



Introduction
The emergence of extraordinarily large Chalcolithic settlements in the Bug-Dnieper region from the late fifth millennium to the mid-fourth millennium BC marked an unusual trajectory in the evolution of complex societies. Chalcolithic Trypillia mega-settlements often exceeded 200ha and supported thousands of dwellings that were divided by streets and loosely organised into quarters (Chapman et al. Reference Chapman, Gaydarska, Hale, Müller, Rassmann and Videiko2016). Occupied over relatively short periods of a couple of hundred years (Müller et al. Reference Müller, Müller, Rassmann and Videiko2016), these settlements may have hosted either large, permanent populations exceeding 10 000 individuals (Kruts Reference Kruts, Korvin-Piotrovksiy and Menotti2008) or much smaller communities of a few thousand during seasonal gatherings or pilgrimage activities. Alternatively, they may have provided permanent habitation in smaller numbers of contemporaneously occupied houses (Albert et al. Reference Albert2020). Houses were organised into neighbourhoods (Rassmann et al. Reference Rassmann2014), many of which contained pottery workshops, suggesting specialised craft production for neighbourhood consumption (Korvin-Piotrovskiy et al. Reference Korvin-Piotrovskiy, Hofmann, Rassmann, Videiko, Müller, Rassmann and Videiko2016), along with numerous large ‘Assembly Houses’ that may have served as communal neighbourhood forums (Chapman et al. Reference Chapman2014). Whether Trypillia communities were low-density urban centres (Chapman & Gaydarska Reference Chapman, Gaydarska, Fernández-Götz and Krause2016), settlements with ‘proto-urban’ characteristics (Shmagliy & Videiko Reference Shmagliy and Videiko2003), agglomerations without a significant rural component (Müller & Pollock Reference Müller, Müller, Rassmann and Videiko2016), or something entirely different, is as of yet unresolved. In any case, these sites represent a unique intersection between dense occupation and heterarchical forms of social organisation that downplayed hierarchy (Gaydarska et al. Reference Gaydarska, Nebbia and Chapman2019; Hofmann et al. Reference Hofmann2019). Yet, how these forms of organisation may have been expressed in the landscape, if at all, remains undefined. Analysis of the spatial dynamics of livestock pasturing provides one means by which to examine animal husbandry strategies and social relationships beyond the immediate limits of individual mega-sites. Here, we apply carbon (δ13C) and nitrogen (δ15N) isotope analyses to faunal remains recovered from Maidanetske in order to investigate the relationship between pasture partitioning—the practice of allocating separate pasture grounds to different groups of livestock—and Trypillia social organisation.
The Trypillia mega-site of Maidanetske
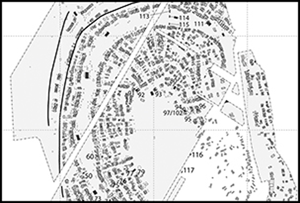
Maidanetske lies in central Ukraine, on the Taljanky River, in the Bug-Dnieper interfluve region, where numerous Trypillia settlements were founded throughout the late fifth and early fourth millennium BC (Figure 1). Occupying some 200ha, it is one of the largest so far identified in the region (Shmagliy & Videiko Reference Shmagliy and Videiko2003). Approximately 3000 rectangular houses (including 2300 structures intentionally burned after abandonment and 700 unburned structures) were arranged in concentric rings, up to eight deep in some areas, encircling a large, open space of approximately 25ha (Figure 2). The rings of buildings were each separated by 70–100m-wide open areas that were possibly used as public spaces; large buildings located in these areas between the rings—the so-called ‘mega-structures’ or ‘Assembly Houses’—probably served as integrative communal facilities (Hofmann et al. Reference Hofmann2019). The position of each ‘mega-structure’ within a cluster of 50–150 smaller dwellings suggests that Maidanetske was organised into distinct neighbourhoods (Müller et al. Reference Müller, Müller, Rassmann and Videiko2016).

Figure 1. Location of the Trypillia settlement of Maidanetske in Ukraine (c. 3800–3650 cal BC; 48°80′37″N, 30°68′17″E) and roughly contemporaneous Trypillia sites nearby (figure by J. Müller, after Müller et al. Reference Müller, Müller, Rassmann and Videiko2016).

