Introduction
Food legumes or pulses are the staple sources of dietary proteins for more than a billion resource poor people thereby playing a key role in food and nutritional security worldwide. Pulses, in general, contain about twice the amount of protein present in whole grain cereals like wheat and therefore considered an excellent complementary food for infants and young children to meet their daily nutritional requirements. Pulses with greater amounts of lysine, leucine, aspartic acid, glutamic acid and arginine generally supply a well-balanced essential amino acids when consumed with foods rich in tryptophan and sulphur containing amino acids (Boye et al., Reference Boye, Aksay, Roufik, Ribéreau, Mondor, Farnworth and Rajamohamed2010). Plant based proteins have been the subject of growing interest among researchers and consumers because of their potential health benefits as well as their positive environmental effect (Ahnen et al., Reference Ahnen, Jonnalagadda and Slavin2019). Presently COVID-19 pandemic caused potential long-term impact on economic downturn and loss of income all over the world and one of its major consequences is on dietary shift from animal protein to legumes as they are considered a more economical source of quality protein compared to animal-source foods (Rarback, Reference Rarback2020). Chickpea (Cicer arietinum L.) is the second most important food legume crop in the world and presently cultivated in more than 65 countries (FAO 2018). Because of its cultivation worldwide, it can contribute significantly to alleviate the problems of protein malnutrition. It is cultivated on an area of 17.81 million hectares with a production of 17.19 million tonnes, contributing about 20–25% to total pulses production globally. India with a share of about 66% (11.38 million tonnes) of its total global production (FAO 2018) is the largest producer and consumer of chickpea. It provides food for humans as well as feed for livestock; its pod walls and seed coats are also used as feed. Chickpea is an important component of staple diet supplying quality carbohydrate with low glycaemic index and protein to the predominantly vegetarian population in India and considered to be a health food in western countries (Abbo et al., Reference Abbo, Molina, Jungmann, Grusak, Berkovitch, Reifen, Kahl, Winter and Reifen2005). Its protein quality is considered to be better than many other pulses because of the balanced amino acid composition, high bioavailability of its protein and presence of relatively low anti-nutritional factors (Newman et al., Reference Newman, Roth, Newman and Lockerman1987; Friedman, Reference Friedman1996). It is considered a cholesterol reducer food because of its high fibre and unsaturated fatty acid content (Jukanti et al., Reference Jukanti, Gaur, Gowda and Chibbar2012). It is a good source of vitamins such as riboflavin, niacin, thiamin, folate and the vitamin A precursor, β-carotene (Abbo et al., Reference Abbo, Molina, Jungmann, Grusak, Berkovitch, Reifen, Kahl, Winter and Reifen2005).
Protein malnutrition which is widespread among infants and children of many developing countries can be addressed by intake of adequate amounts of foods rich in protein (FAO 2013). Dietary Guidelines for Indians (2011) reported that average consumption of pulses in Indian population is less than 50% of the recommended quantity due to cereals as the preferred staple diet. These guidelines suggested that consumption of pulses in daily diet along with cereals will help to mitigate protein energy malnutrition as cereals are deficient in lysine and pulses are deficient in Sulphur containing amino acids and therefore their mixed diet provide all the essential amino acids by complementing each other. Chickpea is one of the cheapest affordable sources of protein with good nutritional and protein quality. There are reports that chickpea has been specifically utilized for treating protein malnutrition and Kwashiorkor in children (Krishnamurti, Reference Krishnamurti1975). Protein concentration of chickpea ranged from 16–28% and is generally 2–3 times higher than cereal grains (Wood and Grusak, Reference Wood, Grusak, Yadav, Redden, Chen and Sharma2007). Dhawan et al. (Reference Dhawan, Malhotra, Dahiya and Singh1991) based on the characterization of few germplasm lines found that their protein content ranged from 20.9 to 25.27%. Total soluble seed protein content among 30 Indian chickpea cultivars ranged from 16.9 to 29.0% with an average content of 22.6% (Kaur et al., Reference Kaur, Kaur, Gupta, Kaur and Javed2014) indicating the scope for combining high grain yield and increased protein content. Jadhav et al. (Reference Jadhav, Rayate, Mhase, Thudi, Chitikineni, Harer, Jadhav, Varshney and Kulwal2015) characterized 187 desi and kabuli collections for seed protein content that ranged from 13.25 to 26.77%. There are mainly two cultivated types, microsperma (desi) and macrosperma (kabuli), representing two genetically diverse groups (van der Maesen, Reference van der Maesen, Saxena and Singh1987) in the cultivated chickpea species. Ramanujam (Reference Ramanujam and Simmonds1976) reported that the desi chickpea was the first domesticated type and the kabuli type might have been originated from the desi type through mutation followed by conscious selection. The desi types are predominantly grown in the Indian sub-continent and Ethiopia, whereas the kabuli types mostly in the Mediterranean region and Latin America (Singh and Malhotra, Reference Singh, Malhotra, Witcombe and Erskine1984). Desi types have angular seed shape, dark seed coat colour, pigmented stem, pink flowers, semi-spreading to bushy growth habit, greater number of secondary branches, pods/plant, seeds/pod and tolerance to drought and heat. The kabuli chickpea types have owl-head shaped beige-coloured seeds, greater number of primary branches, white flowers, smooth seed surface, lack of anthocyanin pigmentation on stem, greater cold tolerance, a more upright and in some cases taller growth habit (Hawtin and Singh, Reference Hawtin and Singh1980). Pundir et al. (Reference Pundir, Reddy and Mengesha1988) observed that the range of variation for seed size among kabuli types was greater than desi types. The desi types are predominantly cultivated accounting for 85–90% of the global chickpea area and production. Information on characterizing the desi and kabuli chickpea involving large number of cultivars and trait specific germplasm for grain protein content and its quality is limited. The concentration of grain protein in chickpea ranged from 16.7 to 30.6% and 12.6 to 29% in desi and kabuli types, respectively (Gangola et al., Reference Gangola, Khedikar, Gaur, Baga, Varshney and Chibbar2012). Upadhyaya et al. (Reference Upadhyaya, Bajaj, Narnoliya, Das, Kumar, Gowda, Sharma, Tyagi and Parida2016) also found similar levels of phenotypic variation for seed protein content among 244 desi (15.6 to 21.2%) and 92 kabuli (16.2 to 22.4%) accessions. Ozer et al. (Reference Ozer, Karakoy, Toklu, Baloch, Kilian and Ozkan2010) reported variation for grain protein ranging from 17.55 to 23.31% in 91 Turkish landraces of kabuli chickpea. The range of genotypic variation for protein content recorded in previous studies is low compared to 35–40% in soybean, might be because of the limited number of accessions characterized and therefore there is a need for characterization of large number of germplasm to explore the extent of natural variation existing in chickpea for its further improvement.
