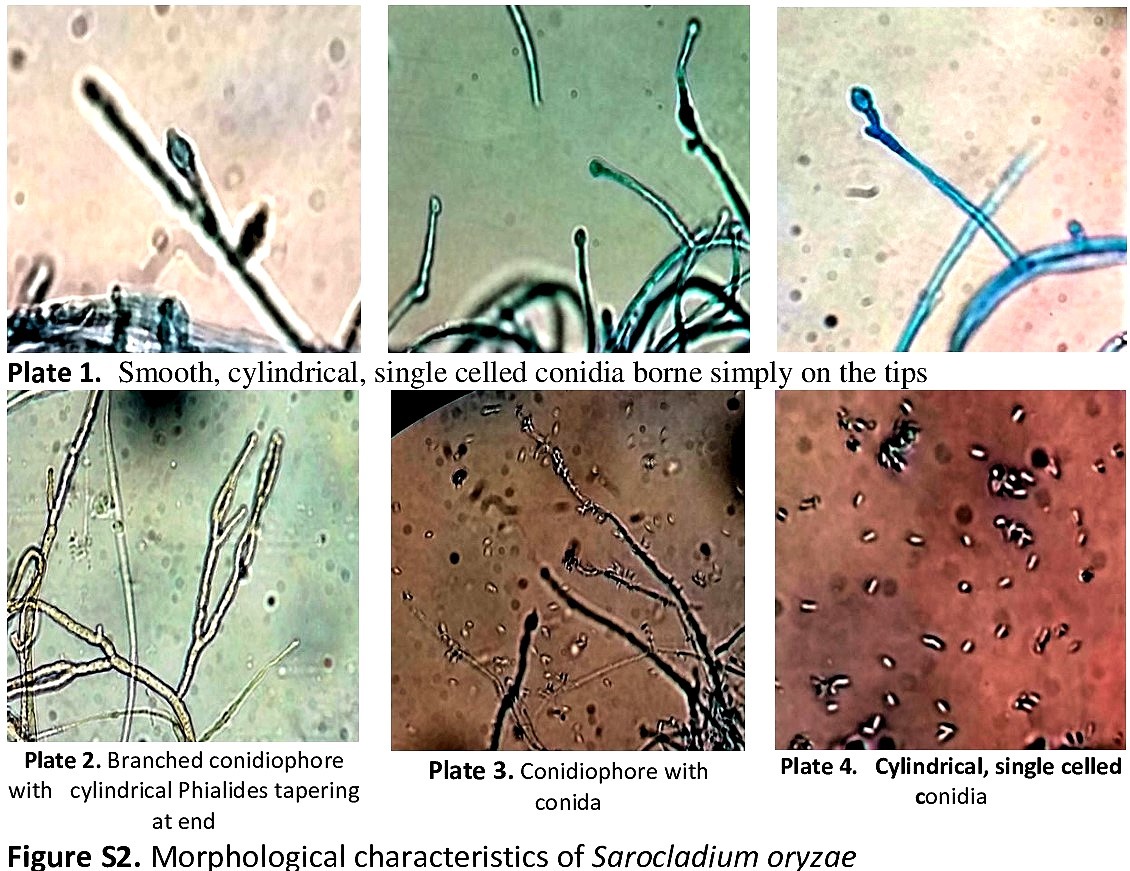
Introduction
Rice (Oryza sativa L.) is the main staple food for more than half of the world population and is being cultivated in 114 countries over an area of 161.2 million hectares. The annual production of rice in the world is 480.01 million tonnes, out of which 90% is being cultivated in Asia (FAO, 2021). Rice is the primary food grain crop in Jammu and Kashmir, India cultivated under irrigated conditions and is frequently challenged by various biotic and abiotic stresses (Teeli et al., Reference Teeli, Bhat, Hussain, Ahangar and Ganaie2018). The impact of diseases on rice production and productivity has increased over time. In current rice cropping system of Kashmir significant yield losses in rice was observed due to a few major diseases such as rice blast (70–80%) sheath blight (20–60%) and 3–95% by bakanae disease (Singh and Sunder, Reference Singh and Sunder2012; Jamaloddin et al., Reference Jamaloddin, Durga Rani, Swathi, Anuradha, VaniSri and Rajan2020; Molla et al., Reference Molla, Karmakar and Molla2020). However, sheath rot incited by Sarocladium oryzae ([Sawada] W. Games and D. Hawksworth) is the most important emerging disease, particularly in Japonica rice causing 20–85% yield loss (Mvuyekure et al., Reference Mvuyekure, Sibiya, Derera, Nzungize and Nkima2018; Peeters et al., Reference Peeters, Haeck, Harinck, Afolabi, Demeestere, Audenaert and Höfte2020). The disease is favoured by cool and moist climate with high humidity and moisture stress during reproductive stages (Reddy et al., Reference Reddy, Reddy and Reddy2001). The disease has gained economic importance during last decade causing yield losses of 14–85% (Singh et al., Reference Singh, Singh, Singh, Pandian, Ellur, Singh, Bhowmick, Gopalakrishnan, Nagarajan, Vinod, Singh, Prabhu, Sharma, Mohapatra and Singh2012). The pathogen is seed borne and also survives in diseased plant debris, weeds and soil (Chakaravarthy and Biswas, Reference Chakaravarthy and Biswas1978; Lakshmanan et al., Reference Lakshmanan, Kumar and Velusamy1992; Gopalakrishnan et al., Reference Gopalakrishnan, Kamalakannan and Valluvaparidasan2010; Bigirimana et al., Reference Bigirimana, Hua, Nyamangyoku and Höfte2015; Tajul Islam Chowdhury et al., Reference Tajul Islam Chowdhury, Salim Mian, Taher Mia, Rafii and Latif2015). The pathogen infects through flag leaf sheath and produces phytotoxins viz., cerulenin and helvolic acids which are