Figure 2. Settlement layout of Maidanetske; the concentric rings are rows of burned and unburned houses (figure by J. Müller, after Müller et al. Reference Müller, Müller, Rassmann and Videiko2016).
Radiocarbon determinations obtained from wood charcoal and animal bones (recovered from dwelling structures, pits, special buildings, ditches and pottery kilns), in combination with stratigraphic analyses, establish four phases of occupation at Maidanetske that took place between c. 3960 and 3650 cal BC, with a peak in activity between 3800 and 3700 cal BC (Müller et al. Reference Müller, Müller, Rassmann and Videiko2016; Ohlrau Reference Ohlrau2020). Substantial quantities of charred awns (bristle-like appendages) of feather grass (Stipa sp.) recovered from the upper cultural layers indicate open steppe grassland in the vicinity of the site (Kirleis & Dreibrodt Reference Kirleis, Dreibrodt, Müller, Rassmann and Videiko2016), while abundant charcoal remains mostly of ash (Fraxinus), followed by oak (Quercus) and elm (Ulmus) attest to the presence of closed canopy forests, as well as mixed open- or riverine woodlands (Dal Corso et al. Reference Dal Corso2019).
Emmer (Triticum dicoccum) and einkorn wheat (T. monococcum), as well as barley (Hordeum vulgare) and garden pea (Pisum sativum), were cultivated at Maidanetske, and hazelnuts (Corylus avellana) collected (Dal Corso et al. Reference Dal Corso2019), while livestock husbandry focused on cattle, sheep and pigs (Zhuravlov Reference Zhuravlov2008; Dal Corso et al. Reference Dal Corso2019; Table 1).
Table 1. Number of identified specimens (NISP) and relative faunal frequencies from Maidanetske (different excavation seasons) and Nebelivka (Orton, in Gaydarska Reference Gaydarska2020).

Faunal remains retrieved from the north-eastern part of the site during the 2014–2016 seasons indicate similar frequencies (based on number of identified specimens (NISP)) of cattle and caprines, each accounting for around 40 per cent of the total assemblage; the remains of domesticated pigs contributed approximately 20 per cent (Dal Corso et al. Reference Dal Corso2019; Table 1). The assemblage recovered from the south-western part of Maidanetske during the 2013 season indicates intensive exploitation of cattle (approximately 67 per cent based on NISPgenus), with some caprine herding (approximately 13 per cent) and pig keeping (10 per cent) (see Table 1). The differences in taxonomic frequencies observed in these two assemblages, as well as for fauna analysed from earlier excavations (Zhuravlov Reference Zhuravlov2008), reflect disparate sample sizes and the different analytical units used to calculate relative abundance (MNI vs. NISP), but probably also diachronic shifts in animal exploitation strategies, household variation in animal use and spatial variation in refuse discard across the site. Faunal remains recovered from the 2013 season—the zooarchaeological focus of this study—highlight the likelihood that differences in household-level animal exploitation practices contributed to intra-site variation in faunal distributions. In the south-western part of the site, faunal remains were recovered from several pits that were adjacent to dwellings and filled with midden waste (Pits 50, 52, 60), or from debris from burnt dwellings (Houses 44, 45–50, 52 & 53; see Table S1 in the Online Supplementary Materials (OSM)). A small number of animal bone specimens from Maidanetske were also obtained from deposits on top of the house debris, and in the chernozem (a black soil rich in humus) layers above the house structures. For isotope analyses, bone specimens analysed include those from the 2014 campaign, including from Pits 80-1, 80-2, 80-3 and debris associated with burnt Houses 65 and 66.
The relatively small faunal assemblage recovered from the south-western sector clearly cannot be taken to be representative of Maidanetske as a whole, but it can illuminate the animal management and discard decisions of the households that contributed to the midden deposits. The remains studied highlight the probability that differences in household practices contributed significantly to intra-site variation in faunal distribution. Spatial variation in taxonomic composition, for example, is also apparent in the moderately sized assemblage recovered from the mega-site of Nebelivka, where the frequencies of cattle—the most abundant species—vary widely between house structures, pits and a ‘mega-structure’, relative to sheep/goat and pigs (Orton, in Gaydarska Reference Gaydarska2020; Table 1).