The inheritance of grain protein content in chickpea was found to be quantitative in nature (Gaur et al., Reference Gaur, Singh, Samineni, Sajja, Jukanti, Kamatam and Varshney2016; Upadhyaya et al., Reference Upadhyaya, Bajaj, Narnoliya, Das, Kumar, Gowda, Sharma, Tyagi and Parida2016) and therefore the phenotypic variation for the trait is likely to be influenced by the genotype, duration of the crop growth phases, location and season and their interaction. However, Singh et al. (Reference Singh, Bejiga and Malhotra1993) showed that the protein content in chickpea seeds was little affected by the growing environment. There are a few relatively high protein genotypes identified in chickpea that are part of its global composite collection (Upadhyaya et al., Reference Upadhyaya, Ortiz, Bramel and Singh2002). Gaur et al. (Reference Gaur, Singh, Samineni, Sajja, Jukanti, Kamatam and Varshney2016) studied inheritance of protein content in ICC 5912, a trait-based (high protein) accession included in the global composite collection. They suggested that the seed protein content in chickpea was a quantitative trait controlled by several genes. ICC 5912 is a pea shaped genotype with grey seed coat and blue flowers and their study showed pleiotropic effects of genes for seed shape and colour on seed size and protein content. The pea shaped and grey coloured chickpea has no consumer acceptability and therefore, search for diverse sources of high grain protein genotypes having desirable seed traits (size, shape, colour) is required for utilizing them in breeding to develop high yielding high protein varieties in chickpea.
The protein quality of chickpea depends on the concentrations of the essential amino acids present in it. Oomah (Reference Oomah2001) reported that proteins characterized by higher levels of branched chain amino acids (BCAA) (leucine, isoleucine and valine) and lower amounts of aromatic amino acids (AAA) are more beneficial to health. Swanson (Reference Swanson1990) found that proteins present in grain legumes are rich in aspartic acid, glutamic acid, lysine, leucine and arginine. The amino acid composition of chickpea is rich in lysine and well balanced except for Sulphur containing amino acids and tryptophan (Ashur et al., Reference Ashur, Clark, Moon and Malzer1973). A better understanding of the extent of variation for grain protein and amino acid profiling would facilitate breeding and development of chickpea cultivars with enhanced grain quality. Identification of germplasm with higher grain protein content and better amino acid composition would greatly facilitate the use of such variability in enhancing its nutritional quality.
Chickpea has great potential for genetic improvement of both quantity and quality of grain protein as well as many other nutritional traits. For several decades, chickpea breeders have focused their efforts on genetic enhancement of grain yield and resistance to major biotic and abiotic stresses, but not improving nutritional quality of the chickpea grain. During the last 50 years, more than 200 improved chickpea cultivars have been released for commercial cultivation in India. A very limited number of studies were undertaken in the past to characterize the large number of chickpea cultivars and germplasm for grain protein, and hence information on this most important nutritional trait is still lacking for many of these cultivars. In this context, further investigations on characterization of chickpea germplasm for grain protein are highly relevant in improving the nutritional quality of chickpea which has a great potential in alleviating protein hunger and malnutrition widespread in developing countries. Chickpea serves as a good and cheap source of protein, particularly for the vegetarian population of India. Therefore, there is an urgent need to understand the genetic variation for grain protein content for improving its nutritional value. The objective of the investigation was to study the extent of genotypic variation for grain protein present in large number of cultivars and amino acid profiling of selected high protein genotypes that is useful in developing high yielding cultivars with better protein profile. Increasing the nutritional value of major food crops like chickpea for alleviating protein hunger and malnutrition is also a priority under the United Nations Sustainable Development Goals.
Materials and methods
Plant material
A panel of four hundred and two chickpea genotypes (303 desi and 99 kabuli types) of diverse origin were grown in two environments with three replications each: one field trial conducted at ICAR-IARI, New Delhi (Rabi 2016–17) and the other at PAU, Ludhiana (Rabi 2017–18). The panel comprised of 113 released varieties, 240 germplasm accessions from the global composite collection, 44 breeding lines and 5 landraces. All the recommended production and protection practices were followed to raise good crops at both the locations. The seeds harvested at full maturity were sorted by removing damaged seeds and foreign material, surface cleaned to remove soil and dust and then oven dried. Thus, obtained seed samples were grounded to fine powder, defatted using hexane (flour: hexane 1:10), dried and then used for the estimation of crude seed protein content. The material also involved five of the trait-based desi accessions (ICC 1932, ICC 5384, ICC 5912, ICC 6969 and ICC 8397) having relatively high seed protein that are part of the global composite collection of chickpeas (Upadhyaya et al., Reference Upadhyaya, Furman, Dwivedi, Gowda, Baum, Crouch, Buhariwalla and Singh2006).
Seed crude protein estimation
The powdered sample (0.2 g) weighed on Whatman filter paper, wrapped in it and placed in the digestion tubes along with 3 g of digestion mixture (CuSO4: K2SO4; 1:10) followed by addition of 10 ml of conc. sulphuric acid just before digestion. The digestion was carried out on Kjeldahl digestor (KELPLUS KES 20L VA DLS TS) during which brown coloured medium was changed to colourless solution. After digestion, 20 ml of distilled water was added to each digestion tubes and distillation was carried out in distillation unit [KELPLUS CLASSIC–DX VATS (E)]. Then the samples were titrated with 0.1N HCl on a titrator (Metrohm 877 Titrino plus) to estimate the nitrogen content. Blank tubes without sample were also run along with samples. The nitrogen content thus estimated was converted to protein content by using conversion factor of 6.25.
FTIR analysis
The sample powder (5 mg) was dissolved in 1 ml of 50 mM phosphate buffer (pH 7.4). Potassium bromide (KBr) discs were prepared by grinding 1 g of dry powder and made into pellets with manual hydraulic press (Hardoim et al., Reference Hardoim, Guerra, Costa, Serrano, Sanchez and Coelho2016). Infrared spectrum was obtained in a transmission mode from each pellet containing a single drop of sample, with KBr alone as a background using a Thermo Nicolet 6700 spectrometer (USA).
Amino acid analysis
Fifteen contrasting chickpea genotypes (online Supplementary Table S1), comprising of three low protein (18.19–19.75%) and twelve high protein (24.84–33.56%), were selected on the basis of variation in protein content and analysed for amino acid profile. Chickpea seeds were homogenized into fine powder. The samples were hydrolysed in evacuated test tubes under acidic (6N HCl) and alkaline conditions (6N NaOH for tryptophan only) by incubating at 110°C for 24 h. Methionine and cysteine were analysed as methionine sulphonate and cysteic acid after cold performic acid oxidation overnight before acid hydrolysis. The samples were cooled to ambient temperature and neutralized by adding 6N NaOH to acid hydrolysed sample and 6N HCl to alkaline hydrolysed. The contents were shaken well and filtered through 0.22 μ nylon filter and analysed by High Performance Liquid Chromatography (Agilent HPLC 1260 infinity) equipped with UV 338 nm detector, C 18 column (200 mm × 4.6 × 5 μm). The mobile phase constituted 40 mM sodium sulphate buffer pH 7.5; acetonitrile; methanol :H2O (45:45:10) with a flow rate of 0.45 ml/min. The operating temperature was set at 40°C.
