



Introduction
Oxynoemacheilus Banarescu & Nalbant, 1966 is a species-rich genus of nemacheilid fishes that is distributed from eastern Greece, Albania and Iran to the southern Caucasus regions. Records of 41 fish species belonging to this genus have been reported in Turkey (Çiçek et al. Reference Çiçek, Sungur-Birecikligil and Fricke2015, Reference Çiçek, Sungur-Birecikligil and Fricke2016, Reference Çiçek, Fricke, Sungur and Eagderi2018; Kaya et al. Reference Kaya, Turan and Ünlü2016; Turan et al. Reference Turan, Kaya, Kalaycı, Bayçelebi and Aksu2019). Among them is the Angora loach, Oxynoemacheilus angorae (Steindachner, 1897), which inhabits drainage basins, streams, and muddy lakes, feeding on benthic invertebrates and spawn (Froese & Pauly Reference Froese and Pauly2020). The Angora loach is terra typica in Ankara (Saylar et al. Reference Saylar, Ata and Firidin2020), having been reported in Marmara, Susurluk, Sakarya, and Kızılırmak basins (Özkan et al. Reference Özkan, Gül, Keleş, Aksu, Kaya and Nur2009; Çoban et al. Reference Çoban, Gündüz, Yüksel, Demirol, Yıldırım and Kurtoğlu2013; Yıldırım et al. Reference Yıldırım, Şen, Eroğlu, Çoban, Demirol, Gündüz, Arca, Demir, Gürçay, Uslu and Canpolat2015; Sağlam et al. Reference Sağlam, Şen, Alnay, Dartay, Özbay, Tuğyan, Türker, Ateşşahin and Sapmaz2017). Furthermore, this species is also found in Israel, Jordan, Lebanon, and Syria.
However, the number of studies on the parasite fauna of Oxynoemacheilus fishes is limited (Koyun et al. Reference Koyun, Ulupınar, Mart and Tepe2016). To the best of our knowledge, 32 trematode species have been reported from fishes of different regions of Turkey (Öktener Reference Öktener2014). Of these the genus Allocreadium is represented by only two species. Allocreadium isoporum (Looss Reference Looss1894) Looss, 1902 was described from nine freshwater fish species in Turkey (Aydogdu et al. Reference Aydogdu2002; Koyun & Altunel Reference Koyun and Altunel2007; Öztürk Reference Öztürk2014; Koyun et al. Reference Koyun, Ulupinar and Gül2015, Reference Koyun, Ulupınar, Mart and Tepe2016; Aydogdu et al. Reference Aydoğdu, de León, Emre, Emre and Yabacı2018), whereas A. lobatum Wallin, 1909 was recorded only from Blicca bjoerkna in the Sakarya River by Akmirza and Yardimci (Reference Akmirza and Yardımcı2014). In recent phylogenetic studies nine valid species of Allocreadium have been confirmed: A. isoporum, A. lobatum, A. gotoi (Hasegawa & Ozaki, 1926) Shimazu, Reference Shimazu1988, A. crassum (Wesenberg-Lund, 1934), A. neotenicum Peters, 1957, A. hemibarbi Roitman, 1963, A. khankaiensis Vainutis, Reference Vainutis2020, A. apokryfi Dos Santos, Gilbert, Avenant-Oldewage & Dumbo, Reference Dos Santos, Gilbert, Avenant-Oldewage and Dumbo2021, A. pseudoisoporum Vainutis, Voronova, Urabe & Kazarin, Reference Vainutis, Voronova, Urabe and Kazarin2023 (Vainutis Reference Vainutis2020; Dos Santos et al. Reference Dos Santos, Gilbert, Avenant-Oldewage and Dumbo2021; Vainutis et al. Reference Vainutis, Voronova, Urabe and Kazarin2023).
Here, we provide information on the species diversity of allocreadiid trematodes from the Angora loach from the Nilüfer stream, Marmara Region (Northwestern Turkey). During detailed morphological and molecular assessment of the material a new Allocreadium species was revealed.
Materials and Methods
Specimen collection and morphological examination
Overall, 68 specimens of O. angorae were collected from the Nilüfer stream (40°10′44.8″N 28°58′13.0″E), Bursa drainage of the Marmara Sea between January 2020 and July 2021. Fish were collected by electrofishing and transported alive to the laboratory, where they were kept in aquaria and examined within 2–3 h. Fish species were killed by vertebral separation. The gastrointestinal tracts were examined under a stereo microscope. Samples of live worms were fixed in 5% formalin or 70% ethanol, depending on purpose.
The whole-mounts of trematodes were prepared using the following method: samples were stained in iron-acetocarmine, differentiated with mixture of hydrochloric acid and ethanol and washed in water; then worms were dehydrated in a graded ethanol series from 50%, 70%, and 90% to absolute ethanol and poured into carbol-xylene; finally, samples were transferred to pure xylene, put on a microslide and mounted in Canada balsam.
All measurements were made with an ocular micrometer and presented in micrometers (μm) unless otherwise stated, with sample size (n) and range followed by mean value in brackets.
Photomicrographs were taken from mounted worms using an Olympus microscope (Shinjuku, Tokyo, Japan) with Digital Camera Leica DFC295 (Leica Microsystems, Wetzlar, Germany). Illustrations were made with the aid of a drawing tube attached to Olympus compound microscope. Prevalence and intensity of infection were calculated following Bush et al. (Reference Bush, Lafferty, Lotz and Shostak1997). Type material including holotype (number ZMADYU 2022/001) and three paratypes (numbers ZMADYU 2022/002–4) have been deposited in the Zoology Museum of Adıyaman University (ZMADYU), Adıyaman, Turkey.