Intensive cattle husbandry for dairy and meat production
The intensive exploitation of cattle by individual households at Maidanetske mirrors animal-use patterns identified at other Trypillia settlements (Zhuravlov Reference Zhuravlov2008; Orton, in Gaydarska Reference Gaydarska2020). Traction-related pathologies identified on cattle bones (Zhuravlov Reference Zhuravlov2008), together with ceramic models of sledges, show that Trypillia cattle were used as draft animals (Ţurcanu & Bejenaru Reference Ţurcanu, Bejenaru, Spinei, Ursulescu and Cotiugă2015; Shatilo Reference Shatilo, Ursu, Poruciuc and Lazarovici2017). Cattle also played an important role in the symbolic repertoire of Trypillia communities. They were depicted on pottery (Ţurcanu & Bejenaru Reference Ţurcanu, Bejenaru, Spinei, Ursulescu and Cotiugă2015), represented as figurines that were perhaps used for ritual purposes (Starkova 2020), and their skulls placed in human burials (Lillie et al. Reference Lillie2017). Cattle may have also been used for feasting, as suggested by an unusual deposit identified at Maidanetske consisting of a pit (no. 50 in Figure 2) that contained a large quantity of bowls, bone debris and two complete cattle skulls (Müller et al. Reference Müller, Müller, Rassmann and Videiko2016).
Biometric data collected from appendicular skeletal elements recovered from Maidanetske inform on the demographic composition of cattle herds. Body-size distribution, calculated using the LSI (Log Size Index) method (Meadow Reference Meadow, Becker, Manhart, Peters and Schibler1999), indicates the exploitation of small-bodied, domesticated cattle (Figure 3; Table S2)—a pattern also seen at Nebelivka (Orton, in Gaydarska Reference Gaydarska2020). The high outlier LSI value of 0.03 (measured from a first phalanx) represents a large-bodied, male animal perhaps used for traction. Mixture analysis, a maximum likelihood method that characterises the mean, standard deviation and proportion of a sampled population consisting of two or more normally distributed groups (Monchot et al. Reference Monchot, Mashkour, Vigne, Vigne, Helmer and Peters2005), was applied to the LSI distribution using PAST v4.09 [(Hammer et al. Reference Hammer, Harper and Ryan2001); outlier representing large-bodied individual removed and Akaike Information Criterion value expressed at two groups]. This analysis indicates even representation of smaller-bodied females and larger-bodied males (Figure 3; female mean = −0.08±0.01, probability = 52.7 per cent; male mean = −0.04±0.01, probability = 47.3 per cent).

Figure 3. Cattle body-size distribution based on log-transformed index (LSI) values, calculated from length and breadth measurements; the solid line indicates mixture distributions (figure by C. Makarewicz).
Epiphyseal fusion data indicate that cattle were slaughtered as adults, with 75 per cent of animals surviving beyond 2.5 years in age (Table 2). Mandibular tooth-wear data (MNI = 5; Table S3) indicate kill-off of cattle at 26−36 months and slightly older, while maxillary deciduous premolars exhibiting moderate wear (dp4; MNI = 2) indicate that younger calves were also slaughtered. Calves are needed to stimulate milk let-down in unimproved breeds. Hence, the harvest of young animals at 6–12 months during the weaning period is consistent with post-lactation slaughter (Gillis et al. Reference Gillis2017). For pigs, tooth wear from a small sample of loose mandibular teeth indicates the slaughter of adult animals aged between 18 and 96 months (MNI = 4) and juveniles between 6 and 8 months (Table S3), while the presence of both unfused and fused bones also indicates the kill-off of young and mature animals.
Table 2. Epiphyseal fusion data for Bos sp. from Maidanetske. Age at epiphyseal fusion according to Silver (Reference Silver, Brothwell and Higgs1969).

The regular exploitation of cows, along with the kill-off of young adults and adults, together suggest the production of both meat and dairy at Maidanetske. Dairy products, even if generated at low levels, would have supplied proteins and fats for household subsistence throughout much of the year. The slaughter of young adults at approximately 26–36 months and older adults is characteristic of meat production. The retention of cattle to adulthood also implies that pasture and fodder resources were sufficient to support cattle herds through multiple winters. Although exploited at relatively low intensities, pigs provided an easily renewable supply of meat in smaller quantities that were more easily consumed at the household level. Low-level sheep and goat herding would also have provided meat and presumably milk, skins and possibly fibre for individual households.