responsible for production of greyish-brown necrotic lesion in flag leaf sheath and restricts translocation of photosynthates to the developing panicles, causing quantitative and qualitative yield loss including half-filled or chaffy grains, discolouration of grains, reduced seed viability, nutritional and market value (Nandakumar et al., Reference Nandakumar, Babu, Amutha, Raguchander and Samiyappan2007; Gopalakrishnan et al., Reference Gopalakrishnan, Kamalakannan and Valluvaparidasan2010).
The genetic variability and pathogenicity of pathogen are the key factors for determining host plant resistance and for the formulation of viable strategies for disease management. The pathogen S. oryzae is known to be highly variable in cultural characteristics, pathogenicity and genetic traits (Sunil and Patibanda, Reference Sunil Kumar and Patibanda2017). Loss of varietal resistance over time is very common due to variability in the pathogenic population besides the ability of pathogens to develop new and more damaging races. Inherent resistance or tolerance of crop plants to infection by the pathogen can most likely be a safe alternative and most economical and eco-friendly disease management strategy. However, to develop the high-yielding disease-resistant cultivars, it is imperative to analyse and understand the variability in the pathogen. Identification of variability among the isolates of a pathogen is an important step to devise a suitable disease management programme for a particular region and for the development of disease-resistant cultivars in many host pathogen system where major genes control resistance. The present investigation was conducted to study the pathogenic variability in S. oryzae and to identify new sources of resistance against sheath rot disease of rice, besides the status of sheath rot disease of rice was also assessed under temperate agro-ecologies.
Materials and methods
Status of sheath rot disease of rice under temperate conditions of Kashmir
The status of the sheath rot disease of rice was ascertained by undertaking systemic surveys in major rice-growing districts of Kashmir by using multistage sampling scheme technique. Plants from each field were selected in Z transact for recording observations on incidence and intensity of the disease. Total number of plants examined and the number of plants showing sheath rot symptoms were recorded from each plot and per cent disease incidence was calculated.