DNA extraction, amplification, and sequencing
Genomic DNA was extracted from three adult worms using an alkaline lysis method HotShot (Truett, Reference Truett and Kieleczawa2006). 28S rDNA fragment was amplified using forward primer U178 (5′-GCA CCC GCT GAA YTT AAG-3′) and reverse primer L1642 (5′-CCA GCG CCA TCC ATT TTC A-3′) (Lockyer et al. Reference Lockyer, Olson and Littlewood2003). Partial cox1 mtDNA were amplified using forward primer JB3 (5′-TTT TTT GGG CAT CCT GAG GTT TAT-3′) (Morgan & Blair Reference Morgan and Blair1998) and reverse primer CO1-R-trema (5′-CAA CAA ATC ATG ATG CAA AAG G-3′) (Miura et al. 2005). The amplification protocol was performed under the following conditions: for 28S rDNA, 1 min denaturation hold at 95°C, 30 cycles of 30 s at 95°C, 30 s at 54°C, 2 min at 72°C and a 5 min extension hold at 72°C; for cox1 mtDNA, 1 min denaturation hold at 94°C, 30 cycles of 15 s at 94°C, 30 s at 50°C, 2 min at 72°C, and a 7 min extension hold at 72°C. Each polymerase chain reaction (PCR) included negative and positive controls, using both primers to detect possible contamination. PCR products were directly sequenced using the Big Dye Terminator Cycle Sequencing kit (Applied Biosystems, Texas, Austin, USA) (following manufacturer’s instructions) and internal sequencing primers only in case with 28S rDNA fragment: 3S (5′-CGG TGG ATC ACT CGG CTC GTG-3′) (Bowles et al. Reference Bowles, Blair and McManus1995), 1200F (5′-CCC GAA AGA TGG TGA ACT ATG C-3′), 1200R (5′-GGG CAT CAC AGA CCT G-3′), 900F (5′-CCG TCT TGA AAC ACG GAC CAA G-3′) (Lockyer et al. Reference Lockyer, Olson and Littlewood2003). The PCR products were read with the ABI PRISM 3130 Genetic Analyzer at the Federal Scientific Center of the East Asia Terrestrial Biodiversity FEB RAS (Vladivostok).
Alignment and phylogenetic analyses
DNA sequences were assembled with SeqScape v.2.6 and aligned in MEGA X software (https://www.megasoftware.net/) with default options (Kumar et al. Reference Kumar, Stecher, Li, Knyaz and Tamura2018). The final alignment of 28S rDNA sequences was trimmed to the length of the shortest—1251 bp long (loaded from GenBank, see Table 1). Genetic distances between the 13 species of Allocreadium were estimated using Tamura-Nei corrected pairwise distances (p values in %) in MEGA X software.
Table 1. Genetic p-distances estimated for Allocreadium spp. based on the 28S rRNA gene sequences

Standard deviation is shown above the diagonal.
Phylogenetic analyses were carried out using four different algorithms: Bayesian inference (BI), maximum likelihood (ML), maximum parsimony (MP), and neighbor joining (NJ). Forty-six sequences of 28S rRNA gene including outgroups were used for the reconstructions. Root-group taxa were presented with four species of the family Callodistomidae (superfamily Gorgoderoidea): Prosthenhystera obesa (NCBI Accession number AY222206), P. oonastica (KM871180), P. caballeroi (KM871183), and P. gattii (MF664223).
BI was conducted in MrBayes 3.1.2 software (Huelsenbeck et al. Reference Huelsenbeck, Ronquist, Nielsen and Bollback2001) using GTR + I + G model (general-time reversible model with gamma distribution and invariant sites) estimated in jModeltest v. 2.1.5 (Darriba et al. Reference Darriba, Taboada, Doallo and Posada2012). The Markov chain Monte Carlo (MCMC) algorithm was performed using 400000 generations and two independent runs; 25% of generations were discarded as burn-in. ML analyses were performed with PhyML 3.0 (Guindon et al. Reference Guindon, Dufayard, Lefort, Anisimova, Hordijk and Gascuel2010) run on the ATGC bioinformatics platform (http://www.atgc-montpellier.fr) with a non-parametric bootstrap validation based on 1000 pseudo-replicates. In calculating maximum likelihood trees, values of G and substitution rate parameters were reset to those estimated by jModeltest, and nucleotide frequencies used were empirical. The MP tree was constructed in PAUP* 4.0 (Swofford, Reference Swofford2002) with a heuristic search. Estimates were obtained using 100 replicates and 100 random addition sequences (saving no more than 1000 trees per replicate), with the TBR (Tree bisection-reconnection) branch swapping option. Gaps were treated as missing data. Branches with a minimum length of zero were collapsed. Data quality was assessed by bootstrapping using 200 replicates under the conditions used to attain the optimal trees. The single most parsimonious tree (length = 682, CI = 0.5674, HI = 0.4326, RI = 0.8229, RC = 0.4670) was recovered from the analysis. The NJ tree was constructed using MEGA X with 100 bootstrap replicates.