Amino acid score (Chemical score) was calculated with reference to FAO/WHO standard amino acid profile for humans (WHO 1985). Recommended values of amino acid are given in Table 4.

Statistical analysis
Analysis of variance (ANOVA) was carried out by using SPSS statistical program. Frequency distribution curve and correlation matrix were obtained by using XLStat statistical program (Addin soft SARL, New York, NY, USA).
Results
Genotypic variation for seed protein content in chickpea germplasm
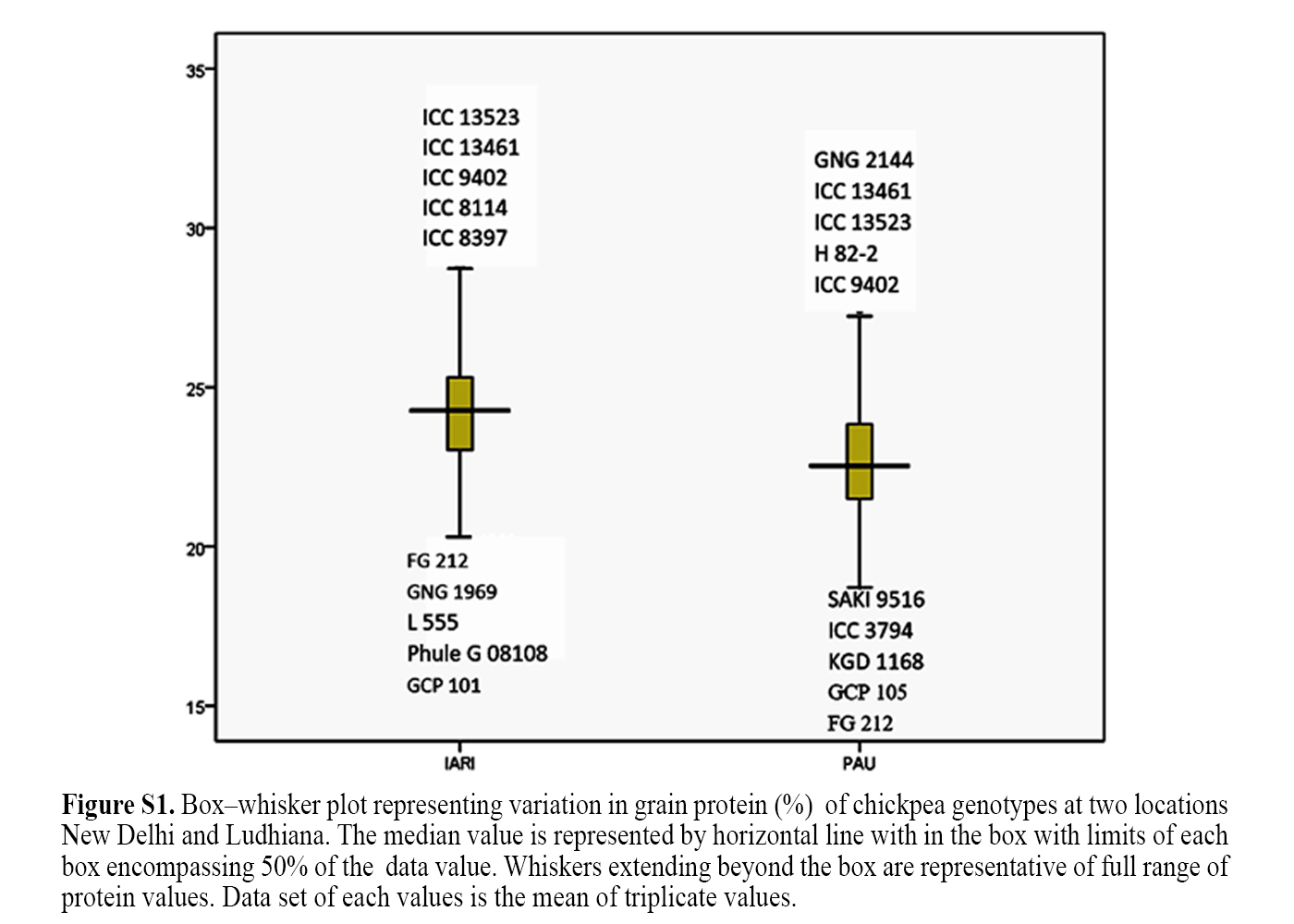
Mean and range of seed protein content estimated on four hundred and two chickpea germplasm at two locations (ICAR-IARI, New Delhi and PAU, Ludhiana) are given in Table 1. The mean seed protein content among the chickpea germplasm studied ranged from 18.19–33.56% (mean 24.30%) and 18.72–30.44% (mean 22.74%) at New Delhi and Ludhiana, respectively. The mean protein content among desi and kabuli genotypes ranged from 18.19–31.72% (mean 24.09%) and 19.53–33.56% (mean 24.93%), respectively at New Delhi; and 18.72–30.44% (mean 22.66%) in desi and 20.00–28.31% (mean 22.99%) in kabuli at Ludhiana. The mean protein contents among commercially cultivated varieties were relatively lower than germplasm accessions and it ranged from 18.19–26.28% (mean 22.74%) and 18.72–30.44% (mean 22.68%) at New Delhi and Ludhiana, respectively. Whereas the mean seed protein content among germplasm accessions ranged from 20.00–33.56% (mean 25.10%) at New Delhi and 19.41–28.79% (mean 22.90%) at Ludhiana. Pusa 5023 (26.28%), Pusa 417 (26.00%) and Pusa 112 (25.53%) at New Delhi and GNG 2144 (28.81%) and H 82-2 (28.29% at Ludhiana are some of the cultivars that recorded relatively higher seed protein content. The mean seed protein content in breeding lines ranged from 22.34-25.88% (mean 24.16%) at New Delhi, while it ranged from 20.03–24.19% (mean 21.73%) at Ludhiana. The improved breeding lines such as BG 1044 (25.88%), BG 1073 (25.69%), BG 1054 (25.63%), BGD 9812 (25.75%), BG 1077 (25.44%), BG 3054 (25.34%) at New Delhi and BG 2059 (25.31%), BG 1044 (24.19%) and BG 1054 (24.06%) at Ludhiana recorded significantly higher seed protein content as compared to others. The mean seed protein content of few landraces included in the study ranged from 24.19–25.16% (mean 24.54%) at New Delhi; and 21.06–24.19% (mean 22.12%) at Ludhiana. Among the landraces, Katila, a landrace having black tuberculated seed coat was found to have the highest protein content of more than 23% at both the locations. Frequency distribution of chickpea genotypes for seed protein content in different groups is given in Fig. 1. The study also showed that about 75 to 80% of genotypes contained seed protein in the range of 21 to 27% at both the locations (Fig. 1). The Box plot analysis led to identification of contrasting genotypes with the highest and the lowest protein content at both the locations (online Supplementary Fig. S1) and these included both desi and kabuli chickpea. The genotypes identified for higher seed protein in New Delhi are, ICC 13523 (33.56%, kabuli), ICC 13461 (31.72%, kabuli), ICC 9402 (31.06%, kabuli), ICC 8114 (30.22%, kabuli) and ICC 8397 (30.22%, desi). The genotypes for higher seed protein content at Ludhiana are GNG 2144 (30.44%, desi), ICC 13461 (28.79%, kabuli), ICC 13523 (28.31%, kabuli), H 82-2 (28.28%, desi) and ICC 9402 (28.10%, kabuli). Pooled analysis of coefficient of variability (CV) for seed protein content indicated significant variation in genotypes and genotype × location interaction (Table 1). Interactions between genotypes as well as genotype × location were significant for released varieties, germplasms, breeding lines, landraces and desi and kabuli seed types for seed protein content. Combined analysis revealed that seed protein content was higher among germplasm evaluated in New Delhi compared to those at Ludhiana.