Isolation and maintenance of S. oryzae isolates
Diseased rice plant samples collected during survey from 18 different locations representing nine districts of Kashmir province were attempted for the isolation of S. oryzae isolates. Pathogenic isolates were isolated by standard pathological technique (Johnston and Booth, Reference Johnston and Booth1983) and then purified by single-spore technique. The single-spore cultures of S. oryzae isolated were maintained on potato dextrose agar slants for studying the variability of the pathogen and screening of rice genotypes for resistance against the sheath rot disease. The pathogenicity of the isolates of S. oryzae was recognized by proving the Koch's postulates.
Variability
The cultural variability of S. oryzae isolates was studied on potato dextrose agar medium. The variability in cultural characteristics viz., colony colour, pigmentation, radial growth and sporulation was recorded at regular intervals. The average spore size (length and width) of each isolate was measured under a microscope. Pathogenic variability of 18 isolates of S. oryzae was tested on a set of 11 genotypes of rice which showed variable resistance response after screening under field and controlled conditions. Based on similarity in disease reaction pattern of the test isolates on inbred rice lines, the isolates were discerned into different pathogenic groups. The average data categorized into 0–9 scale were subjected to cluster analysis to identify the similarity of virulent pattern among isolates. For this analysis, a similarity matrix was derived with the Simqual program (NTSYS 1993 PC, version 1.7) using simple matching coefficient of similarity. A dendrogram was produced by the unweighted pair group method for arithmetic average in the SAHN program.
The virulence of isolates in terms of latent period, virulence index and lesion size was tested on susceptible rice cultivar aromatic zag. Seedlings of the susceptible variety were raised in pots. Three replications were maintained for each isolate. At boot leaf stage, 25-d-old single-spore culture of S. oryzae isolates grown on paddy seeds was placed separately in between the flag leaf sheath and the un-emerged panicle, after gentle pinpricking with a sterilized entomological pin (Vengadeshkumar et al., Reference Vengadeshkumar, Meera, Balabaskar and Jaiganesh2019). Latent period was taken as time in number of days from inoculation to appearance of first disease symptoms. Disease severity was recorded using 0–9 disease rating scale (IRRI, 2013). Virulence index was observed by using the numerical values of disease reaction (disease intensity) and latent period with the formula: Virulence index = Per cent Disease Intensity × 1/latent period.
Disease screening of rice germplasm under field conditions
For the identification of sources of resistance to S. oryzae, a set of 219 genotypes of rice consisting of indigenous and exotic lines along with popular commercial cultivars available at Mountain Research Centre for Field Crops Khudwani, SKUAST-K, were initially screened under artificially inoculated field conditions. The experiment was conducted during Kharif 2020 and 2021 in an augmented design at two locations viz., Mountain Crop Research Station (MCRS) Larnoo (latitude 33.37° N, longitude 75.22° E, altitude: 2286 m amsl) and Mountain Research Center for Field Crops (MRCFC) Khudwani, SKUAST-Kashmir (altitude: 1560 m amsl) SKUAST-Kashmir. SO10 and SO9, the most virulent isolates of S. oryzae were used for inoculation at MCRS Larnoo and MRCFC Khudwani, respectively. Test lines were planted in two row plots of 4 m length with a plant spacing of 15 × 20 cm2. The susceptible checks were planted after every 10 rows and the plot was bordered by susceptible disease spreader rows on each side. Planting was done in the first week of June. Observations on per cent disease intensity and disease score were recorded as per 0–9 disease rating scale (IRRI, 2013) by randomly selected 10 plants from each row. The weather data including mean temperature, relative humidity and total rainfall were recorded at agro-meteorology observatory of SKUAST-Kashmir, Shalimar (online Supplementary Table S2).
Disease screening under controlled conditions
Resistant set of 42 genotypes selected under field screening was further evaluated under controlled epiphytotic conditions with all the collected isolates of S. oryzae to validate their resistance. Seedlings of the selected rice genotypes were raised separately in pots filled with sterilized soil. Fertilizers were applied as per recommendation and watering was done as per the moisture status of the potting medium. After germination two plants were maintained in each pot. The treatments were maintained in a glass house, arranged in a completely randomized block design with three replications per treatment. At boot leaf stage, spore suspension of 25-d-old culture of 18 isolates of S. oryzae was sprayed in evening by using a glass atomizer after gentle pinpricking with a sterilized entomological pin. Observations on disease intensity were recorded as per given procedure and the genotypes were categorized accordingly.