Extensive pasturing of sheep, goat and some cattle
Carbon and nitrogen isotope analysis of faunal bone collagen provides insights into animal management decisions that affect livestock dietary intake, including where to pasture animals and whether to place fewer or more livestock on pastures (extensive vs. intensive pasturing) (Makarewicz Reference Makarewicz, Ventresca-Miller and Makarewicz2018). At Maidanetske, bovid livestock exhibit similar carbon isotope values averaging −20.3‰±0.4 (range = –20.9 to –19.1‰; Table 3; Table S4). Consequently, carbon isotope values of vegetation ingested by livestock range from approximately –26‰ to –24‰ (accounting for a −5‰ diet-collagen fractionation factor), indicating that cattle, sheep and goats all grazed on open C3 pastures, with no discernible input from 13C-enriched C4 species, water-stressed C3 vegetation, or 13C-depleted seasonally inundated floodplain flora.
Table 3. Summary statistics for carbon (δ13C) and nitrogen (δ15N) isotope values for cattle (Bos sp.), sheep (Ovis aries), goat (Capra hircus) and pig (Sus scrofa) from Maidanetske.

* Sheep and goat include specimens from the 2014 excavation season not described in Table 1.
Notably, these carbon isotope values are considerably higher than those that would be expected for herbivores foraging in heavily wooded forest, confirming that cattle and caprines at Maidanetske were pastured on open grasslands. In forests, understorey growth is depleted in 13C due to recycling of CO2 trapped under the leaf canopy and decreased photosynthesis rates caused by low light conditions (Broadmeadow & Griffiths Reference Broadmeadow, Griffiths, Ehleringer, Hall and Farquhar1993). While the carbon isotopic composition of herbivores grazing in dense forest varies according to local environmental and habitat conditions (Stevens et al. 2006), herbivore bone collagen δ13C values ranging from –25‰ to –22‰ are thought to reflect foraging under forest canopy, with lower carbon isotope values expected for dense forest cover and higher values for light forest canopy (Drucker et al. Reference Drucker2011). Collagen δ13C values ranging from approximately –23‰ to –19‰ (mean = –20.9‰) for Neolithic and Eneolithic faunal remains from the Bug-Dnieper interfluve demonstrate that both canopied woodland and more open landscapes were present during these periods (Figure 4; Table S5); directly defining the carbon (and nitrogen) isotopic composition of vegetation undisturbed by livestock grazing at Maidanetske was not possible due to the general paucity of wild taxa in the faunal assemblage.

Figure 4. a) Carbon (δ13C) and nitrogen (δ15N) isotope values of cattle (Bos sp.; blue circles), sheep (Ovis sp.; black ‘X’), goats (Capra sp.; red cross) and pig (Sus scrofa; green diamonds) from Maidanetske. Orange boxes indicate the range of isotope values for wild fauna from Neolithic and Eneolithic sites located in the Bug-Dnieper interfluve region (Lillie et al. Reference Lillie, Budd and Potekhina2011, Reference Lillie2017; see Table S3); b) δ13C and δ15N values of cattle (blue circles) and large mammals (small light blue circles, likely Bos sp.) from Nebelivka (Chapman et al. 2018; Millard, in Gaydarska 2020: 408). The gradient bar indicates the estimated range of collagen δ13C values expected for animals grazing in woodland pastures (green) and open pastures (yellow), as well as the general nitrogen isotopic composition of temperate environment floral growth in foodwebs unperturbed by grazing and anthropogenic modification of soils (figure by C. Makarewicz).