Fig. 1. Frequency distribution of grain protein content in chickpea genotypes at IARI, New Delhi and PAU, Ludhiana.
Table 1. Mean, range, mean squares of combined ANOVA (Mean Sum of Squares, MSS), interaction and coefficient of variability (CV) for grain protein in chickpea at ICAR-IARI, New Delhi (Rabi 2016–17) and PAU, Ludhiana (Rabi 2017–18)

* p=0.05.
The characteristics of genotypes selected for high seed protein content (average >27/100 g protein) at New Delhi and Ludhiana are given in Table 2. Twelve of the eighteen high protein genotypes were kabuli chickpea that included both small and large seeded types. However, all the high protein desi genotypes were small seeded. The characteristics of chickpea released varieties selected for high seed protein content (average 24–25/100 g and more) at New Delhi and Ludhiana are given in Table 3. The selected cultivars included almost equal number of desi and kabuli seed types. The seed size (22.3 to 38.2 g/100 seeds) of high protein containing kabuli cultivars was found to be greater than those of desi types (13.0 to 23.1 g/100 seeds).
Table 2. Characteristics of chickpea accessions selected for high seed protein content (average >27/100 g) at New Delhi (Rabi 2016–17) and Ludhiana (Rabi 2017–18)

Seed type: D, Desi; K, Kabuli and P, Pea.
Table 3. Characteristics of released varieties selected for high seed protein content (average 24–25/100 g and more) at New Delhi (Rabi 2016–17) and Ludhiana (Rabi 2017–18)

Seed type: D, Desi; K, Kabuli.
FTIR spectrum
Two contrasting chickpea genotypes with the highest (ICC 13523) and the least (FG 212) seed protein content were analysed by Fourier transform infrared (FTIR) spectroscopy. Spectra observed for ICC 13523 (high protein) and FG 212 (low protein) genotypes between 500–4000 cm−1 is divided into three categories: weaker, medium and stronger (Fig. 2). Splitting of peaks between ~658–970.6 and ~3645–4000 cm−1 was observed to be more in spectrum analysis of FG 212 as compared to ICC 13523 which gave broad and prominent peaks. The spectra of prominent peaks in FG 212 was at ~2365, ~1552 cm−1 and in ICC 13523 at ~1637, ~1007 cm−1 correspond to protein content in seed samples. Band assignment in water for α helix and β sheets is between 1648–1657 cm−1 and 1623–1695 cm−1, respectively (Jackson and Mantsch, Reference Jackson and Mantsch1995). The O-H stretching vibration peaks corresponding to wavelength ~3500–4000 cm−1 was maximum in FG 212 whereas in ICC 13523 no such peaks observed. Amide A associated with amide specific group N-H was observed in FG 212 whereas amide IV associated with amide specific group O-C-N was present only in ICC 13523.

Fig. 2. FTIR spectra of two contrasting chickpea genotypes for grain protein content: FG212 (Least protein) and ICC13523 (High protein).
Amino acid analysis
The range (mean) of essential amino acids (g/100 g protein) for histidine, isoleucine, leucine, lysine, methionine, phenylalanine, threonine, tryptophan and valine in fifteen contrasting chickpea genotypes selected for grain protein varied from 2.20–4.75 (2.88); 1.96–5.90 (4.22); 5.61–9.07 (7.64); 6.00–8.19 (7.30); 0.33–2.11 (0.97); 2.27–4.89 (4.06); 2.24–5.03 (3.63); 0.58–2.89 (1.41) and 2.90–5.80 (3.80), respectively (online Supplementary Table S1). The most abundant amino acid in chickpea was found to be leucine followed by lysine, isoleucine, phenylalanine, valine, threonine and histidine. High protein chickpea genotypes had higher lysine content, with lysine content greater than 8.0/100 g in BGD 132 (kabuli, large seeded) and T39-1 (desi, small seeded) chickpea genotypes.
The range (mean) of non-essential amino acids (g/100 g protein) varied from 4.04–6.88 (5.73) for alanine; 6.04–995 (8.20) for arginine; 8.54–14.00 (10.91) for aspartic acid; 0.95–3.06 (1.66) for cysteine; 10.96–20.01 (15.08) for glutamic acid; 3.07–5.68 (4.24) for glycine; 2.90–6.46 (4.06) for proline; 3.59–6.31 (5.30) for serine; and 1.65–2.89 (2.50) for tyrosine (online Supplementary Table S1). During hydrolysis glutamine and asparagine are completely converted to their acidic forms as glutamic acid and aspartic acid, respectively (Pickering and Newton, Reference Pickering and Newton1990). So the content of aspartic acid is representation of both aspartic acid and asparagine. Similarly, glutamic acid represents both glutamic acid and glutamine.
The average essential amino acid scores (Table 4) of six amino acids namely isoleucine, leucine, lysine, phenylalanine, tyrosine and threonine were found to be higher in high protein genotypes whereas for Histidine and Sulphur containing amino acids (Cysteine and Methionine) the scores were higher in low protein genotypes. Sulphur containing amino acids were found to be more limiting in high protein genotypes. However, tryptophan was not found to be a limiting amino acid in all the chickpea genotypes characterized in the present study.
Table 4. Comparison of essential amino acid content (g/100 g) in chickpea seeds with amino acids in reference proteins and pattern for protein requirement

BCAA refers to branched chain amino acid.