Preparation of inoculum and inoculation
Spore suspension of each isolate was prepared by washing the conidia with distilled water from 30-d-old cultures of S. oryzae. The spore concentration was measured by a haemocytometer and maintained at 3 × 105 spores per ml. Tween-20 was added at 0.1% to the spore suspension. The equal volume of spore suspension of 18 isolates was mixed (Matthiesen et al., Reference Matthiesen, Abeysekara, Ruiz-Rojas, Biyashev, Saghai Maroof and Robertson2016) and spraying of spore suspension was done in evening by using a glass atomizer at boot leaf stage of plants. Disease reaction was recorded by using 0–9 evaluating scale (IRRI, 2013), where 0 means no disease (highly resistant, HR); 1 = 1–10% disease intensity (resistant, R); 3 = 11–25% disease intensity (moderately resistant, MR); 5 = 25–50% disease intensity (moderately susceptible, MS); 7 = 50–75% disease intensity (susceptible, S) and 9 = >76% disease intensity (highly susceptible, HS). From this screening relatively resistant lines from various genetic backgrounds were identified.
Results
Disease status, isolation and maintenance of the sheath rot pathogen
Information regarding the incidence of the disease in a particular agro-ecological region and the diversity of the pathogen are very much essential factors for initiating a suitable disease management programme. Sheath rot disease of rice was found prevalent throughout the Kashmir valley with varied levels of incidence and intensity. The incidence of the disease ranged from 18.8 to 45.7% while the intensity was recorded from 8.7 to 23.6%. High incidence and intensity of sheath rot disease was observed at the locations where mean maximum temperature was moderate (below 30°C) and relative humidity was high (above 80%) during the cropping season. Also, the Japonica rice varieties were found more susceptible as compared to Indica varieties of rice. During symptomatology the sheath rot symptoms were first observed as rotting of uppermost leaf sheaths enclosing young panicles. The lesions start as oblong or somewhat irregular spots, with brown margins and grey centres. The lesions enlarge and coalesce covers most of the leaf sheath. The young panicles remain within the sheath or only partially emerge and an abundant whitish powder growth developed inside the affected sheaths. The grains were discoloured and chaffy (online Supplementary Fig. S1).
For the aetiology of the disease and the nature of the associated pathogen, the causal organism was isolated and purified by single-spore isolation technique from 18 diseased samples representing 18 diverse locations of Kashmir valley besides six rice cultivars and six local land races. Identification of the pathogen was done based on morphological characteristics and the associated pathogen was found as S. oryzae ([Sawada] Games & Hawksworth) (online Supplementary Table S1 and Fig. S2).
Cultural and morphological variability among the S. oryzae isolates
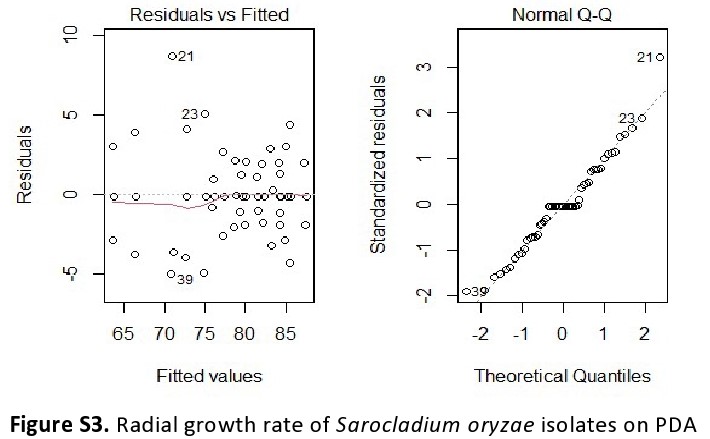
Eighteen single-spore isolates obtained were designated as SO-1 to SO-18. Mycelial growth of the pathogen S. oryzae was measured continuously on potato dextrose agar medium. The radial growth rate of the isolates of S. oryzae was generally slow and varied from 1.0 to 2.5 mm/d on Potato Dextrose Agar (PDA) at 26°C. Identification of variability among the isolates of a pathogen is an important step to develop a disease management programme for a particular region. During the present study a considerable variation among the isolates of S. oryzae was observed in terms of cultural characteristics, morphology and pathogenicity. Variation in colony characters, growth and sporulation among the isolates of S. oryzae was observed on PDA medium (online Supplementary Fig. S4). The radial growth of four isolates viz., SO-9, SO-7, SO-13 and SO-15 was profuse and fast with good sporulation and exhibited colony diameter of 87.4, 85.3, 85.0 and 83.3 mm, respectively, after 38 d of incubation (Table 1). Mycelial growth of SO-11 and SO-3 was very slow. The radial growth rate of S. oryzae indicated substantial diverse and the errors had a normal distribution (online Supplementary Fig. S3). The average spore length recorded ranged from 4.0 to 8.8 μm while the spore width ranged from 0.9 to 2.0 μm.