Results
Allocreadium bursensis n. sp. (Figure 1)
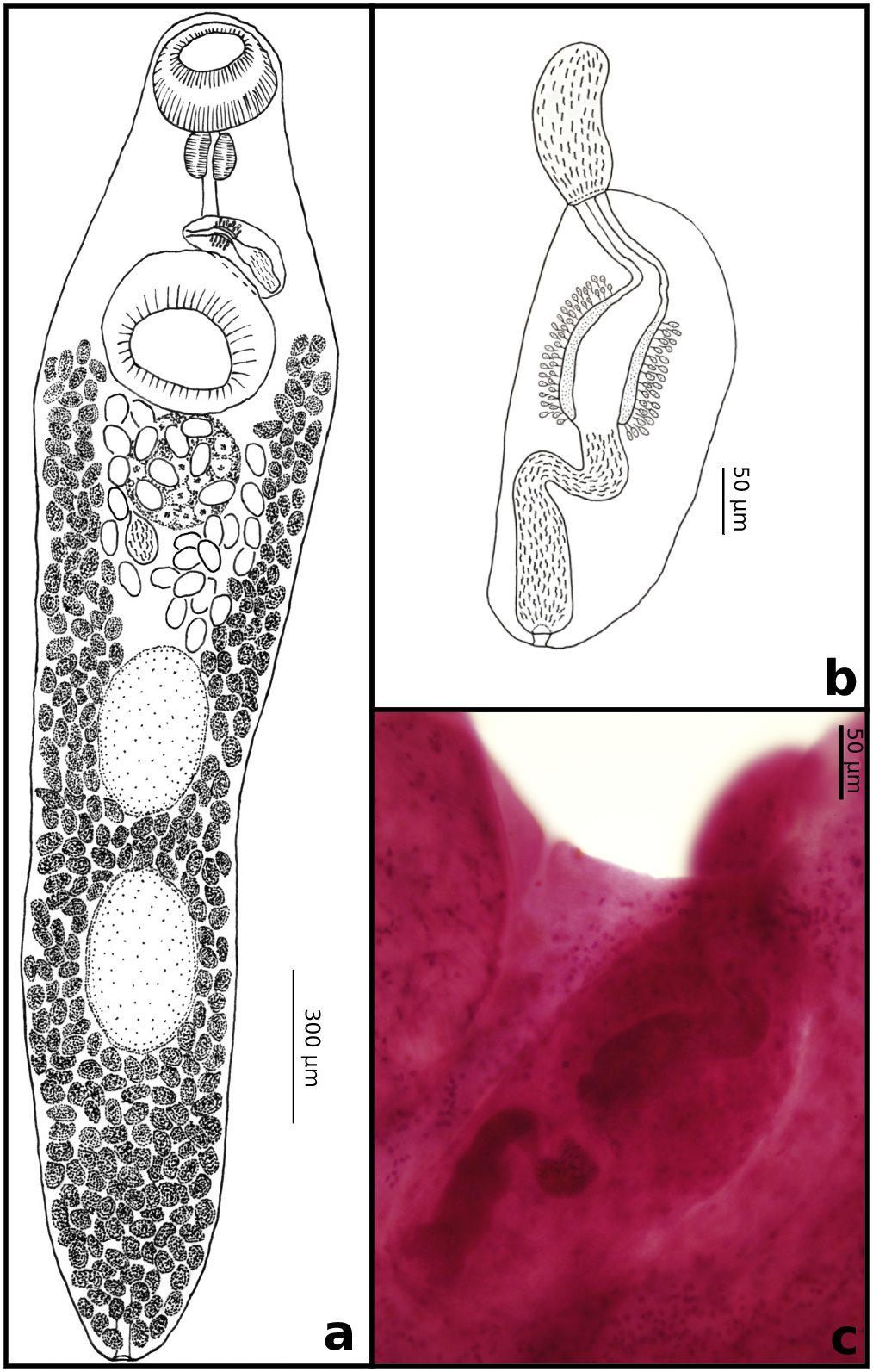
Figure 1. Allocreadium bursensis n. sp. a) Entire adult worm, ventral view; b) schematic drawing of cirrus pouch; c) Photograph of the cirrus pouch with everted cirrus.
Type locality: The Nilüfer Stream, Bursa, Turkey (40°10′44.8″N 28°58’13.0″E)
Type host: Oxynoemacheilus angorae (Steindachner, 1897)
Site of infection: intestine
Prevalence of infection: 32 of 68 (47%)
Mean intensity: 1.7 trematodes per infected host
Type material: Holotype (ZMADYU2022/001) and three paratypes (ZMADYU2022/002–4) have been deposited in the Zoology Museum of Adıyaman University (ZMADYU), Adıyaman, Turkey; additional voucher specimens can be obtained on request from the authors.
Etymology: The specific name, bursensis, refers to the name of the city of Bursa where the type host Oxynoemacheilus angorae was caught from the Nilüfer Stream.
Zoobank registration: urn:lsid:zoobank.org:act:B7621764-11AE-40B4-86B1-608F359D79EA
Genbank registration: Six new sequences were submitted to GenBank under the following accession numbers: OK045521–OK045523 for 28S rRNA gene (1733 bp in length); OQ249866–OQ249868 for cox1 mtDNA gene (816 bp in length).