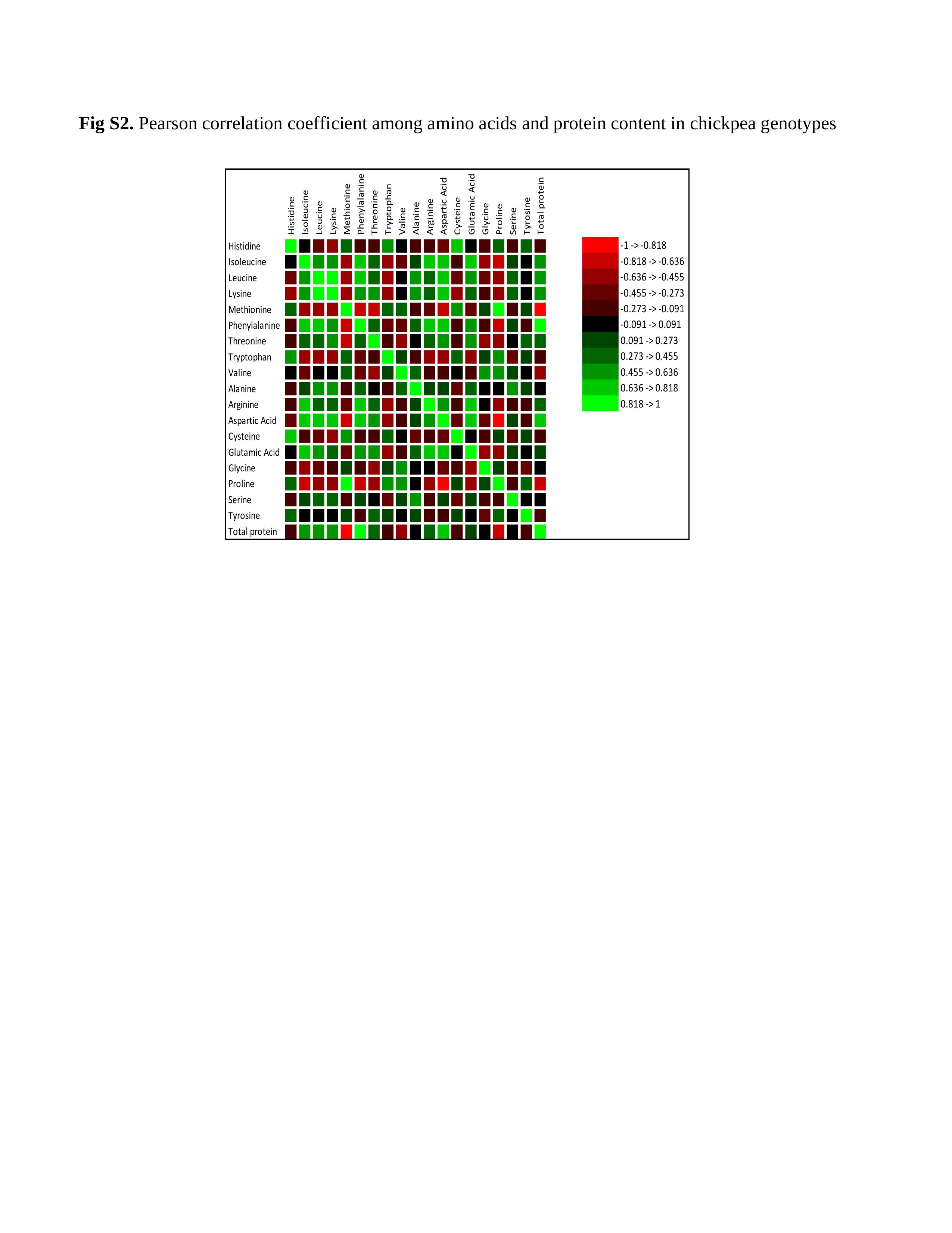
Online Supplementary Fig. S2 depicts the correlation map that uses a green-black scale to display the correlations. The black colour corresponds to a correlation close to −1 (example isoleucine vs histidine) and the green colour corresponds to a correlation close to 1 (example total protein vs phenylalanine). Total protein content showed significantly positive and strong correlation with phenylalanine (r = 0.836), aspartic acid (r = 0.621) and isoleucine (r = 0.569). It was observed that aspartic acid had significant positive correlation with isoleucine (r = 0.776), leucine (r = 0.635), lysine (r = 0.554), phenylalanine (r = 0.780), threonine (r = 0.503) and arginine (r = 0.700). Aspartic acid is synthesized via the transamination of oxaloacetate and is the precursor of threonine, isoleucine and lysine. Lysine showed positive significant correlation with phenylalanine (r = 0.611) which is an essential amino acid. The seed protein content showed negative correlation with methionine (r = −0.836) as well as cysteine (−0.273).
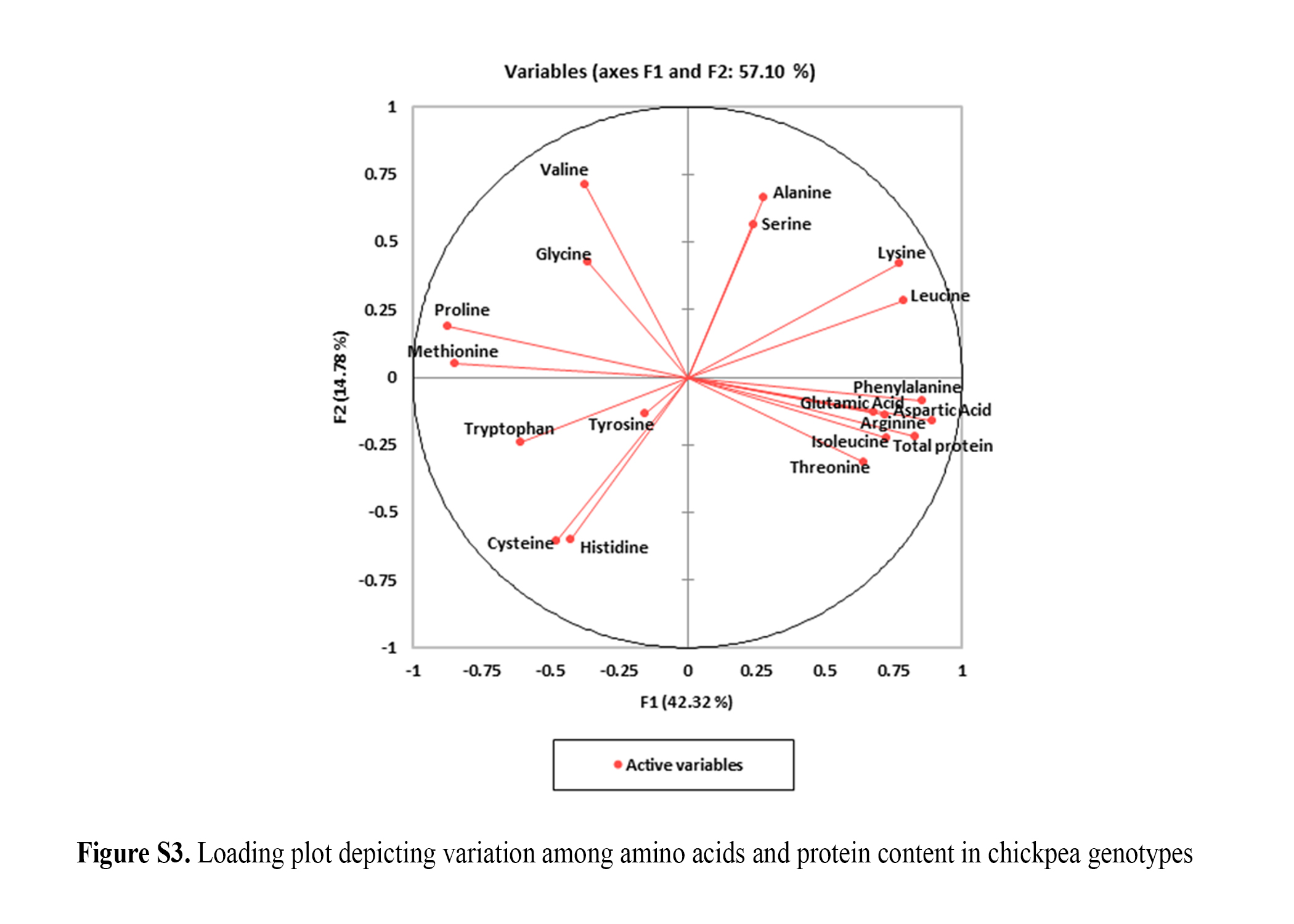
Loading plot (online Supplementary Fig. S3) depicting variation among amino acids and protein content indicated that tryptophan, tyrosine, cysteine and histidine showed negative correlation with all other amino acids and total protein. The plot described that line of one amino acid lie close to the other amino acid indicating that these two amino acids were highly correlated. Phenylalanine, aspartic acid, glutamic acid, isoleucine, arginine and total protein were highly correlated. Methionine, proline, glycine and valine showed no correlation with alanine, serine, lysine, leucine, phenylalanine, aspartic acid, glutamic acid, isoleucine, arginine and total protein. Two principal components were extracted having higher eigen values. Cumulative percentage of the two components explained a total of 57.10% variability with their individual contribution being 42.32 and 14.78%, respectively.