The higher carbon isotope values exhibited by Maidanetske bovid livestock relative to (E)Neolithic wild fauna may reflect new pressures on woodland resources, intensification of herding activities, or both. At Nebelivka, occupation of which overlapped that at Maidanetske (Millard, in Gaydarska Reference Gaydarska2020), livestock exhibit overall carbon isotopic similarity with animals from Maidanetske. Some Nebelivka cattle (which possibly include aurochs; see Table 1), however, exhibit lower values, approaching approximately –22‰, which is consistent with foraging in lightly canopied woodlands, and also higher values up to approximately –19.5‰, which is associated with grazing in more open pastures (Figure 4b). The absence of forest pasturing at Maidanetske as seen in the carbon isotope record—despite charcoal evidence indicating the presence of woodland species associated with canopied forest nearby—may reflect the cumulative effects of long-term harvesting pressures on woodland resources and/or the intensification of cattle husbandry in Trypillia animal management systems. Maintaining a steady wood supply for house building and other construction, as well as for fuel, was important at Maidanetske (Dal Corso et al. Reference Dal Corso2019). Intensive wood harvesting would have reduced forest density, while steady woodland grazing would have impeded tree regeneration (e.g. Bernes et al. Reference Bernes2018). Both practices would have encouraged the growth of 13C-enriched steppe grasslands and open pasturelands. Despite Maidanetske's location adjacent to the Taljanky River, floodplain grazing is not evident in the carbon isotopes of bovid livestock. Riparian environments typically support plants depleted in 13C due to seasonal inundation of soils (Fan et al. Reference Fan, Morozumi, Moaximov and Sugimoto2018).
Dual pasturing systems in use at Maidanetske
Pronounced nitrogen isotopic differences visible in the bovid livestock from Maidanetske reflect variation in animal pasturing strategies that affect the nitrogen isotopic composition of soils and plants. Cattle in particular exhibit an unusually wide range in δ15N values, ranging from 6.3–12.7‰ (Table 3). K-means cluster analysis separating the cattle into two groups indicates that cluster A (Mdn = 11.1‰) exhibited higher nitrogen isotope values than cluster B (Mdn = 7.5‰); a non-parametric Mann-Whitney test indicated that this difference is statistically significant (U = 0, z = −3.1258, p = 0.0018). The isotopic compositions of Neolithic and Eneolithic wild fauna demonstrate animals that foraged in vegetation undisturbed by intensive livestock grazing, exhibiting low nitrogen isotope values that average 6.3‰ (Figure 4; Table S5). The considerable 15N-enrichment in some cattle (cluster A) relative to other cattle (cluster B), sheep and goats clearly indicates that cluster A cattle were intensively grazed—probably at high stocking rates—on pastures that regularly received animal manure. The addition of mineral nutrients and organic matter through manuring contributes to 15N-enrichment of soils via bacterially mediated reactions that preferentially discriminate against isotopically lighter 14N to produce 15N-enriched soil nitrogen pools accessed by plants (Kendall et al. Reference Kendall, Elliot, Wankel, Michener and Lathja2007). Thus, manuring increases the δ15N values of soils and associated flora (Bogaard et al. Reference Bogaard, Heaton, Poulton and Merbach2007; Fraser et al. Reference Fraser2011; Makarewicz Reference Makarewicz2014). Intensive herbivore grazing also promotes, in general, leaching of nitrates (NO3−), ammonia (NH3) volatilisation, and denitrification processes that produce 15N-enriched residual substrates and, thus, higher foliar δ15N values (Robinson Reference Robinson2001; Frank et al. Reference Frank, Evans and Tracy2004). Altogether, higher grazing intensities and/or stocking rates are generally associated with higher herbivore tissue δ15N values (see Szpak Reference Szpak2014).
Previous work has demonstrated that sheep herds intensively grazed as part of large, multi-species herds on spatially restricted pastures exhibit a 2‰ increase in nitrogen isotope values for tissues that formed during the period of intensive grazing—a direct result of their ingestion of 15N-enriched pasture fertilised with manure (Makarewicz Reference Makarewicz2014). The wide nitrogen isotopic spacing between the two cattle groups at Maidanetske suggests that cattle yielding high δ15N values were intensively grazed, probably within a spatially limited pasture area, on a consistent basis. These animals may have included valuable female cattle (i.e. dairying herds), possibly grazed in pastures located close to the settlement to safeguard against rustling and to facilitate milking. The open spaces in the centre of Trypillia settlements, often interpreted as a central enclosure for cattle or, possibly, an agricultural area that was converted to a winter-feeding ground after crops were harvested, may have served as such a pasture. Whether these central, open spaces supported such quotidian subsistence activities, or functioned as a space for ceremonial and regional meetings (e.g. Gaydarska et al. Reference Gaydarska, Nebbia and Chapman2019)—or both—the intensive pasturing of some (but not all) cattle would have required coordination between households, and suggests that some open areas, whether internal or external to the settlement, were managed social spaces.