Table 1. Cultural variability of S. oryzae isolates

++++, excellent >75 conidia per microscopic field; +++, good 51–75 conidia per microscopic field; ++, fair 26–50 conidia per microscopic field; +, poor <25 conidia per microscopic field.
The same letters indicate statistical insignificant differences (p > 0.05)
Disease reaction pattern of S. oryzae isolates
Reaction pattern study of 18 isolates of S. oryzae was recorded on a set of 11 identified rice lines (Table 2). Differences were observed for the mean sheath rot intensity on selected rice genotypes against S. oryzae isolates as well as for isolates across host genotypes while using 0–9 disease rating scale. The isolates of S. oryzae showed varied response in terms of disease reaction against the identified rice lines. The isolates SO-7, SO-9 and SO-15 have been found most aggressive and showed compatible interaction with 10, 9 and 8 genotypes, respectively, while SO-1 and SO-4 was found least aggressive isolates. The genotypes SKUA-354 and GS-183 showed resistant response with the maximum number of isolates while aromatic zag was found susceptible to all the isolates of S. oryzae.
Table 2. Reaction pattern of S. oryzae isolates on selected rice lines

R, resistant reaction (score 0, 1, 3); S, susceptible reaction (score 5, 7, 9).
Cluster analysis based on similarity or dissimilarity in disease reaction pattern exhibited by differential hosts grouped the isolates into three major pathogenic groups (Fig. 1). The pathogenic group-I comprised of isolates SO-7 and SO-9 by way of resistance response from differential hosts. SO-7 and SO-9 developed compatible reactions with 9 and 10 genotypes, respectively. The pathogenic group-II comprised of isolates SO-2, SO-3, SO-5, SO-6, SO-8, SO-10, SO-11, SO-12, SO-13, SO-15, SO-17 and SO-18 while group-III comprised of isolates SO-1, SO-4, SO-14 and SO-16.

Figure 1. Dendrogram showing the virulence similarity and successive clustering of 18 isolates of S. oryzae.
Virulence variability of S. oryzae isolates
The isolates of S. oryzae tested in the present study exhibited considerable variation in the per cent disease intensity, virulence index, incubation period and lesion size (Table 3). Regression analysis of the data indicates that incubation period is negatively correlated with disease intensity, suggesting that greater the number of days taken to incubate less will be disease incidence and vice versa (online Supplementary Fig. S5). Among the 18 isolates of S. oryzae the maximum per cent disease intensity (64.30), virulence index (19.42), lesion length (3.0 × 0.51 cm2) and shortest incubation period (3.31 d) was observed by the isolate SO-9 from Khudwani, Anantnag district, followed by SO-10 from Larnoo, Anantnag district with per cent disease intensity (55.50%), virulence index (7.60), lesion size (1.89 × 0.21cm) and incubation period of 7.31. The isolate SO-1 from Wagam, Budgam district showed the minimum per cent disease intensity (40.23), virulence index (5.20) and the longest incubation period of 7.74 d.