Discussion
Malnutrition caused by inadequate intake of nutrients such as protein, minerals and vitamins are increasing at an alarming rate in developing countries. Growing chickpeas with their excellent protein value could be a way to fight protein malnutrition in the poorest and most arid areas of the world (Varshney et al., Reference Varshney, Thudi, Nayak, Gaur, Kashiwagi, Krishnamurthy, Jaganathan, Koppolu, Bohra, Tripathi, Rathore, Jukanti, Jayalakshmi, Vemula, Singh, Yasin, Sheshshayee and Viswanatha2014). In a long chain of nutrition on global scale, chickpea has a great potential to serve as an important source of protein for the increasing vegetarian population in developed as well as developing countries (Bar-El Dadon et al., Reference Bar-El Dadon, Abbo and Reifen2017). Plant proteins being more economical in meeting the environmental and social aspects can reduce the dependence on animal protein in human nutrition (Almeida et al., Reference Almeida, Nayfach, Boland, Strozzi, Beracochea, Shi, Katherine, Pollard, Sakharova, Parks, Hugenholtz, Segata, Kyrpides and Finn2020). Chickpea is reported to contain 15 to 30% of grain protein compared to 35 to 40% in soybean which is known as a source of high quality protein. A better understanding of the extent of natural genotypic variation existing in the germplasm of cultivated chickpea is helpful in identification of genotypes likely with greater amounts of grain protein that can be utilized as parental material for breeding of cultivars with enhanced protein content. The finding of high yielding high protein cultivars would help in directly promoting the production of such protein rich cultivars for increasing availability and consumption of high quality chickpea to overcome protein malnutrition. The characterization of 402 germplasm comprising of more than 100 commercially grown cultivars of India, trait specific genotypes from the global composite collection, few landraces and improved breeding lines in the present study resulted in the identification of chickpea germplasm containing up to 33.56% grain protein. The average protein content at the two locations combined together ranged from 18.19 (FG212) to 33.56% (ICC13523). It was observed that the grain protein content of commercial cultivars was relatively lower (18.19–28.81%) than the trait specific germplasm accessions (19.41–33.56%) suggesting that there is further scope for improving the grain protein contents of chickpea cultivars through recombination breeding. The grain protein content recorded in the present study was about 5 to 10% greater than some of the recent studies on germplasm characterization such as Kaur et al., Reference Kaur, Kaur, Gupta, Kaur and Javed2014 (16.9–29.0%), Jadhav et al., Reference Jadhav, Rayate, Mhase, Thudi, Chitikineni, Harer, Jadhav, Varshney and Kulwal2015 (13.25–26.77%) and Upadhyaya et al., Reference Upadhyaya, Bajaj, Narnoliya, Das, Kumar, Gowda, Sharma, Tyagi and Parida2016 (15.6–22.4%). The frequency distribution of chickpea genotypes for seed protein content in different groups indicated the presence of high level of genotypic diversity among the germplasm evaluated at the two locations (Fig. 1). The distribution of variation was found to be normal for seed protein content suggesting the quantitative nature of their inheritance in chickpea. The quantitative nature of inheritance of grain protein content in chickpea was also observed by Gaur et al. (Reference Gaur, Singh, Samineni, Sajja, Jukanti, Kamatam and Varshney2016) and Upadhyaya et al. (Reference Upadhyaya, Bajaj, Narnoliya, Das, Kumar, Gowda, Sharma, Tyagi and Parida2016). The study also showed that about 75 to 80% of the genotypes contained seed protein in the range of 21 to 27% at both the locations. A similar range for seed protein contents of chickpea has been reported by different workers globally. Protein content ranged from 12.4–31.5% in Canadian chickpea (Wang and Daun, Reference Wang and Daun2004); 24.63% in Egyptian cultivars (Arab et al., Reference Arab, Helmy and Bareh2010); 14.6–23.2% in European chickpea germplasm (Serrano et al., Reference Serrano, Carbas, Castanho, Soares, Patto and Brites2017); 18.2–20.5% in Italian chickpea genotypes (Summo et al., Reference Summo, De Angelis, Ricciardi, Caponio, Lotti, Pavan and Pasqualone2019): 27.56 in a Turkish cultivar Aksu (Varol, Reference Varol2020); 15.6–21.2% (desi) and 16.2–22.4% (kabuli) in Indian chickpea accessions (Upadhyaya et al., Reference Upadhyaya, Bajaj, Narnoliya, Das, Kumar, Gowda, Sharma, Tyagi and Parida2016). In the present study 37 genotypes that included both desi and kabuli types contained seed protein content of >27.0% at the two locations. Jadhav et al. (Reference Jadhav, Rayate, Mhase, Thudi, Chitikineni, Harer, Jadhav, Varshney and Kulwal2015) observed that protein content in 187 desi and kabuli chickpea genotypes ranged from 13.25–26.77% with mean value of 20.16% and the maximum number of genotypes contained protein content of about 18%. They also observed that the popular chickpea cultivars grown by farmers had relatively lower protein content and those with smaller seeds contained higher protein.