The low δ15N values exhibited by some cattle, domesticated sheep and goats from Maidanetske—similar to wild fauna—suggest that these livestock were herded using an extensive pasturing strategy that dispersed animals widely across multiple pastures. Extensive grazing connected with low stocking rates is associated with lower δ15N plant values (Frank et al. Reference Frank, Evans and Tracy2004; Makarewicz Reference Makarewicz2014; Szpak Reference Szpak2014). Furthermore, the considerable isotopic variation within Maidanetske bovid livestock exhibiting low δ15N values (cluster B) suggests that animals were variously allocated to diverse pastures, probably according to livestock species, age and/or sex, or intra-community social relationships that influenced pasture access rights. Although it is unclear whether animals were husbanded by individual households or herds were collectively managed by several households within the community, the combination of limited pasture near settlements and proximity—to each other and to Maidanetske—of other, roughly contemporaneous Trypillia settlements, spaced out at approximately 15km intervals within the Bug-Dnieper interfluve (Figure 1), suggests that communities competed or cooperated for grazing resources and that pasture access was constantly negotiated. An extensive pasturing system that distributed animals across the landscape would have also helped maintain a number of animals sufficient for subsistence purposes, regardless of whether the settlement was seasonally occupied by a smaller community or inhabited year-round by a large population.
The use of both extensive and intensive pasturage systems probably entailed different modes of social negotiation to ensure consistent access to forage for livestock. The very high and low nitrogen isotope values exhibited by cattle and large mammals from Nebelivka suggest that the establishment of dual intensive and extensive pasturing systems was underway even earlier, by c. 4000–3750 BC, although the absence of clear nitrogen isotopic grouping within bones identified as belonging to cattle—contrasting to the pattern seen at Maidanetske—may indicate that the dedicated partitioning of cattle into separate pastures was a slightly later development (Figure 4b). Variation in extensive pasture usage, as reflected in nitrogen isotope variation in cattle (cluster B) from Maidanetske, may have been due to careful partitioning of the landscape, determined by pasture availability, intra-community affiliations or inter-settlement access rights. Strong inter-household affiliations, evidenced in the close clustering of dwellings but separation between the clusters that were often further delineated by ditches and lines of pits (Chapman et al. Reference Chapman2014; Rassmann et al. Reference Rassmann2014), suggest that social partitioning expressed within Trypillia settlements may have been further projected onto the landscape. Yet, intensive pasturing of some cattle (cluster A) at Maidanetske also reflects the special character of Trypillia communities—one characterised by a strong cooperative ethos that downplayed social hierarchy within the settlement via integrative socio-political structures.
Pig rearing at Maidanetske
Pig husbandry practices in the ancient world ranged from loose supervision of herds feeding on pannage to highly controlled stall feeding of single animals for rapid weight gain (Halstead & Isaakidou Reference Halstead, Isaakidou, Albarella and Trentacoste2011). Extensively husbanded pigs that forage in woodlands for plant food, as well as insects and worms, typically yield low nitrogen isotope values (Balasse et al. Reference Balasse2013; Hammond & O'Connor Reference Hammond and O'Connor2013). Free-ranging pigs from the early sixth-millennium BC Neolithic settlement of Mağura-Boldul (Romania), for example, show δ15N values of 6.7–6.9‰ (Balasse et al. Reference Balasse2013). In contrast, pigs from Maidanetske yield high δ15N values averaging 11.5‰, indicating that swine were not managed under a pannage system but fed in an anthropogenic environment. High δ15N values in pigs and nitrogen isotopic similarity between pigs and cattle (cluster A), the latter which fed on 15N-enriched plants, may indicate that pigs were eating plant-based foods, such as cereal grains or crop residues; if this is the case, agricultural activities at Maidanetske involved the manuring of cultivation plots. Dung, especially if concentrated in spatially restricted pastures grazed by cattle, would have been easy to collect and transfer to fields to improve soil fertility and increase crop yields. Or, cattle may have been grazed on harvested fields to feed on crop stubble and to fertilise the fields directly. If pigs did ingest cereal crops, their high nitrogen isotope values would indicate that fields were regularly manured at Maidanetske, as sporadic fertilising has little effect on plant δ15N values (Fraser et al. Reference Fraser2011).