Table 3. Pathogenic variability of S. oryzae isolates

Screening of genotypes under field conditions
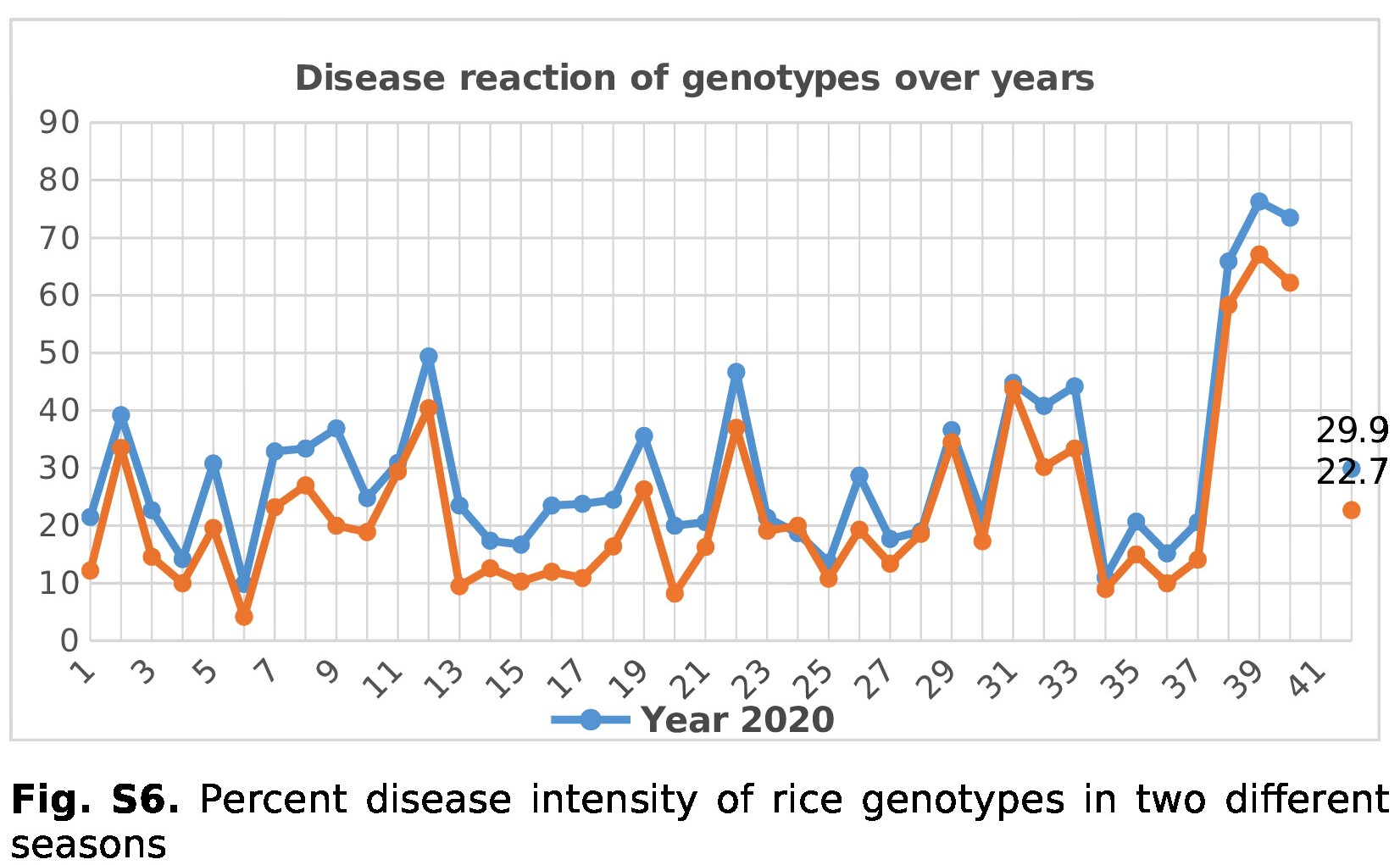
A significant effect (P ⩽ 0.01) of year as well as location was observed in disease intensity of test genotypes. The average disease intensity of 22.7% was observed during 2020 as compared to 29.9% during 2021 (online Supplementary Fig. S6). The effect of maturity period (date of maturity) on disease intensity among the genotypes was also significant. Disease intensity was less on the early maturing genotypes as compared to late-matured genotypes. Two hundred and nineteen genotypes were initially screened for resistance against S. oryzae under field conditions during 2020–2021 in two different environments. Among the test genotypes screened, 25 genotypes (SKUA 292, SKUA 354, SKUA-484, Shalimar Rice-4 [SR-4], ORV-V4, GS-135, GS-139, GS-142, GS-145, GS-150, GS-154, GS-157, GS-169, GS-171, GS-174, GS-183, GS-188, GS-246, GS-248, GS-249, GS-284, GS-324, GS-357, GS-379 and GS-490) were found highly resistant, 131 were categorized as resistant with disease score of 1 and 47 as moderately resistant with disease score of 3 on 0–9 evaluation scale. The remaining genotypes showed moderately susceptible to susceptible reaction.