The Box plot analysis led to the identification of contrasting genotypes with the highest and the lowest protein content at the two locations (online Supplementary Fig. S1) and these included both desi and kabuli chickpea. The genotypes identified for higher seed protein in New Delhi involved ICC 13523 (33.56%, kabuli), ICC 13461 (31.72%, kabuli), ICC 9402 (31.06%, kabuli), ICC 8114 (30.22%, kabuli) and ICC 8397 (30.22%, desi). Similarly, the genotypes found for higher seed protein content at Ludhiana are GNG 2144 (30.44%, desi), ICC 13461 (28.79%, kabuli), ICC 13523 (28.31%, kabuli), H 82-2 (28.28%, desi) and ICC 9402 (28.10%, kabuli). Jadhav et al. (Reference Jadhav, Rayate, Mhase, Thudi, Chitikineni, Harer, Jadhav, Varshney and Kulwal2015) and Jukanti et al. (Reference Jukanti, Gaur, Gowda and Chibbar2012) have also reported that there was no correlation between seed type, testa colour and protein content in chickpea. The characteristics of genotypes selected for relatively high seed protein content (>27/100 g) at New Delhi and Ludhiana are given in Table 2. Twelve of the high protein genotypes selected was kabuli chickpea that included both small and large seeded types. However, all the high protein desi genotypes were small seeded. The two of the five traits specific accessions of global chickpea composite collection such as ICC 8397 and ICC 5912 included in this study contained an average protein content of 30.22 & 27.74% and 27.34 & 26.06%, respectively at New Delhi and Ludhiana. It was observed that both accessions produced blue flowers and pea shaped seeds with undesirable characteristics. The three kabuli chickpea accessions (ICC 13441, ICC 13461 and ICC 13523) are some of the new sources of variation (genes) for consistently high seed protein with desirable seed traits (shape, size and colour) identified from the present investigation at two locations which can be effectively utilized in chickpea breeding to further improve its nutritional value. Kabuli and desi are the two genetically diverse varietal groups of the same cultivated species, C. arietinum L. and thus there are no barriers in transferring genes for greater seed protein contents from the kabuli chickpea accessions to desi types through simple hybridization. Such genotypes that contain consistently high seed protein content over environments are also of immense use in understanding the genetics, identifying the genomic regions involved in its inheritance and further increase through molecular breeding. The study also resulted in the selection of chickpea released varieties with relatively high seed protein content (average 24–25/100 g and more) and is given in Table 3. The selected varieties included almost equal number of desi and kabuli seed types. The seed size of high protein containing kabuli varieties (22.3 to 38.2 g/100 seeds) was found to be greater than those of desi types (13.0 to 23.1 g/100 seeds). An important breeding objective in kabuli chickpea improvement is for their increased seed size along with the seed yield. The findings of the study suggests that past breeding efforts for increased seed size in kabuli chickpea had no adverse effect on their protein content and therefore it may be possible to breeding of higher yielding kabuli cultivars with greater seed size and protein contents that enhances their nutritional as well as export value.
FTIR spectrum and amino acid analysis
Amide is the protein group marker having distinctive infrared spectrum of proteins giving nine amide bands with vibrational contributions from both protein backbone and amino acids side chains. The absorptions associated with vibrational stretching of C = O are designated as Amide I, while absorption associated with bending vibrations of N-H bond are represented as Amide II (Barth, Reference Barth2007). Specific marker groups of amides A, B I, II, III and IV have been reported at 3300, 3100, 1600–1690, 1480–1575, 1229–1301 and 325–767 cm−1 wavelength range, respectively (Kong and Yu Reference Kong and Yu2007). Broad and prominent peaks for Amide I, II and III representing vibrations with amide specific marker group of proteins C = O, C-O and N-H in ICC13523 (high protein genotype), whereas minor splitting peaks observed in FG 212 (least protein genotype) indicate that C = O and N-H that are involved in hydrogen bonding of secondary structure of proteins are more prominent in high protein genotype.
Humans consume 60% of plant-based protein and 40% of animal protein on a global scale (FAOSTAT 2018). Plant proteins are in high demand due to concerns about health, the environment and animal welfare. Therefore, additional plant-based protein sources with high protein quality and a well-balanced amino acid composition will be required in the future (Ritchie et al., Reference Ritchie, Reay and Higgins2018). Dietary protein is hydrolysed into amino acids, dipeptides and tripeptides by proteases and peptidase into lumen of small intestine and so the relative proportions of amino acids are the determinants of protein quality. Amino acids cannot be replaced with other nutrients such as carbohydrates and lipids as they are precursors to hydrocarbon skeleton, nitrogen, sulphur, proteins, peptides, glutathione, nitric oxide, dopamine, serotonin, RNA and DNA (Wu Reference Wu2016). The content of particular amino acid in pulse proteins depends upon relative proportion of individual seed storage protein fractions. Seed storage proteins had specific amino acid composition. Chickpea seed protein has globulins as the major seed protein fraction (41.79%), followed by albumin (16.18%), glutelin (9.99%) and prolamin (0.48%) (Boye et al., Reference Boye, Aksay, Roufik, Ribéreau, Mondor, Farnworth and Rajamohamed2010). Globulins in comparison to albumins has higher content of glutamine, aspartic acid and arginine (Singh et al., Reference Singh, Wani, Kaur and Sogi2008) but had relatively lower content of sulphur containing amino acids and tryptophan (Duranti, Reference Duranti2006; Agarwal, Reference Agarwal2017). Albumin fraction is rich in sulphur containing amino acids (Semba et al., Reference Semba, Shardell, Sakr Ashour, Moaddel, Trehan, Maleta, Ordiz, Kraemer, Khadeer, Ferrucci and Manary2016).
Plants are capable of synthesizing all 20 amino acids contained in proteins. However humans can synthesize only 11 of them referred to as non-essential amino acids. Therefore, dietary sources are needed to fulfil the requirements of the remaining nine amino acids known as the essential amino acids (histidine, isoleucine, leucine, lysine, methionine, phenylalanine, threonine, tryptophan and valine). The sum of essential amino acids in the present study varied from 30.58/100 g protein in FG 212 (a small seeded desi cultivar) to 40.00/100 g protein in Pusa 5023 (a large seeded kabuli cultivar). The sum of essential amino acids in chickpea cultivars has been reported to be 40.81/100 g (Shaha et al., Reference Shaha, Ullahb and Munir2021). The quantity (g/100 g) of all the essential amino acids was found to be comparable with chickpea grown in Poland (Grela et al., Reference Grela, Samolińska, Kiczorowska, Klebaniuk and Kiczorowski2017); Brazil (Tavano et al., Reference Tavano, da Silva, Demonte and Neves2008); Egypt (Arab et al., Reference Arab, Helmy and Bareh2010); Pakistan (Iqbal et al., Reference Iqbal, Khalil, Ateeq and Khan2006; Zia-Ul-Haq et al., Reference Zia-Ul-Haq, Iqbal, Ahmed, Imran, Niaz and Bhanger2007) and Spain. Soybean seed is high in protein and is regarded as a ‘complete protein’ source since it has a good balance of all the essential amino acids required by humans and animals (Zarkadas et al., Reference Zarkadas, Gagnon, Gleddie, Khanizadeh, Cober and Guillemette2007). In this study, the amino acid profile of chickpea seeds was found to be comparable to that of soybean seeds indicating that chickpea also deserves to be considered as a source of complete protein.