While it cannot be excluded that pigs ate some meat or dairy scraps, significant ingestion of protein would probably result in nitrogen isotope values in pigs higher than those observed in herbivorous cattle (Figure 4a). Comparison with the high δ15N values of 10.0–14.3‰ yielded by archaeological fish remains from the Middle and Lower Dnieper region (Budd & Lillie Reference Budd, Lillie, Lillie and Potekhina2020) indicates that the pigs at Maidanetske did not eat fish. Pigs may have been intentionally fed kitchen refuse and meal scraps, or their diets indirectly augmented by foraging on food waste within the settlement. Either feeding strategy would have served the same purpose. First, the conversion of human food discard into food for pigs ensured the consistent generation of fatty meat and lard. Although the relatively low frequency of pigs at Maidanetske indicates that meat production through pig rearing was of lesser importance, pigs that fed on food discard would have more quickly gained weight and attained a larger body size for household consumption (e.g. Halstead & Isaakidou Reference Halstead, Isaakidou, Albarella and Trentacoste2011). Second, these husbandry choices would have served as an effective refuse management strategy—one that quickly removed household food discard and, consequently, disease-carrying vermin from the settlement.
House burning was commonly practised at Trypillia settlements, and is variously interpreted as a deliberate act associated with ritual activities and the closing down of structures or to prevent disease by eliminating pests, amongst other explanations (Chapman Reference Chapman, Gustafsson and Karlsson1999). Recent genomic work identifying the spread of plague (Yersinia pestis) in Neolithic populations across Europe posits that Y. pestis lineages emerged in Trypillia settlements, citing close contact between both humans and livestock, as well as the accumulation of food waste that attracted vermin, creating disease vectors that ultimately contributed to the decline of Neolithic populations in Western Eurasia and Europe (Rascovan et al. Reference Rascovan2019). The isotopic data presented here indicating that most cattle and caprines were husbanded via an extensive pasturing system and that pigs fed on human food discard, however, suggest that Trypillia settlements effectively managed their waste.
Conclusion
The intensive grazing of some cattle, probably dairy cows, at high stocking rates in spatially restricted pastures suggests that high-quality pasture was a limited resource at Maidanetske and that pasture access was negotiated between households, or, if cattle were communally held by several households, through integrative activities perhaps directed within the built environments of ‘mega-structures’. The practice of intensive pasturing itself therefore may have also served socially cohesive functions. Cattle dung, concentrated on specific pastures, may have also been a community resource, available for routine fertilising of agricultural plots. If this was the case, agricultural activities were also tied into the community, even if cropping was undertaken at the household level. At the same time, the partitioning of the local landscape, exemplified in extensive pasturing systems and the different roles played by domesticated animals in those landscapes, also suggests that access to pastures was further determined by intra- and inter-community cooperation or competition. Altogether, this attests to the deep-seated role of careful spatial calculations—most visible within the concentric planning and layout of Trypillia mega-sites—in maintaining and reifying Trypillia social organisation.
Acknowledgements
Faunal materials were provided by joint excavations conducted by the Ukrainian National Academy of Science, the Borys Grinchenko University Kyiv, the Roman Germanic Commission of the German Archaeological Institute, and the University of Kiel.
Funding statement
The 2013 and 2014 excavation seasons were financed by the Institute for Prehistoric and Protohistoric Archaeology (Lehrstuhl Urgeschichte) and the Deutscheforschungsgemeinschaft (project no. GSC 208). Isotope analyses were funded by the Archaeological Stable Isotope Laboratory (ASIL) and faunal analyses supported by the Zooarchaeology Laboratory, University of Kiel. This research was also supported by the DFG (SFB 1266/290391021; R.H., J.M.) and the European Research Council (ERC) under the European Union's Horizon 2020 Research and Innovation Programme (Grant Agreement no. 772957-ASIAPAST; C.A.M.).
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.15184/aqy.2022.32










 Open access
Open access