Screening of genotypes under controlled conditions
The 42 genotypes which showed resistant to highly resistant disease reaction under field screening were further evaluated under controlled conditions against all the 18 isolates of S. oryzae along with susceptible genotypes. Out of 42 test lines, four genotypes (SKUA-354, ORN-V4, GS-183 and GS-357) showed highly resistant disease reaction against S. oryzae. Eleven genotypes (SR-4, SKUA-292, SKUA-415, SKUA-484, ORN-V8, GS-63, GS-142, GS-169, GS-174, GS-379 and GS-490) were found resistant, 18 genotypes (Jehlum, SR-1, SR-2, SR-3, China-1039, SKUA-292, SKUA-478, SKUA-525, ORN-V9, ORN-V10, GS-2, GS-37, GS-88, GS-142, GS-150, GS-248, GS-338 and GS-382) were found moderately resistant and 11 genotypes showed moderately susceptible to highly susceptible disease reaction. Disease severity of resistant rice genotypes during the controlled evaluation ranged from 2.43 to 10.00% (Table 4). The significant (P < 0.05) genotype × environment (G × E) interactions were observed in the present study which indicated that some rice genotypes had varying disease severity scores in the different environments. Successive clustering of 42 genotypes was done based on their disease reaction which clustered the genotypes into four different groups. Group I include the resistant and moderately resistant genotypes. In group-II, four genotypes were included which are highly resistant to sheath rot disease of rice, groups III and IV consists of genotypes which showed susceptible to highly susceptible disease reaction (Fig. 2). The resistant genotypes were registered and deposited at National Bureau of Plant Genetic Resources, Delhi.
Table 4. Disease response of the selected test genotypes against sheath rot disease under controlled conditions


Figure 2. Successive clustering of 42 genotypes on the basis of their disease reaction.