The nutritional quality of a protein is determined by its essential amino acids content that must be comparable to the established reference standards as determined by WHO/FAO/UNU expert consultation. The quantity (g/100 g) of essential amino acids in chickpea seeds (Table 4) was found to be also comparable with the reference protein as recommended by WHO (1985) and FAO (2013). Moreover, chickpea seed has the potential to satisfy the dietary requirements of protein for all age groups as compared to the reference pattern of FAO (2013). Each amino acid has a different and essential function to carry out metabolic needs of the body. The identification of chickpea genotypes with higher amino acid contents will be useful in future research programmes to improve nutritional quality. The average values of Histidine and Lysine in chickpea seed protein are comparable to those in human milk and egg protein (Table 4). Leucine content of high protein chickpea genotype (T39-1) is found to be higher than egg protein and comparable to milk protein. Pusa112, Pusa5023 and ICC13287 have leucine content comparable to that of egg protein. Leucine stimulates muscle protein synthesis and under conditions of increased protein breakdown help to minimize muscle wasting (Davoodi et al., Reference Davoodi, Shahbazi, Esmaeili, Sohrabvandi, Mortazavian, Jazayeri and Taslimi2016). BGD132 and T39-1 are the chickpea genotypes found with higher lysine content (>8.0/100 g) that can serve as useful sources to complement lysine deficient cereal grain based diets as cereals are deficient in lysine. IG 72933 has a higher threonine content than the egg and milk protein; while L555 and T39-1 are higher in Valine content than the milk protein. In the present study, seed protein showed negative correlation with S-amino acids. For example, KGD1168, a low protein genotype had higher content of Sulphur containing amino acids than the milk protein and comparable with that of the egg protein. Sulphur containing amino acids are precursors of glutathione, which has antioxidant, anticarcinogenic and immunostimulatory properties (Miller et al., Reference Miller, Jarvis and McBean2007).
The sum of BCAA in chickpea were higher than the reference protein pattern recommended by WHO (1985) and FAO (2013) (Table 4) showing that chickpea grain protein can meet the dietary requirements of BCAA. Low protein chickpea genotypes had lower levels of BCAA (13.17/100 g) than high protein genotypes (15.66/100 g). Pusa112, Pusa5023, BGD132 and T39-1 are nutritionally important genotypes as they contained 17.46 to 18.83/100 g protein of BCAA which is comparable to milk and egg proteins (20.66/100 g). BCAA are essential for carrying out several metabolic and regulatory processes in humans and animals. These amino acids are the building blocks for muscle tissues, enhance protein synthesis, intestinal development, amino acids transportation in intestine and production of mucin; metabolic regulator in lipid and glucose metabolism, regulate signalling pathways; important for embryo growth and mammary health; upregulate innate and adaptive immune response (Zhang et al., Reference Zhang, Xiangfang Zeng, Man Ren, Mao and Qiao2017). The present study showed that glutamate was the most abundant amino acid among non-essential amino acids present in chickpea seeds followed by aspartic acid and arginine. This might be due to the higher globulin content in high protein genotypes. The proportion of globulin protein fraction was reported to be higher in high protein genotypes of pigeonpea as compared to genotypes with normal protein levels (Singh et al., Reference Singh, Jambunatha, Saxena and Subrahmanyam1990). The globulin proteins contained greater amounts of glutamine, aspartic acid, arginine and lysine (Hall et al., Reference Hall, Cassandra Hillen and Julie Robinson2017). The higher contents of aspartic and glutamic acid followed by arginine found in high protein chickpea genotypes is important for nutritional point of view as dietary glutamate, glutamine and aspartate serve as a major metabolic fuel for human intestine. Gupta et al. (Reference Gupta, Tripathi, Rai, Dwivedi, Mishra, Srivastava and Inouhe2006) also reported that proline, glutamic acid, lysine and aspartic acid were found in greater amounts in chickpea seeds. Moreover, glutamine is an exclusive source of energy to the arterial blood and also provides ATP for maintenance of immune system in macrophages and lymphocytes (Wu Reference Wu2016). In the present study, Pusa112, Pusa5023, IG72933, ICC8114, ICC9402, ICC13441 and BGD132 had higher contents of glutamic acid, aspartic acid and arginine (online Supplementary Table S1). The higher content of aspartic acid could be due to the fact that they are the nitrogen storage forms and the starting material for the formation of amino acid backbone (Onwuliri and Obu, Reference Onwuliri and Obu2002). Glutamic acid and aspartic acid played important regulatory roles in nutrition, energy metabolism and oxidative stress (Chen et al., Reference Chen, Wu, Duan, Huang, Li, Yin and Yin2021). Arginine is a semi-essential or conditionally essential amino acid as animals can synthesize it, but exclusion of arginine in the diet leads to sub-optimal weight gain (Tapiero et al., Reference Tapiero, Mathe, Couvreur and Tew2002). It is an intermediate of urea cycle, substrate for synthesizing ornithine, polyamines and an important signalling molecule involved in various physiological and pathological processes and also directly involved with synthesis of ATP, calcium release, neuro transmission, cell proliferation and immunity (Cortes-Giraldo et al., Reference Cortés-Giraldo, Megías, Alaiz, Girón-Calle and Vioque2016). The present study on characterization of chickpea germplasm identifying high protein genotypes rich in essential amino acids suggests that chickpea deserves to be considered as an important source of complete protein.
Conclusion
The characterization of 402 chickpea genotypes at two locations revealed wide range of genotypic variability for grain protein. Trait specific germplasm containing up to 33.56% grain protein identified for utilization in chickpea breeding. ICC13523, ICC13461, ICC9402, ICC8114, ICC8397, GNG2144 and H82-2 are some of the genotypes and cultivars containing higher protein content. High protein genotypes had greater average amino acid score for all the essential amino acids. The content of glutamic acid, aspartic acid and arginine was higher in high protein genotypes. The observed genotypic variation for protein content and amino acids could be used to improve nutritional quality of chickpea through transgressive breeding for germplasm enhancement and backcross breeding or marker assisted backcross breeding to introgress genomic regions controlling higher seed protein content into high yielding varieties. Further research on the transcriptomics and mapping is underway for the identification of genomic regions involved in the inheritance of grain protein content with the goal of using this information in genetic improvement of chickpea protein and its quality.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S147926212300028X
Acknowledgement
Authors are thankful for the financial support by ICAR-National Agricultural Science Fund (NASF) F. No NASF/ABP-6007/2016-17 to carry out this work. We are also grateful to Dr HD Upadhyaya, ICRISAT, Patancheru, for providing some of the trait specific chickpea accessions included in this study that are part of the Global Composite Collection. We thank the kind help of Dr KK Vinod, Genetics, for improving the quality of figures that are part of this manuscript.
Author contributions
Venkatraman Hegde and Satvir Kaur contributed to the study conception, design, material preparation and manuscript writing; Data collection and analysis were performed by Kanu Priya Sharma, Rachana D. Bharadwaj, Sukhpreet Kaur Sidhu, Sarvjeet Singh, Souliha Rasool and Dev Kumar Arya; Pradeep Kumar Jain, Pawan Kumar Agrawal and Biswajit Mondal contributed to reviewing and editing the manuscript. All authors commented on the original version of the manuscript as well as read and approved the final one.
Conflict of interest
The authors declare that they have no relevant financial or non-financial interests to disclose. Authors declare no conflict of interest.