Description. This description is based on twelve whole mounted specimens: medium-sized trematodes, body elongate, dorso-ventrally flattened, 1485–3024 (2164) long, tapering both anteriorly and posteriorly; maximum body width at level of ventral sucker, 324–621 (468). Eyespot pigments are absent. The tegument is smooth. The forebody is short, narrowed at the anterior end, 300–600 (435) long, occupying 20–19% (20%) of the body length. The hindbody is long, with length 864–2241 (1473). The distance between oral sucker and ventral sucker is 120–310 (210; n = 12). The distance from the posterior end of body to posterior testis is 270–670 (456; n = 12). The posterior end of the body is tapered. Oral sucker oval to spherical, subterminal, muscular lobes and surrounding tegumental papillae are absent, 150–290 long (228), 140–320 wide (218). The prepharynx is absent. The pharynx is bean-shaped, muscular, 67–125 long (95; n = 10), 75–137 wide (100; n = 10). The esophagus is long, bifurcating at the middle level of the ventral sucker (n = 10) or at the posterior border of the ventral sucker (n = 10). The intestinal caeca terminates near the middle, between the posterior testis and posterior region of body (n = 1) or extends to a quarter (n = 1). The ventral sucker is subspherical, 180–400 long (288; n = 11), 210–400 wide (299; n = 11), larger than the oral sucker, and located approximately in the anterior quarter of body. The two testes are large, entire, distinctly ellipsoid, tandem, and contiguous (n = 5) near the middle of the hind body; well separated from the ovary (n = 12). The anterior testis is 150–350 long (259; n = 10), 120–300 wide (206; n = 10); the posterior testis is 190–400 long (303; n = 10), 120–270 wide (211; n = 10). Post-testicular distance is 270–700 long (469). The cirrus sac is relatively large, 187–370 long (231; n = 6), 87–160 wide (113; n = 6) and situated in the midline of the posterior part of the forebody. The internal seminal vesicle is present, saccular convoluted 75 long (n = 1), long, sinuous, enlarged from anterior to posterior; this is part 140 long (n = 1) and saccular convoluted 62.5 long (n = 1) in anterior. Pars prostatica is distinct. The ejaculatory duct is tubular, convoluted, 50–62 long (56; n = 2). Cirrus is unarmed, 110–225 (163; n = 5). The genital atrium is middle sized, overlapping the anterior end of ventral sucker. The genital pore is sinistral (n = 3) or median (n = 3). Ovary is smaller than testes, subspherical, entire, smooth, with the median nearly overlapping the posterior border of the ventral sucker,130–250 long (191; n = 9), 120–230 wide (183; n = 6); the distance between ovary and anterior testis is 70–280 long (141; n = 9). The seminal receptacle is situated between ovary and anterior testis, sub-ellipsoid 75–120 long (97.5; n = 2), 70–75 wide (72.5; n = 5), and often invisible. Vitelline follicles are large, 187–370 long (231; n = 6), 87–160 wide (113; n = 6), ellipsoid or spherical, forming two lateral fields extending from midlevel of the ventral sucker (or level slightly anterior to the ventral sucker) to the posterior end of body, separated anteriorly, confluent in the post testicular region. The uterus is pretesticular, from the anterior margin of the anterior testis to the posterior border of the ventral sucker (n = 1). Eggs are numerous (12–74), large, ellipsoid, 67.5–95 long (79.4; n = 6), 42.5–60 wide (49.6; n = 6). The excretory vesicle is I-shaped, reaching anteriorly to the posterior margin of the anterior testis. The excretory pore terminal is at the middle line of the posterior end of body.
Remarks
The new species ‘bursensis’ can be placed in the genus Allocreadium according to the following morphological features: the body is elongate-fusiform; perioral muscular papillae are absent; the posterior border of vitellarium completely fills the posttesticular space; lateral fields of vitellarium do not connect on the dorsal side of pretesticular space, reaching the forebody or ventral sucker, or not reaching it at some distance. These features are characteristic of Allocreadium species, most of which are found in the Palearctic zone. Among them are A. isoporum, A. hasu Ozaki, 1926, A. japonicum Ozaki, 1926, A. tribolodontis Shimazu & Hashimoto, 1999, A. brevivitellatum Shimazu, 1992, A. hemibarbi, A. aburahaya Shimazu, 2003, A. tamoroko Shimazu & Urabe, 2013, A. khankaiensis and A. carparum Odening, 1959. Allocreadium bursensis n. sp. is also morphologically close to species from Afrotropic and Indomalayan realms: South Africa and South Asia (India), respectively. Here we highlight some species very similar to A. bursensis in morphology: A. isoporum, A. Transversale, and A. Aburahaya from the Palearctic Region, as well as A. kosia from the Indian Region and A. apokryfi from South Africa.
Allocreadium bursensis n. sp. is similar to the type species A. isoporum of Looss (Reference Looss1894) in having a fusiform body and a long esophagus bifurcating dorsally to the ventral sucker. Allocreadium bursensis n. sp. highly resembles A. isoporum of Slusarski (Reference Slusarski1958) in body length and width, size of oral and ventral suckers and eggs, and in having the vitellarium extending to the posterior margin of the ventral sucker. Allocreadium bursensis n. sp. and the Turkish A. isoporum (Aydogdu et al. Reference Aydoğdu, de León, Emre, Emre and Yabacı2018) morphologically diverge in relation to true European A. isoporum in having the vitellarium reaching the mid-level of ventral sucker length vs. the vitellarium not reaching the ventral sucker.
Of the regularly reported species, Allocreadium bursensis n. sp. most closely resembles A. transversale (Rudolphi, 1802) Odhner, 1901 by having the following characteristics: extension of the anterior border of vitellarium to the midlevel of the ventral sucker or posterior to it (Szidat 1938; Bauer 1948; Koval 1957; Roitman 1963; Popiołek et al. Reference Popiołek, Okulewicz and Kotusz2003; Šmiga et al. Reference Šmiga, Fedorčák, Faltýnková, Stefanov, Bystrianska, Halačka, Koščová and Koščo2020); the ventral sucker larger than the oral (Szidat 1938; Bauer 1948; Popiołek et al. Reference Popiołek, Okulewicz and Kotusz2003; Šmiga et al. Reference Šmiga, Fedorčák, Faltýnková, Stefanov, Bystrianska, Halačka, Koščová and Koščo2020); preacetabular position of the cirrus pouch (Koval 1957; Šmiga et al. Reference Šmiga, Fedorčák, Faltýnková, Stefanov, Bystrianska, Halačka, Koščová and Koščo2020). On the other hand, A. bursensis n. sp. differs from A. transversale by having a different ratio of forebody to entire body, longer forebody, vitelline follicles adjoining in the intertesticular space, and lower maximum values of egg sizes.