Discussion
Sheath rot is an emerging disease of rice which is causing considerable yield losses particularly on Japonica rice varieties under temperate ecologies of Kashmir. The disease has become endemic in almost all the rice-growing regions around the world in both rain-fed and irrigated ecosystems and is now considered as one of most important emerging and destructive diseases of rice (Raja, Reference Raja2013; Hittalmani et al., Reference Hittalmani, Mahesh, Mahadevaih and Prasannakumar2016). The present study is the first report on temperate Indica and Japonica rice germplasm lines showing resistance/tolerance to sheath rot disease of rice. Indica-type rice lines were, in general, more resistant to sheath rot compared to Japonica rice genotypes regardless of year and location. Although S. oryzae was found to be the causal organism in the present study, sheath rot of rice is a complex disease caused by various fungal and bacterial pathogens. Syafiqa et al. (Reference Syafiqa, Ani, Arif and Achmadi2022) reported Fusarium proliferatum as the responsible pathogen that causes this disease. While the sheath rot disease was found in all the surveyed areas of the Kashmir valley, the intensity of the disease was greater in high-altitude temperate regions where Japonica-type rice is being cultivated under irrigated conditions. Amritpal et al. (Reference Amritpal, Singh, Amrish, Ranbir, Sachin, Ashwani, Singh and Singh2022) also reported the prevalence and distribution of sheath rot disease of rice in subtropical regions of Jammu, India. Maximum sheath rot disease intensity develops at low temperatures between 11 and 20°C, with 73–94% relative humidity during morning hours and 26.0–75.0% during evening hours and sunshine of 2–9 h (Amritpal et al., Reference Amritpal, Singh, Owais, Shafat, Umer, Amrish, Sonali and Ashwani2023). Lanoiselet et al. (Reference Lanoiselet, You, Li, Wang, Shivas and Barbetti2012) reported that higher yield losses due to sheath rot disease of rice occur mostly in lowland environments, particularly in rainy seasons in both rain-fed and upland ecosystems and hot and humid weather favours the disease development. Eighteen isolates of S. oryzae isolated from different locations of the Kashmir valley showed substantial morphological and pathogenic variability. The variability among isolates might have been due to variation in the resistance of host plants, variation in environment, variation in genetic ability of pathogenic isolates or from interaction among these variables. Peeters et al. (Reference Peeters, Haeck, Harinck, Afolabi, Demeestere, Audenaert and Höfte2020) also found variability among the isolates of S. oryzae in terms of growth and toxin production. Cultural and morphological variability between the isolates of S. oryzae was also reported by Vivekananthan and Rabindran (Reference Vivekananthan and Rabindran2008) and Nithin Kumar and Rai (Reference Nithin Kumar and Rai2021). S. oryzae also shows genetic diversity and variability in pathogenicity (Sunil and Patibanda, Reference Sunil Kumar and Patibanda2017). The fungus S. oryzae is thus highly variable in nature and for the first time its variability has also been found under temperate agro-climatic conditions of Kashmir. In the present study, more virulent isolates exhibited a shorter latent period. Regression analysis inferred that the disease severity is negatively correlated with the incubation period (online Supplementary Fig. S5). The shorter latent period benefits the pathogen development while the longer latent period indicates the implication of dilatory resistance by the host as reported by Thakur et al. (Reference Thakur, Rao, Mathur, Tailor, Kushwaha, Dwivedi, Hiremath and Indira2007).
As there are no reports of single gene controlling the resistance against sheath rot of rice, the breeding programme for sheath rot resistance must be supported by new sources of resistance at regular intervals, obtained by continuous screening of germplasm across the years and environment. As the mechanism of resistance among the rice genotypes of temperate ecology is not well understood, in the present study temperate rice germplasm was screened and lines with resistance to sheath rot of rice were identified. Rice genotypes with varied levels of resistance against sheath rot disease have been identified earlier by various workers for other regions (Mohanty et al., Reference Mohanty, Sethil, Sahu and Panigrahi1996; Singh and Raju, Reference Singh and Raju2012; Rini Pal et al., Reference Rini Pal, Dipankar and Naik2015; Mvuyekure et al., Reference Mvuyekure, Sibiya, Derera, Nzungize and Nkima2017). The resistant temperate rice germplasm lines identified in the present study could be used in breeding programme for the development of sheath rot disease-resistant rice varieties suitable for Kashmir and other temperate ecologies.
Conclusion
Sheath rot disease of rice is prevalent in all the rice-growing areas of Kashmir valley having temperate agro-climatic conditions. The Japonica rice varieties are more susceptible to sheath rot disease than Indica rice varieties. Also, it was found that late-maturing varieties are more susceptible than early maturing rice varieties. The significant interaction of genotypes and isolates suggest the specialization in the fungus population because there are variations both in the resistance level of rice genotypes and in the aggressiveness of the pathogen isolates. The resistant sources with varied levels of resistance exist against the sheath rot disease. The determination of genetic basis of these sources and incorporation of their resistant genes into susceptible commercial cultivars could help in the development of high-yielding sheath rot-resistant rice cultivars suitable for Kashmir and other temperate ecologies.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S1479262123000801