Considering the keys to Palearctic Allocreadium spp. (Vainutis et al. Reference Vainutis, Voronova, Urabe and Kazarin2023), A. bursensis is closer to the Japanese species A. aburahaya by general morphology. However, the Turkish species differs from the Japanese one in these characteristics: entire ellipsoid testes vs. slightly indented irregularly; entire subspherical ovary vs. globular or somewhat irregular ovary; eyespots absent vs. scattered in the forebody. Since the latter feature is not a characteristic of most Allocreadium spp., only the shape of the testes and ovary distinguish Turkish and Japanese species.
Of the African species, A. bursensis n. sp. has morphological affinity to A. apokryfi in the following features: body length and maximum width at the level of the ventral sucker; intestinal bifurcation at the level of the ventral sucker; preacetabular position of the cirrus pouch; presence of an intertesticular space. These species can be distinguished by the ventral sucker being larger than the oral sucker vs. the ventral and oral suckers being almost equal in size; vitelline follicles filling the intertesticular space; uterus intercaecal, between the ventral sucker and testes vs. inter- and extracaecal uterus; anterior border of vitellarium at mid-level of the ventral sucker vs. not reaching the ventral sucker, in the ovarian region.
Comparison with the Indian Allocreadium species shows that A. bursensis n. sp. is similar to A. kosia (Pande, Reference Pande1938) in having: an elongate body; extension of the anterior border of the vitellarium to the middle of the ventral sucker; the vitelline follicles overlapping the caeca and entering the intertesticular space. However, A. bursensis n. sp. differs from A. kosia by having a smaller body both in length and width; the intestinal bifurcation at midline of the ventral sucker; the ventral sucker smaller than the oral sucker; and the cirrus pouch anterior to ventral sucker.
According to the comparison based on the morphology of the adult trematodes, Allocreadium bursensis n. sp. is evidently a separate species in relation to all previously known Allocreadium spp.
Molecular data
Variability of 28S rRNA gene
Intraspecific variability within Allocreadium bursensis n. sp. was 0%; singleton and parsimony-informative sites were absent. Genetically, Allocreadium bursensis n. sp. was closer to A. pseudoisoporum from Southeast Primorye (Russia), A. hemibarbi, and A. gotoi: 1.78 ± 0.0037%, 3.803 ± 0.0056%, and 3.722 ± 0.0056%, respectively, and more distant from A. apokryfi, 5.825 ± 0066% (Table 1). In general, p-distances between ten valid species and three unidentified lineages (Allocreadium sp. 1, sp. 2, sp. 3) were in the range of 0.162 ± 0.0011–6.068 ± 0.0066%.
Variability of cox1 mtDNA gene
There was no intraspecific variability in the cox1 sequences of Allocreadium bursensis n. sp., and singleton and parsimony-informative sites were absent. According to p-distances, Allocreadium bursensis n. sp. was closest to A. gotoi from Japan (18.777 ± 0.015%) and distant from A. lobatum (22.562 ± 0.016%) (Table 2). Genetic distances between the five valid species and Allocreadium sp. 1 were in the range of 15.915 ± 0.014–22.562 ± 0.016%.
Table 2. Genetic p-distances estimated for Allocreadium spp. based on the cox1 gene sequences

Standard deviation is shown above the diagonal.
Phylogenetic reconstructions
Four phylogenetic trees (Figures 2a–b) based on the 28S gene revealed six monophyletic subclades: Acrolichanus, Stephanophiala, Bunodera, Allocreadium, and three subclades representing polyphyletic Crepidostomum sensu lato: C. chaenogobii, Eurasian Crepidostomum sensu stricto, and American Crepidostomum spp. The order of generic clustering differs depending on the method of reconstruction, except for the genus Acrolichanus, which took basal position, and Allocreadium, which is terminal in each tree (Figures 2a–d). The genus Stephanophiala and Crepidostomum chaenogobii took the closest position to Acrolichanus in BI, MP, and ML reconstructions (Figures 2a,b,d), but in NJ reconstruction (Figure 2c) C. chaenogobii was closer to Acrolichanus, and Stephanophiala was nested amongst Holarctic Crepidostomum spp. (Eurasian sensu stricto and American sensu lato) and Allocreadium with statistical support lower than 50%. A subclade of Holarctic Crepidostomum spp. formed a monophyletic branch with C. chaenogobii and Stephanophiala in MP and ML reconstructions (Figures 2b,d) or with C. chaenogobii and Bunodera in BI reconstruction (Figure 2a). The subclade of Bunodera had an intermediate position between C. chaenogobii and Holarctic Crepidostomum spp. (Figures 2a,c) or was sister to the subclade of Allocreadium (Figures 2b,d). The NJ resolution (Figure 2c) of the genera Bunodera, Crepidostomum sensu stricto, Stephanophiala, and C. chaenogobii was not supported statistically (bootstrap lower than 50%).

Figure 2. Phylogenetic trees of the family Allocreadiidae reconstructed with four methods based on the partial 28S rRNA gene: a) Bayesian Inferrence; b) Maximum Parsimony. Mutation changes and bootstrap support ≥ 50% are shown above and below branches respectively; c) Neighbor-Joining; d) Maximum Likelihood. Trees were rooted with four Prosthenhystera species. Bootstrap support (BI, ML, NJ) ≥ 50% is shown above the branches.
Only BI (Figure 2a) and MP (Figure 2b) trees showed identical, well-supported topology within the genus Allocreadium, where stepwise clustering of eight lineages can be seen (groups a–h). Group a contained A. bursensis n. sp. and A. pseudoisoporum and took the closest position to the ancestral node. Then, towards the top: group b included A. gotoi from Japan and Allocreadium sp. 2 from Ukraine; group c was presented with the single species A. hemibarbi; group d included A. khankaiensis and Allocreadium sp. 1 from the Primorsky region, Russia; group e included two European species, A. isoporum and A. crissum; group f consisted of the single species Allocreadium sp. 3 from China; group g was formed with A. lobatum from North America and A. neotenicum from Europe. The African species A. apokryfi took the terminal position in group h. The resolution of several Allocreadium spp. branches was not supported by NJ (bootstrap lower than 50%), and A. apokryfi took position between a and b groups. Generally, the ML and MP reconstructions (Figures 2b,d) shared the same topology except for the internal species resolution of the genus Allocreadium. In the ML tree (Figure 2d), A. apokryfi was nested closer to the common ancestral node of the genus Allocreadium; the next group consisted of A. bursensis and A. pseudoisoporum; the third one was formed of A. gotoi and Allocreadium sp. 2; the fourth, A. hemibarbi; the fifth, A. khankaiensis and Allocreadium sp. 1; the sixth, A. isoporum and A. crassum; the seventh, A. lobatum and A. neotenicum; and the terminal group, Allocreadium sp. 3.
Discussion
The geography of Turkey is one of the most important factors in its rich biodiversity. There are numerous lakes, ponds, streams, and rivers suitable for fish species, many of which have endemism potential that affects their parasitic fauna (Geiger et al. Reference Geiger, Herder, Monaghan, Almada, Barbieri, Bariche, Berrebi, Bohlen, Casal-Lopez, Delmastro, Denys, Dettai, Doadrio, Kalogianni, Kärst, Kottelat, Kovačić, Laporte, Lorenzoni, Marčić, Özuluğ, Perdices, Perea, Persat, Porcelotti, Puzzi, Robalo, Šanda, Schneider, Šlechtová, Stoumboudi, Walter and Freyhof2014). Ultimately, the parasitic fauna of any host species reflects its history, interaction with the host`s feeding niche, latitudinal range, and social system. Unfortunately, the parasite fauna of Turkish fish (especially nemacheilids) has been poorly studied. In this study we describe a new species from the Angora loach, O. angorae, thereby increasing the number of Palearctic representatives to 26 species.
Revised phylogeny of the genus Allocreadium Looss, 1900
The general morphology of Allocreadium bursensis n. sp. is closest to that of 15 species including ten that represent the Palearctic Allocreadium spp., which are arranged in the keys of Vainutis et al. (Reference Vainutis, Voronova, Urabe and Kazarin2023). Generally, the differentiation of the species was based on a combination of external (ventral and oral suckers’ ratio; body length and width; and its ratio to forebody) and internal (cirrus pouch configuration; uterine extension in hindbody; egg size; disposition of anterior border of vitellarium; esophagus length) features. Based on detailed morphological assessment of found trematodes, we concluded that these worms are significantly different from other Allocreadium spp., in particular A. isoporum from Europe (Looss Reference Looss1894; Slusarski Reference Slusarski1958), but with notable similarity to A. isoporum reported from Turkey (Aydoğdu et al. Reference Aydoğdu, de León, Emre, Emre and Yabacı2018).
The current phylogenetic analysis involved ten valid species, also including Allocreadium sp. 1 from Russia (Primorye), Allocreadium sp. 2 from Ukraine, and Allocreadium sp. 3 from China. The taxonomic status of trematodes collected in Turkey was established as A. bursensis n. sp. based on the interspecific divergence in relation to other valid Allocreadium spp.: 1.78–5.83% in the 28S gene (ten species); 18.78–22.56% in the cox1 gene (five species). Similar values were previously established for the species within the genus Allocreadium (Vainutis Reference Vainutis2020; Vainutis et al. Reference Vainutis, Voronova, Urabe and Kazarin2023) and several genera of the family Allocreadiidae (Vainutis et al. Reference Vainutis, Voronova and Urabe2021).
Internal resolution of the genus Allocreadium on the BI and MP phylogenetic reconstructions (Figures 2a,b) and distribution of its species into groups a–h completely corresponds, as previously shown, with the studies of Vainutis (Reference Vainutis2020) and Vainutis et al. (Reference Vainutis, Voronova, Urabe and Kazarin2023). The NJ tree showed the same topology within the clade of Allocreadium as that presented in study of Dos Santos et al. (Reference Dos Santos, Gilbert, Avenant-Oldewage and Dumbo2021), but statistical support for such a division (subclades h and b–g) was lower than 50% (Figure 2c). The African species Allocreadium apokryfi recently described by Dos Santos et al. (Reference Dos Santos, Gilbert, Avenant-Oldewage and Dumbo2021) had a variable position on the reconstructed phylogenetic trees (Figures 2a–d). In the original description, A. apokryfi was nested between the branches of A. pseudoisoporum (Allocreadium sp.) and A. gotoi based on the BI and ML models with equivocal support for such a resolution (0.8 posterior probabilities/53%) (Dos Santos et al. Reference Dos Santos, Gilbert, Avenant-Oldewage and Dumbo2021). Such topology corresponds to the tree reconstructed with NJ method but was not supported by us (Figure 2c). In our study both BI and PAUP models showed the terminal position of A. apokryfi was well supported statistically—0.98 posterior probabilities and 60% of Bootstrap respectively (Figures 2a,b). The terminal position of this species was recently revealed by Vainutis et al. (Reference Vainutis, Voronova, Urabe and Kazarin2023). Inconsistency was observed in the ML tree (Figure 2d) where A. apokryfi was closer to the ancestral node of all Allocreadium species (87%), but the internal resolution of the other Allocreadium spp. had weak statistical support (31–66%). Based on that, we suggest that A. apokryfi is likely to keep its terminal position in the genus.
In summary, the resolution of the genera and species of the Allocreadiidae on the MP tree is probably the most reliable: the resolution between genera is similar to that of the ML tree and within the genus Allocreadium is fully consistent with the BI tree. The NJ tree topologies showing A. apokryfi not related to A. lobatum and A. neotenicum were signifcantly worse interpretations of sequence data.
Intriguingly, but considering the values of divergence estimated in this study and those of Dos Santos et al. (Reference Dos Santos, Gilbert, Avenant-Oldewage and Dumbo2021), the Far Eastern species Allocreadium hemibarbi is closest to the African A. apokryfi (4.8%). To the contrary, A. apokryfi and A. bursensis n. sp., having habitats more closely related to each other (South Africa and Turkey respectively) than to the East Asian species, show the highest genetic divergence in the 28S gene than the other Allocreadium spp., that is approximately 6%. Based on the phylogenetic reconstructions (Figures 2a,b), we may assume the later origin of A. apokryfi in South Africa compared to the Asian species—West Asian A. bursensis and North-East Asian A. pseudoisoporum, A. gotoi, and A. hemibarbi. Furthermore, taking into account the paraphyletic relationships of A. apokryfi and the Chinese Allocreadium sp. 3 (Figures 2a,b), there is a probability that these worms were formed from the same lineage in the recent past. If there were a morphological description for Allocreadium sp. 3, then we might see symplesiomorphies confirming this hypothesis. The undescribed Chinese Allocreadium sp. 3 isolated from Schizothorax yunnanensis and S. parvus may be host specific to Schizothorax species and therefore could possibly be A. schizothoracis, which has been described from Schizothorax micropogon in India (Pande Reference Pande1938; Kaw Reference Kaw1950) and then reported from S. sinensis in Yanbian County of Sichuan province, China (Wang & Jiang Reference Wang and Jiang1985). Schizothorax spp. have a wide distribution in Central, South, and South East Asia (He & Chen Reference He and Chen2006; Semenchenko & Ostrovskaya Reference Semenchenko and Ostrovskaya2020) and, as well as other schizothoracin genera (Diptychus and Schizopygopsis), have never been observed in the rivers of North East Asia, particularly in the Russian Far East (Bushuev & Barabanshchikov Reference Bushuev and Barabanshchikov2012; Goriyanov et al. Reference Goriyanov, Barabanshchikov and Shapovalov2014). Thus, A. schizothoracis may inhabit only Central, South, and South East Asia.
However, inclusion of the newly obtained 28S sequences in the analysis, which were unexpectedly submitted during preparation of this manuscript by Rajput et al. (unpublished) for the true Indian A. schizothoracis (GenBank Accn. No. OP584922) and the Lithuanian A. transversale (OQ359128-OQ359130; Petkevičiūtė et al. Reference Petkevičiūtė, Stunžėnas and Stanevičiūtė2023), does not confirm the hypothesis concerning the relationships of A. apokryfi and Allocreadium sp. 3. In the study of Petkevičiūtė et al. (Reference Petkevičiūtė, Stunžėnas and Stanevičiūtė2023), 28S sequences of Allocreadium sp. 3 were absent that resulted in tree topology different from that obtained in our work (Figure 2). Thus, A. apokryfi took an intermediate position within the genus and was sister to A. neotenicum and A. lobatum. The quality of the submitted sequence of A. schizothoracis with length of no more than 500 bp, corresponding to only one of the variable domains of 28S rDNA–D2, does not afford performance of a robust phylogenetic analysis. However, preliminary calculation of interspecific divergence showed that A. schizothoracis was closely related to A. apokryfi (5.25%) and highly distinct from Allocreadium sp. 3 (9.69%). The distances’ range (using only D2) was nearly two times higher (0.4–11.7%) than those calculated using the standard data matrix (D1–D3) (Table A.1) due to a smaller number of conservative sites. Therefore, based on these preliminary results, as a possible scenario, we propose two independent lineages formed in East Asia—North East Asia (A. bursensis, A. pseudoisoporum, A. gotoi, A. hemibarbi, and A. khankaiensis) and South East Asia (A. schizothoracis, A. apokryfi, and Allocreadium sp. 3). Allocreadium apokryfi could have speciated indirectly through the North East African lineages that originated from the South East Asian lineage to which A. schizothoracis also belongs.
Biogeographic assumptions
Among allocreadiid genera the genus Allocreadium has the widest species diversity. The genus includes small and medium-sized individuals, parasitic in the digestive system of mostly freshwater fish, which spread throughout Asia, Europe, Africa, North America, and more recently to South America (Caira & Bogea Reference Caira, Bogéa, Jones, Bray and Gibson2005; Shimazu Reference Shimazu2016; Dos Santos et.al. Reference Dos Santos, Gilbert, Avenant-Oldewage and Dumbo2021; Vainutis et al. Reference Vainutis, Voronova, Urabe and Kazarin2023). According to Saoud et al. (1974), 46 species were assigned to the genus Allocreadium. Since that time, numerous studies on the genus Allocreadium from many countries show that there are 104 valid species at present (e.g., WoRMS 2023).
Despite the species diversity reported from different parts of the world, so far, the genus Allocreadium is represented by only two species in Turkey. Allocreadium isoporum was found in Barbus plebejus from the Dogancı Dam Lake (Aydogdu et. al. Reference Aydogdu2002); in Alburnus alburnus from the Enne Dam Lake (Koyun & Altunel Reference Koyun and Altunel2007); in Squalius cephalus from the Akçay Stream (Öztürk Reference Öztürk2014); in Barbus lacerta and Oxynoemacheilus tigris from the Murat River (Koyun et.al. Reference Koyun, Ulupinar and Gül2015, Reference Koyun, Ulupınar, Mart and Tepe2016); in Barbus nilüferensis from the Nilüfer Steram (Aydogdu & Kubilay 2017); in Capoeta antalyensis from the Aksu Stream; in C. caelestis from the Göksu River; in C. angorae from the Fırnız Stream (Aydogdu et.al. 2018); and in Capoeta pestai from the Çayköy stream (Emre & Kubilay Reference Emre and Kubilay2019). Another species, A. lobatum, was found in Blicca bjoerkna from the Sakarya River (Akmirza & Yardimci Reference Akmirza and Yardımcı2014).
Based on our molecular genetic data, we conclude that the divergence of the Allocreadium species did not occur exceptionally linearly from East Asia to the West, as was previously stated (Bogatov & Vainutis Reference Bogatov and Vainutis2022; Vainutis et al. Reference Vainutis, Voronova, Urabe and Kazarin2023). It is likely that the ‘A. isoporum’ reported from Turkey (Aydogdu et al. 2018) is the newly described A. bursensis n. sp. A possible way of migration of A. isoporum from Southeastern Europe (Balkan Peninsula) to Turkey could be with various cyprinid fish as follows: through the Black Sea or Marmara Sea along coasts with desalinated water. Nevertheless, the invasion of A. isoporum to the south of both seas in the Anatolian region seems very doubtful considering the separation of the Balkans and Anatolia by the Bosphorus strait and the Dardanelles, which connect the waters of the Black, Marmara, and Aegean seas. Also, no cases of Allocreadium spp. distribution across the seas have been recorded, except for some that still have not been confirmed by molecular genetic data (e.g., Shimazu Reference Shimazu1988). Allocreadium spp. are usually freshwater helminths infecting cypriniform fish. This limits their distribution through salt water, particularly the migration of A. isoporum from Europe to Turkey. To the contrary, newly estimated genetic distances showed A. bursensis n. sp. and A. pseudoisoporum were closest to A. transversale—5.44% and 4.64% respectively (Table A.1). The latter species is also morphologically similar to A. bursensis n. sp. infecting cobitid and nemacheilid fish in Lithuania (Petkevičiūtė et al. Reference Petkevičiūtė, Stunžėnas and Stanevičiūtė2023), Poland (Popiołek et al. Reference Popiołek, Okulewicz and Kotusz2003; Kuśmierek et al. Reference Kuśmierek, Stefaniak, Skorzewski, Janko, Kotusz and Popiolek2019), and Bulgaria (Šmiga et al. Reference Šmiga, Fedorčák, Faltýnková, Stefanov, Bystrianska, Halačka, Koščová and Koščo2020). The latter is the closest European (Balkan) country to Turkey. Considering this fact, we suggest that the introduction of cobitid or nemacheilid fish through straits (Bosphorus or Dardanelles) resulted in the divergence into two lineages, namely, A. bursensis n. sp. and A. transversale.
The observation of A. lobatum (American species) in the Sakarya Province (bordering the Bursa Province) of Turkey leads us to conclude that there has been either a human-induced introduction or a wrong species identification (Akmirza & Yardimci Reference Akmirza and Yardımcı2014). Thus, the case of finding both A. isoporum and A. lobatum should be examined in the future through molecular genetic analysis after collecting new material in Europe and Turkey. Given that the only molecular data were provided from Europe for A. isoporum without morphological description (Petkevičiūtė et al. Reference Petkevičiūtė, Stunzenas, Staneviciūte and Sokolov2010), it is doubtful whether they belong to the type species.
Conclusions
Data on the Angora loach, O. angorae, morphology, life cycles, biogeography, and molecular genetics are far from complete for most species of the genus Allocreadium. Thus, an integrative approach that includes all of the above criteria cannot be applied to each species of Allocreadium. The present study raises the number of Allocreadium species recorded in Turkey to three and their definitive hosts to 12 species. To date, ten species have been validated by molecular methods including those newly described in this paper: A. isoporum, A. lobatum, A. gotoi, A. crassum, A. neotenicum, A. hemibarbi, A. khankaiensis, A. apokryfi, A. pseudoisoporum, and A. bursensis n. sp. The latter also increases the number of Palearctic representatives of Allocreadium spp. to 26.
Supplementary material
The supplementary material for this article can be found at http://doi.org/10.1017/S0022149X23000287.
Acknowledgements
We thank Lesley Warner from the Parasitology Section of South Australian Museum, North Terrace, Adelaide, Australia for proofreading the article. We also thank Öğr. Gör. Alper BİLSEL from the Faculty of Education, Bursa Uludag University for help and comments on morphology of the adult worms and schematic drawing of cirrus pouch of Allocreadium bursensis n. sp.
Financial support
This research received no specific grant from any funding agency, commercial or not-for-profit sectors.
Conflict of interest
All authors have read and agreed to the published version of the manuscript. The authors declare that they have no conflicts of interest. The authors have also nothing to disclose.
Ethical standard
No ethical approval was required, as this study did not involve clinical trials or experimental procedures. During the study, no treatment/experiment was implemented on live animals. All sampling and laboratory work on fish have complied with the Republic of Turkey Ministry of Agriculture and Forestry animal welfare laws.






